


Epizootic Hemorrhagic Disease Virus: Current Knowledge and Emerging Perspectives

Abstract
:1. Introduction
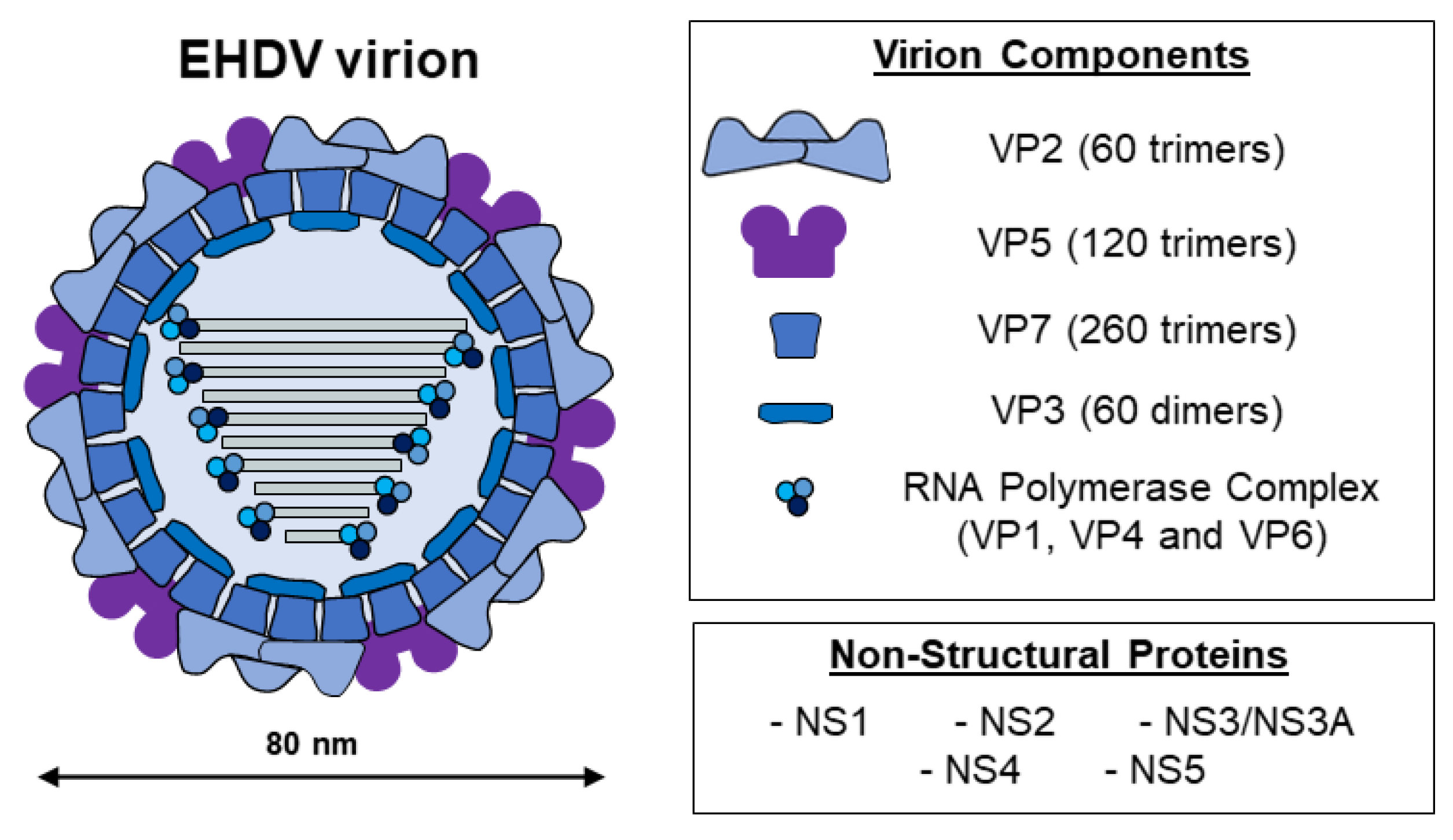
2. EHDV, the Etiological Agent of EHD
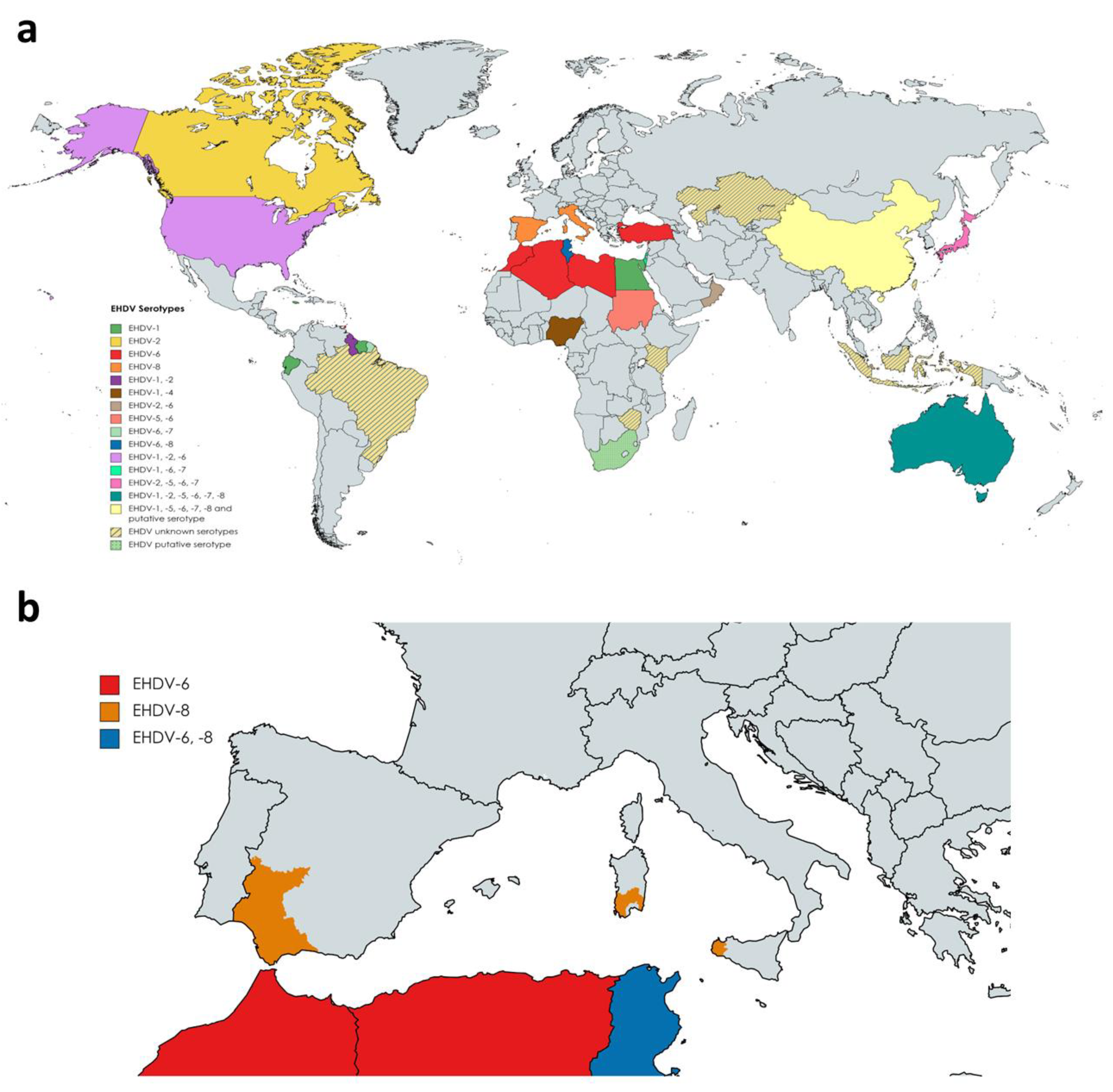
3. Changes in EHDV Epidemiology: Influence of Global Warming
Role of Culicoides Insect Vector in EHDV Spread
4. Disease and Pathology
5. Experimental Animal Models of EHDV
5.1. White-Tailed Deer (WTD) and Other Cervid Species

{kind=link}
{kind=link}
| Specie | Age | Challenge Virus | Dose | Inoculation Route | Mortality | Clinical Disease | Pathological Abnormalities | Viremia | Virus Isolation/RNA in Tissues | Gross and Histopathological Lessions | Immune Response | Ref |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Red deer (Cervus elaphus) | 1–4 years old | EHDV-1 USA1955/01 | 106 TCID50 | IV | No | No | No | Yes a | Not evaluated | Not evaluated | Homologous nAbs | [123] |
| Fallow deer (Dama dama) | 1–4 years old | EHDV-1 USA1955/01 | 106 TCID50 | IV | No | No | No | Yes a | Not evaluated | Not evaluated | Homologous nAbs | [123] |
| Roe deer (Capreolus capreolus) | 1–4 years old | EHDV-1 USA1955/01 | 106 TCID50 | IV | No | No | No | Yes a | Not evaluated | Not evaluated | Homologous nAbs | [123] |
| Muntjac deer (Muntiacus muntjac) | 1–4 years old | EHDV-1 USA1955/01 | 106 TCID50 | IV | No | No | No | Yes a | Not evaluated | Not evaluated | Homologous nAbs | [123] |
| White-Tailed deer (Odocoileus virginianus) | 2–24 months old | EHDV-1 USA1955/01 | Unknown (inocula from spleen, liver, lung, blood, and kidney of severely infected WTD) | SC or IM | Yes (>56% mortality rate) | Severe to fatal clinical disease | Not evaluated | Not evaluated | Not evaluated | Yes | Homologous nAbs | [125] |
| EHDV isolated in South Dakota (no data specified) | Yes (>33% mortality rate) | |||||||||||
| 9–36 months old | EHDV-1 USA1955/01 | Unknown (inoculum from severely infected WTD spleen) | IM | Yes (>50% mortality rate) | Yes | Thrombocytopenia | Not evaluated | Not evaluated | Yes | Not evaluated | [124] | |
| 5 months old | EHDV-6 (EHDV-6/EHDV-2 reassortant isolate CC-304-06 Indiana, US, 2006) | 106.4 TCID50 | SC and ID | Yes (60% mortality rate) | Yes | Leukopenia, lymphopenia, hypoproteinemia | Yes a | Spleen, lung, lymph node and skin | Yes | Homologous nAbs | [126] | |
| 8 months old | EHDV-7 ISR2006/04 | 105.27 TCID50 (inoculum from severely infected WTD blood) | SC and ID | Yes (66.7% mortality rate) | Yes | Leukopenia, lymphopenia, hypoproteinemia, thrombocytopenia | Yes a | Spleen, lung, lymph node, heart, cerebellum, cerebrum and skin | Yes | Homologous nAbs | [127] | |
| 2-months-old Odocoileus virginianus texanus | EHDV-1 (from field isolate from Walton County, Georgia in 1999) | 107 TCID50 (inoculum from severely infected WTD blood) | SC and ID | No | Mild clinical disease | No | Yes a | Not evaluated | Not evaluated | Homologous nAbs | [128] | |
| EHDV-2 (field isolate from Clarke County, Georgia) | 107.1 TCID50 (inoculum from severely infected WTD blood) | No | ||||||||||
| 2-months-old Odocoileus virginianus borealis | EHDV-1 (from field isolate from Walton County, Georgia in 1999) | 107.1 TCID50 (inoculum from severely infected WTD blood) | Yes (100% mortality rate) | Severe clinical disease | Lymphopenia, hypoproteinemia | Gross lesions (not specified) | ||||||
| EHDV-2 (field isolate from Clarke County, Georgia) | 107 TCID50 (inoculum from severely infected WTD blood) | Yes (20% mortality rate) | ||||||||||
| 3–4 months old | EHDV-2 (field isolate from Clarke County, Georgia in 1990) | 105.5 TCID50 (inoculum from severely infected WTD blood) | SC and ID | Yes (31% mortality rate) | Yes | Lymphopenia | Yes a | Not evaluated | Not evaluated | Homologous nAbs | [118] | |
| 3–4-months-old EHDV-2 convalescent animals | EHDV-2 (field isolate from Clarke County, Georgia in 1990) | 103 TCID50 (inoculum from severely infected WTD blood) | No | No | No | No a | Not evaluated | No | ||||
| 27–47-days-old fawns (feed with colostrum with maternal nAbs) | EHDV-2 (field isolate from Clarke County, Georgia in 2016) | 105.6 TCID50 (inoculum from virus isolated in severely infected WTD spleen and propagated in BHK21 cells) | SC and ID | No | Mild or absent clinical disease | Not evaluated | No (transient viraemia in two fawns a) | Spleen | No | Maternal homologous nAbs | [131] | |
| 27–47-days-old fawns | No | Moderate clinical disease | Yes a | |||||||||
| 3–4 months old | EHDV-2 (strain not specified) | 105.5 TCID50 (inoculum from infected WTD blood) | SC and ID | Yes (25% mortality rate) | Yes | Lymphopenia | Yes a | Not evaluated | Not evaluated | Homologous nAbs | [132] | |
| 4–5 months old | EHDV-2 (from field isolate from Clarke County, Georgia in 2016) | 106.6 TCID50 (inoculum from infected WTD blood) | SC and ID | No | Moderate clinical disease | Not evaluated | Yes a | Not evaluated | Not evaluated | Homologous nAbs and Cell-mediated response | [103] | |
| EHDV-1 (from field isolate from Walton County, Georgia in 1999) | 107.6 TCID50 (inoculum from virus isolated in severely infected WTD spleen and propagated in BHK21 cells) | Yes (100% mortality rate) | Severe clinical disease | Lymphopenia, thrombocytopenia | ||||||||
| 4–5-months-old EHDV-2 convalescent animals | EHDV-1 (from field isolate from Walton County, Georgia in 1999) | 107.6 TCID50 (inoculum from virus isolated in severely infected WTD spleen and propagated in BHK21 cells) | No | Mild or absent clinical disease | No | |||||||
| 4 months old | EHDV-2 (from field isolate from Clarke County, Georgia in 1990) | 105 TCID50 (inoculum from infected WTD blood) | SC and ID | Not specified | Not evaluated | Not evaluated | Yes a | Not evaluated | Not evaluated | Not evaluated | [134] | |
| 6-months-old Odocoileus virginianus borealis | EHDV-2 (not specified) | 107.03 TCID50 | Not specified | Yes (25% mortality rate) | Moderate or severe clinical disease | Not specified | Yes a | Spleen, lung, buccal mucosa and skin | Yes | Induction of pro-inflammatory cytokines | [115] | |
| 6-months-old Odocoileus virginianus texanus | Mild or absent clinical disease | No | ||||||||||
| 8 months old | EHDV-7 ISR2006/04 | Unknown | Infected C. sonorensis | Yes * | Severe clinical disease | Leukopenia, lymphopenia, hypoproteinemia, thrombocytopenia | Yes a | Cerebrum, cerebellum, heart, lung, spleen, lymph node, skin, epididymis | Yes | Homologous nAbs | [133] | |
| 4–7 months old | EHDV-7 ISR2006/04 | 105.1 TCID50 (inoculum from virus isolated in severely infected WTD spleen and propagated in BHK21 cells) | SC and ID | Yes * | Not evaluated | Not evaluated | Yes a | Not evaluated | Not evaluated | Not evaluated | [135] | |
| 7 months old | EHDV-2 (from field isolate from Coffey County, Kansas in 2012) | 106.5 TCID50 (inoculum from virus isolated in infected WTD spleen and propagated in CPAE, BHK21, and CuVaW8A cells) | SC and ID | Not specified * | Mild to moderate clinical disease | Not evaluated | Yes a | Not evaluated | Not evaluated | Not evaluated | [136] | |
| 4–6 months old | EHDV-1 (from field isolate from Walton County, Georgia in 1999) | 107.1 TCID50 (inoculum from virus isolated in severely infected WTD spleen and propagated in BHK21 cells) | SC and ID | No | Mild to severe clinical disease | Not evaluated | Yes a | Virus isolated from rectal and oral cavities | Not evaluated | Not evaluated | [137] |
5.2. Cattle and Other Farm Animals
5.3. Mouse Models
| Species | Breed | Challenge Virus | Dose | Inoculation Route | Mortality | Clinical Disease | Viremia | RNA in Tissues | Tissue Damage | Immune Response | Ref |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cattle | Japanese black | Kyushu-1 and Ibaraki-5 strains | 1-5 mL infected blood | IV | No | Yes (conjunctivae and oro-nasal inflammation, fever, leukopenia) | Not evaluated | Not evaluated | Necrotic changes of epithelial cells | Not evaluated | [67] |
| 6–18-months-old Jersey or Friesian cattle | EHDV-1 USA1955/01 | 106 TCID50 | IV | No | No | Yes a | Not evaluated | Not evaluated | Homologous nAbs | [123] | |
| 4–6-months-old calves | EHDV-1 USA1955/01 | 106 TCID50 | ID and SC | No | No | Yes a,b | Not evaluated | Not evaluated | Humoral response to EHDV-1 or EHDV-2 (neutralization not tested) | [139] | |
| EHDV-2 CAN1962/01 | |||||||||||
| 7–9-months-old Holstein-Friesian cattle | EHDV-6 TUR2007/01 | 107.5 TCID50 | SC | No | No | Yes a,c | Spleen and lymph nodes | No | Homologous nAbs | ||
| EHDV-6 MOR2006/17 | 107.5 TCID50 | ||||||||||
| 12-months-old Holstein cattle | EHDV-7/ISR2006/13 | 5 × 105 TCID50 | SC | No | No | Yes a,c | Not evaluated | Not evaluated | Homologous nAbs | [144] | |
| 18-months-old Holstein cattle | EHDV-6 (Reunión Island, 2008) | Two doses of 6 × 106 TCID50 | SC | No | Yes (conjunctivitis, epiphora. One animal displayed moderate oedema, gingival ulcer) | Yes a,c | Spleen, liver, skin, kidney, lymph nodes and heart | Not evaluated | Homologous nAbs | [143] | |
| 1-year-old Holstein steer and adult Holstein cattle | EHDV-6 (EHDV-6/EHDV-2 reassortant isolate CC-304-06 Indiana, US, 2006) | 107.27 TCID50 | SC and ID | No | No | Yes a | No | No | Homologous nAbs | [126] | |
| >4-years-old Holstein cattle | EHDV-7 ISR2006/04 (WTD blood inoculum) | 106.1 TCID50 | ID and SC | No | No | Yes c | Spleen and lymph node | No | Homologous nAbs | [147] | |
| 2-month-old Holstein calves | EHDV-7 ISR2006/04 (BHK cell culture supernatant) | 107.12 TCID50 | ID and SC | No | No | Yes c | Spleen, lung and lymph node | No | Homologous nAbs | ||
| ID, IV and SC | No | No (elevated rectal temperature at two timepoints) | Yes c | Spleen, lung and lymph node | No | Homologous nAbs | |||||
| EHDV-7 ISR2006/04 | Undetermined | Infected C. sonorensis | No | No | Yes c | Spleen, lung and lymph node | No | Homologous nAbs | |||
| 2-month-old Prim’Holstein calves | EHDV-1 USA1955/01 | 107 TCID50 * | I.M. | No | No | Yes c | Not evaluated | Not evaluated | Homologous nAbs. Low nAb titers against EHDV-7 ISR2006/01 | [145] | |
| EHDV-2 CAN1962/01 | 5 × 107 TCID50 * | No | No | Homologous nAbs. Low nAb titers against EHDV-1 USA1955/01 and EHDV-7 ISR2006/01 | |||||||
| EHDV-4 NIG1968/01 | 3 × 106 TCID50 * | No | No | Homologous nAbs. Low nAb titers against EHDV-5 AUS1979/06 | |||||||
| EHDV-5 AUS1979/06 | 1.6 × 106 TCID50 * | No | No | Homologous nAbs. Low nAb titers against EHDV-4 NIG1968/01 | |||||||
| EHDV-6 AUS1981/07 | 107 TCID50 * | No | No | Homologous nAbs. | |||||||
| EHDV-7 ISR2006/01 | 107 TCID50 * | Yes (euthanasia) | Yes (apathy, diarrhea, prostration, unable to stand, anorexia) | Homologous nAbs. Low nAb titers against EHDV-2 CAN1962/01 | |||||||
| EHDV-8 AUS1982/06 | 5 × 107 TCID50 * | No | No | Homologous nAbs. Low nAb titers against EHDV-6 AUS1981/07 | |||||||
| Sheep | 12–24-months-old Suffolk cross or Dorset Horn sheep | EHDV-1 USA1955/01 | 106 TCID50 | IV | No | No | Yes a | Not evaluated (Presence of virus in vulvae of a viraemic sheep | Not evaluated | Homologous nAbs | [123] |
| Cheviot-SouthDown sheep (unknow age) | EHDV-1 USA1955/01 | Unknown (inoculum from severely infected WTD spleen) | IM | No | No | Not evaluated | Not evaluated | No | Not evaluated | [124] | |
| 10-months-old East Frisian sheep | EHDV-7/ISR2006/13 | 5 × 105 TCID50 | SC | No | No | No | Not evaluated | Not evaluated | Not evaluated | [144] | |
| Goat | 6–24-months-old British Alpine goat | EHDV-1 USA1955/01 | 106 TCID50 | IV | No | No | No | Not evaluated | Not evaluated | Homologous nAbs | [123] |
| Pig | 3-months-old Large Whites | EHDV-1 USA1955/01 | 106 TCID50 | IV | No | No | No | Not evaluated | Not evaluated | Homologous nAbs | [123] |
| Mouse | Swiss newborn mice | EHDV-1 USA1955/01 | Not specified | IC | Yes (98.6% mortality rate, 436 out of 442 mice) | Yes (loss of postural reflexes, irregular respiration, cyanosis, and tonic and clonic convulsions) | Not evaluated | Not evaluated | Not evaluated | Not evaluated | [156,157] |
| Adult IFNAR(−/−) mice | EHDV-7/ISR2006/13 | Unknown (blood inoculum from infected cattle) | IP | Yes (8.33% mortality rate, 1 out of 12 mice) | Yes (ruffled fur, apathy (no conjunctivitis)) | Yes c | Spleen (in 10 out of 12 mice) | Enlarged spleens | Not evaluated | [144] | |
| Adult IFNAR(−/−) mice | EHDV-7/ISR2006/13 | 5 × 102 TCID50 | IP | Yes (30% mortality rate, 1 out of 3 mice) | Yes (ruffled fur, apathy (no conjunctivitis)) | Not evaluated | Spleen (in both dead and surviving mice) | Enlarged spleens and necrotic foci in liver in dead animals) | Not evaluated | [159] | |
| 5 × 105 TCID50 | Yes (100% mortality rate, 2 out of 2 mice) |
6. Classic and Novel Vaccine Approaches against EHDV
| Vaccine Type | Antigen Included | Serotype of Antigen | Animal Model | Dose | Adjuvant | Immunogenicity | Challenge | Protection | Ref |
|---|---|---|---|---|---|---|---|---|---|
| Recombinant VP2 protein 1 | VP2 | EHDV-6 (Indiana 2012, 12-38993) | 6-weeks-old female CD1-ISG | Two doses of 20 μg | Montanide ISA25 adjuvant | Neutralization against EHDV-6 | Not challenged | - | [164] |
| 3-4-months-old male Holstein | Two doses of 150 μg | Neutralization against EHDV-6 | |||||||
| VP2 | EHDV-2 (Alberta 1962, SV-124-Canada) | 6-weeks-old female CD1-ISG | Two doses of 20 μg | Montanide ISA25 adjuvant | Neutralization against EHDV-2 | ||||
| 4-months-old male Holstein calves | Two doses of 150 μg | Neutralization against EHDV-2 | |||||||
| 5-month-old male WTD | Two doses of 150 μg | Neutralization against EHDV-2 | 106.74 PFU EHDV-2 (Kansas 2012, strain cc12-304) | Yes a,b,c,d,e | |||||
| CLP 1 | VP7, VP3 | EHDV-1 USA1955/01 | Rabbit | Three doses (first dose: 500 µg; second and third doses: 250 µg) | Incomplete Fruend’s adjuvant | Induction of VP3- and VP7-specific antibodies. | Not challenged | - | [166] |
| VLP 1 | VP2, VP5, VP7, VP3 | Neutralization against EHDV-1/Low neutralization against EHDV-2 and EHDV-6 | |||||||
| VLP 1 | VP2, VP5, VP7, VP3 | VP3 and VP7 from EHDV-1 USA1955/01. VP2 and VP5 from EHDV-2 CAN1962/01 | Not evaluated in animal model | - | - | - | - | - | |
| VLP 1 | VP2, VP5, VP7, VP3 | EHDV-6 (MOR2006/05) | Not evaluated in animal model | - | - | - | - | - | [168] |
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Huismans, H.; Bremer, C.W.; Barber, T.L. The Nucleic Acid and Proteins of Epizootic Haemorrhagic Disease Virus. Onderstepoort J. Vet. Res. 1979, 46, 95–104. [Google Scholar]
- Stewart, M.; Hardy, A.; Barry, G.; Pinto, R.M.; Caporale, M.; Melzi, E.; Hughes, J.; Taggart, A.; Janowicz, A.; Varela, M.; et al. Characterization of a Second Open Reading Frame in Genome Segment 10 of Bluetongue Virus. J. Gen. Virol. 2015, 96, 3280–3293. [Google Scholar] [CrossRef]
- Belhouchet, M.; Mohd Jaafar, F.; Firth, A.E.; Grimes, J.M.; Mertens, P.P.C.; Attoui, H. Detection of a Fourth Orbivirus Non-Structural Protein. PLoS ONE 2011, 6, e25697. [Google Scholar] [CrossRef]
- Mecham, J.O.; Dean, V.C. Protein Coding Assignment for the Genome of Epizootic Haemorrhagic Disease Virus. J. Gen. Virol. 1988, 69 Pt 6, 1255–1262. [Google Scholar] [CrossRef]
- Zhang, X.; Boyce, M.; Bhattacharya, B.; Zhang, X.; Schein, S.; Roy, P.; Zhou, Z.H. Bluetongue Virus Coat Protein VP2 Contains Sialic Acid-Binding Domains, and VP5 Resembles Enveloped Virus Fusion Proteins. Proc. Natl. Acad. Sci. USA 2010, 107, 6292–6297. [Google Scholar] [CrossRef]
- Urakawa, T.; Ritter, D.G.; Roy, P. Expression of Largest RNA Segment and Synthesis of VP1 Protein of Bluetongue Virus in Insect Cells by Recombinant Baculovirus: Association of VP1 Protein with RNA Polymerase Activity. Nucleic Acids Res. 1989, 17, 7395–7401. [Google Scholar] [CrossRef]
- Stäuber, N.; Martinez-Costas, J.; Sutton, G.; Monastyrskaya, K.; Roy, P. Bluetongue Virus VP6 Protein Binds ATP and Exhibits an RNA-Dependent ATPase Function and a Helicase Activity That Catalyze the Unwinding of Double-Stranded RNA Substrates. J. Virol. 1997, 71, 7220–7226. [Google Scholar] [CrossRef]
- Sutton, G.; Grimes, J.M.; Stuart, D.I.; Roy, P. Bluetongue Virus VP4 Is an RNA-Capping Assembly Line. Nat. Struct. Mol. Biol. 2007, 14, 449–451. [Google Scholar] [CrossRef]
- Wu, W.; Roy, P. Sialic Acid Binding Sites in VP2 of Bluetongue Virus and Their Use during Virus Entry. J. Virol. 2022, 96, e0167721. [Google Scholar] [CrossRef]
- Anthony, S.J.; Maan, S.; Maan, N.; Kgosana, L.; Bachanek-Bankowska, K.; Batten, C.; Darpel, K.E.; Sutton, G.; Attoui, H.; Mertens, P.P.C. Genetic and Phylogenetic Analysis of the Outer-Coat Proteins VP2 and VP5 of Epizootic Haemorrhagic Disease Virus (EHDV): Comparison of Genetic and Serological Data to Characterise the EHDV Serogroup. Virus Res. 2009, 145, 200–210. [Google Scholar] [CrossRef]
- Forzan, M.; Marsh, M.; Roy, P. Bluetongue Virus Entry into Cells. J. Virol. 2007, 81, 4819–4827. [Google Scholar] [CrossRef]
- Maan, N.S.; Maan, S.; Potgieter, A.C.; Wright, I.M.; Belaganahalli, M.; Mertens, P.P.C. Development of Real-Time RT-PCR Assays for Detection and Typing of Epizootic Haemorrhagic Disease Virus. Transbound. Emerg. Dis. 2017, 64, 1120–1132. [Google Scholar] [CrossRef]
- Sun, F.; Cochran, M.; Beckham, T.; Clavijo, A. Molecular Typing of Epizootic Hemorrhagic Disease Virus Serotypes by One-Step Multiplex RT-PCR. J. Wildl. Dis. 2014, 50, 639–644. [Google Scholar] [CrossRef]
- Anthony, S.J.; Maan, N.; Maan, S.; Sutton, G.; Attoui, H.; Mertens, P.P.C. Genetic and Phylogenetic Analysis of the Core Proteins VP1, VP3, VP4, VP6 and VP7 of Epizootic Haemorrhagic Disease Virus (EHDV). Virus Res. 2009, 145, 187–199. [Google Scholar] [CrossRef]
- Mecham, J.O.; Stallknecht, D.; Wilson, W.C. The S7 Gene and VP7 Protein Are Highly Conserved among Temporally and Geographically Distinct American Isolates of Epizootic Hemorrhagic Disease Virus. Virus Res. 2003, 94, 129–133. [Google Scholar] [CrossRef]
- Bréard, E.; Viarouge, C.; Donnet, F.; Sailleau, C.; Rossi, S.; Pourquier, P.; Vitour, D.; Comtet, L.; Zientara, S. Evaluation of a Commercial ELISA for Detection of Epizootic Haemorrhagic Disease Antibodies in Domestic and Wild Ruminant Sera. Transbound. Emerg. Dis. 2020, 67, 2475–2481. [Google Scholar] [CrossRef]
- Forzan, M.; Pizzurro, F.; Zaccaria, G.; Mazzei, M.; Spedicato, M.; Carmine, I.; Salini, R.; Tolari, F.; Cerri, D.; Savini, G.; et al. Competitive Enzyme-Linked Immunosorbent Assay Using Baculovirus-Expressed VP7 for Detection of Epizootic Haemorrhagic Disease Virus (EHDV) Antibodies. J. Virol. Methods 2017, 248, 212–216. [Google Scholar] [CrossRef]
- Serroni, A.; Ulisse, S.; Iorio, M.; Laguardia, C.; Testa, L.; Armillotta, G.; Caporale, M.; Salini, R.; Lelli, D.; Wernery, U.; et al. Development of a Competitive Enzyme-Linked Immunosorbent Assay Based on Purified Recombinant Viral Protein 7 for Serological Diagnosis of Epizootic Haemorrhagic Disease in Camels. J. Trop. Med. 2022, 2022, 5210771. [Google Scholar] [CrossRef]
- Kusari, J.; Roy, P. Molecular and Genetic Comparisons of Two Serotypes of Epizootic Hemorrhagic Disease of Deer Virus. Am. J. Vet. Res. 1986, 47, 1713–1717. [Google Scholar]
- Nel, L.H.; Picard, L.A.; Huismans, H. A Characterization of the Nonstructural Protein from Which the Virus-Specified Tubules in Epizootic Haemorrhagic Disease Virus-Infected Cells Are Composed. Virus Res. 1991, 18, 219–230. [Google Scholar] [CrossRef]
- Owens, R.J.; Limn, C.; Roy, P. Role of an Arbovirus Nonstructural Protein in Cellular Pathogenesis and Virus Release. J. Virol. 2004, 78, 6649–6656. [Google Scholar] [CrossRef] [PubMed]
- Boyce, M.; Celma, C.C.P.; Roy, P. Bluetongue Virus Non-Structural Protein 1 Is a Positive Regulator of Viral Protein Synthesis. Virol. J. 2012, 9, 178. [Google Scholar] [CrossRef]
- Anthony, S.J.; Maan, N.; Maan, S.; Sutton, G.; Attoui, H.; Mertens, P.P.C. Genetic and Phylogenetic Analysis of the Non-Structural Proteins NS1, NS2 and NS3 of Epizootic Haemorrhagic Disease Virus (EHDV). Virus Res. 2009, 145, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Clavijo, A.; Sun, F.; Lester, T.; Jasperson, D.C.; Wilson, W.C. An Improved Real-Time Polymerase Chain Reaction for the Simultaneous Detection of All Serotypes of Epizootic Hemorrhagic Disease Virus. J. Vet. Diagn. Investig. 2010, 22, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Becker, M.E.; Healy, S.; Forbes, W.; Roberts, J.; LaCour, J.; Foil, L.D. Postmortem detection of bluetongue and epizootic hemorrhagic disease viruses in the bone marrow of white-tailed deer (Odocoileus virginianus). J. Wildl. Dis. 2020, 56, 58–65. [Google Scholar] [CrossRef]
- Theron, J.; Huismans, H.; Nel, L.H. Identification of a Short Domain within the Non-Structural Protein NS2 of Epizootic Haemorrhagic Disease Virus That Is Important for Single Strand RNA-Binding Activity. J. Gen. Virol. 1996, 77, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Uitenweerde, J.M.; Theron, J.; Stoltz, M.A.; Huismans, H. The Multimeric Nonstructural NS2 Proteins of Bluetongue Virus, African Horsesickness Virus, and Epizootic Hemorrhagic Disease Virus Differ in Their Single-Stranded RNA-Binding Ability. Virology 1995, 209, 624–632. [Google Scholar] [CrossRef]
- Theron, J.; Huismans, H.; Nel, L.H. Site-Specific Mutations in the NS2 Protein of Epizootic Haemorrhagic Disease Virus Markedly Affect the Formation of Cytoplasmic Inclusion Bodies. Arch. Virol. 1996, 141, 1143–1151. [Google Scholar] [CrossRef]
- Jensen, M.J.; Wilson, W.C. A Model for the Membrane Topology of the NS3 Protein as Predicted from the Sequence of Segment 10 of Epizootic Haemorrhagic Disease Virus Serotype 1. Arch. Virol. 1995, 140, 799–805. [Google Scholar] [CrossRef]
- Hyatt, A.D.; Zhao, Y.; Roy, P. Release of Bluetongue Virus-like Particles from Insect Cells Is Mediated by BTV Nonstructural Protein NS3/NS3A. Virology 1993, 193, 592–603. [Google Scholar] [CrossRef]
- Jensen, M.J.; Cheney, I.W.; Thompson, L.H.; Mecham, J.O.; Wilson, W.C.; Yamakawa, M.; Roy, P.; Gorman, B.M. The Smallest Gene of the Orbivirus, Epizootic Hemorrhagic Disease, Is Expressed in Virus-Infected Cells as Two Proteins and the Expression Differs from That of the Cognate Gene of Bluetongue Virus. Virus Res. 1994, 32, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Fablet, A.; Kundlacz, C.; Dupré, J.; Hirchaud, E.; Postic, L.; Sailleau, C.; Bréard, E.; Zientara, S.; Vitour, D.; Caignard, G. Comparative Virus-Host Protein Interactions of the Bluetongue Virus NS4 Virulence Factor. Viruses 2022, 14, 182. [Google Scholar] [CrossRef] [PubMed]
- Ratinier, M.; Shaw, A.E.; Barry, G.; Gu, Q.; Di Gialleonardo, L.; Janowicz, A.; Varela, M.; Randall, R.E.; Caporale, M.; Palmarini, M. Bluetongue Virus NS4 Protein Is an Interferon Antagonist and a Determinant of Virus Virulence. J. Virol. 2016, 90, 5427–5439. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.H.; George, T.D.S. A Preliminary Report of a Comparison of Epizootic Haemorrhagic Disease Viruses from Australia with Others from North America, Japan and Nigeria. Aust. Vet. J. 1986, 63, 233. [Google Scholar] [CrossRef] [PubMed]
- Noronha, L.E.; Cohnstaedt, L.W.; Richt, J.A.; Wilson, W.C. Perspectives on the Changing Landscape of Epizootic Hemorrhagic Disease Virus Control. Viruses 2021, 13, 2268. [Google Scholar] [CrossRef]
- Shope, R.E.; Lester, G.M.; Robert, M. Deer Mortality—Epizootic Hemorrhagic Disease of Deer. N. J. Outdoors 1955, 6, 17–21. [Google Scholar]
- Ruder, M.G.; Lysyk, T.J.; Stallknecht, D.E.; Foil, L.D.; Johnson, D.J.; Chase, C.C.; Dargatz, D.A.; Gibbs, E.P.J. Transmission and Epidemiology of Bluetongue and Epizootic Hemorrhagic Disease in North America: Current Perspectives, Research Gaps, and Future Directions. Vector Borne Zoonotic Dis. 2015, 15, 348–363. [Google Scholar] [CrossRef]
- Ruder, M.G.; Johnson, D.; Ostlund, E.; Allison, A.B.; Kienzle, C.; Phillips, J.E.; Poulson, R.L.; Stallknecht, D.E. The First 10 Years (2006-15) of Epizootic Hemorrhagic Disease Virus Serotype 6 in the USA. J. Wildl. Dis. 2017, 53, 901–905. [Google Scholar] [CrossRef]
- Schirtzinger, E.E.; Jasperson, D.C.; Ruder, M.G.; Stallknecht, D.E.; Chase, C.C.L.; Johnson, D.J.; Ostlund, E.N.; Wilson, W.C. Evaluation of 2012 US EHDV-2 Outbreak Isolates for Genetic Determinants of Cattle Infection. J. Gen. Virol. 2019, 100, 556–567. [Google Scholar] [CrossRef]
- Weir, R.P.; Harmsen, M.B.; Hunt, N.T.; Blacksell, S.D.; Lunt, R.A.; Pritchard, L.I.; Newberry, K.M.; Hyatt, A.D.; Gould, A.R.; Melville, L.F. EHDV-1, a New Australian Serotype of Epizootic Haemorrhagic Disease Virus Isolated from Sentinel Cattle in the Northern Territory. Vet. Microbiol. 1997, 58, 135–143. [Google Scholar] [CrossRef]
- Ahmed, S.; Mahmoud, M.A.E.-F.; Viarouge, C.; Sailleau, C.; Zientara, S.; Breard, E. Presence of Bluetongue and Epizootic Hemorrhagic Disease Viruses in Egypt in 2016 and 2017. Infect. Genet. Evol. 2019, 73, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Viarouge, C.; Lancelot, R.; Rives, G.; Bréard, E.; Miller, M.; Baudrimont, X.; Doceul, V.; Vitour, D.; Zientara, S.; Sailleau, C. Identification of Bluetongue Virus and Epizootic Hemorrhagic Disease Virus Serotypes in French Guiana in 2011 and 2012. Vet. Microbiol. 2014, 174, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Wang, F.; Chang, J.; Zhang, Y.; Zhu, J.; Li, H.; Yu, L. Identification and Complete-Genome Phylogenetic Analysis of an Epizootic Hemorrhagic Disease Virus Serotype 7 Strain Isolated in China. Arch. Virol. 2019, 164, 3121–3126. [Google Scholar] [CrossRef] [PubMed]
- Yadin, H.; Brenner, J.; Bumbrov, V.; Oved, Z.; Stram, Y.; Klement, E.; Perl, S.; Anthony, S.; Maan, S.; Batten, C.; et al. Epizootic Haemorrhagic Disease Virus Type 7 Infection in Cattle in Israel. Vet. Rec. 2008, 162, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Temizel, E.M.; Yesilbag, K.; Batten, C.; Senturk, S.; Maan, N.S.; Mertens, P.P.C.; Batmaz, H. Epizootic Hemorrhagic Disease in Cattle, Western Turkey. Emerg. Infect. Dis. 2009, 15, 317–319. [Google Scholar] [CrossRef]
- Verdezoto, J.; Breard, E.; Viarouge, C.; Quenault, H.; Lucas, P.; Sailleau, C.; Zientara, S.; Augot, D.; Zapata, S. Novel Serotype of Bluetongue Virus in South America and First Report of Epizootic Haemorrhagic Disease Virus in Ecuador. Transbound. Emerg. Dis. 2018, 65, 244–247. [Google Scholar] [CrossRef]
- Rajko-Nenow, P.; Brown-Joseph, T.; Tennakoon, C.; Flannery, J.; Oura, C.A.L.; Batten, C. Detection of a Novel Reassortant Epizootic Hemorrhagic Disease Virus Serotype 6 in Cattle in Trinidad, West Indies, Containing Nine RNA Segments Derived from Exotic EHDV Strains with an Australian Origin. Infect. Genet. Evol. 2019, 74, 103931. [Google Scholar] [CrossRef]
- Mohammed, M.E.; Aradaib, I.E.; Mukhtar, M.M.; Ghalib, H.W.; Riemann, H.P.; Oyejide, A.; Osburn, B.I. Application of Molecular Biological Techniques for Detection of Epizootic Hemorrhagic Disease Virus (EHDV-318) Recovered from a Sentinel Calf in Central Sudan. Vet. Microbiol. 1996, 52, 201–208. [Google Scholar] [CrossRef]
- Aradaib, I.E.; Mohammed, M.E.; Mukhtar, M.M.; Ghalib, H.W.; Osburn, B.I. Serogrouping and Topotyping of Sudanese and United States Strains of Epizootic Hemorrhagic Disease Virus Using PCR. Comp. Immunol. Microbiol. Infect. Dis. 1997, 20, 211–218. [Google Scholar] [CrossRef]
- Sghaier, S.; Sailleau, C.; Marcacci, M.; Thabet, S.; Curini, V.; Ben Hassine, T.; Teodori, L.; Portanti, O.; Hammami, S.; Jurisic, L.; et al. Epizootic Haemorrhagic Disease Virus Serotype 8 in Tunisia, 2021. Viruses 2022, 15, 16. [Google Scholar] [CrossRef]
- Duan, Y.; Yang, Z.; Zhu, P.; Xiao, L.; Li, Z.; Li, Z.; Li, L.; Zhu, J. A Serologic Investigation of Epizootic Hemorrhagic Disease Virus in China between 2014 and 2019. Virol. Sin. 2022, 37, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Mejri, S.; Dhaou, S.B.; Jemli, M.; Bréard, E.; Sailleau, C.; Sghaier, S.; Zouari, M.; Lorusso, A.; Savini, G.; Zientara, S.; et al. Epizootic Haemorrhagic Disease Virus Circulation in Tunisia. Vet. Ital. 2018, 54, 87–90. [Google Scholar] [CrossRef]
- Yamamoto, K.; Hiromatsu, R.; Kaida, M.; Kato, T.; Yanase, T.; Shirafuji, H. Isolation of Epizootic Hemorrhagic Disease Virus Serotype 7 from Cattle Showing Fever in Japan in 2016 and Improvement of a Reverse Transcription-Polymerase Chain Reaction Assay to Detect Epizootic Hemorrhagic Disease Virus. J. Vet. Med. Sci. 2021, 83, 1378–1388. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.; Danzetta, M.L.; di Sabatino, D.; Spedicato, M.; Alkhatal, Z.; Dayhum, A.; Tolari, F.; Forzan, M.; Mazzei, M.; Savini, G. First Seroprevalence Investigation of Epizootic Haemorrhagic Disease Virus in Libya. Open Vet. J. 2021, 11, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Brown-Joseph, T.; Rajko-Nenow, P.; Hicks, H.; Sahadeo, N.; Harrup, L.E.; Carrington, C.V.; Batten, C.; Oura, C.A.L. Identification and Characterization of Epizootic Hemorrhagic Disease Virus Serotype 6 in Cattle Co-Infected with Bluetongue Virus in Trinidad, West Indies. Vet. Microbiol. 2019, 229, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Dommergues, L.; Viarouge, C.; Métras, R.; Youssouffi, C.; Sailleau, C.; Zientara, S.; Cardinale, E.; Cêtre-Sossah, C. Evidence of Bluetongue and Epizootic Haemorrhagic Disease Circulation on the Island of Mayotte. Acta Trop. 2019, 191, 24–28. [Google Scholar] [CrossRef]
- Wilson, W.C.; Ruder, M.G.; Klement, E.; Jasperson, D.C.; Yadin, H.; Stallknecht, D.E.; Mead, D.G.; Howerth, E. Genetic Characterization of Epizootic Hemorrhagic Disease Virus Strains Isolated from Cattle in Israel. J. Gen. Virol. 2015, 96, 1400–1410. [Google Scholar] [CrossRef]
- Murota, K.; Ishii, K.; Mekaru, Y.; Araki, M.; Suda, Y.; Shirafuji, H.; Kobayashi, D.; Isawa, H.; Yanase, T. Isolation of Culicoides- and Mosquito-Borne Orbiviruses in the Southwestern Islands of Japan between 2014 and 2019. Vector Borne Zoonotic Dis. 2021, 21, 796–808. [Google Scholar] [CrossRef]
- Al-Busaidy, S.M.; Mellor, P.S. Epidemiology of Bluetongue and Related Orbiviruses in the Sultanate of Oman. Epidemiol. Infect. 1991, 106, 167–178. [Google Scholar] [CrossRef]
- Gordon, S.J.G.; Bolwell, C.; Rogers, C.W.; Musuka, G.; Kelly, P.; Guthrie, A.; Mellor, P.S.; Hamblin, C. A Serosurvey of Bluetongue and Epizootic Haemorrhagic Disease in a Convenience Sample of Sheep and Cattle Herds in Zimbabwe. Onderstepoort J. Vet. Res. 2017, 84, 1505. [Google Scholar] [CrossRef]
- Toye, P.G.; Batten, C.A.; Kiara, H.; Henstock, M.R.; Edwards, L.; Thumbi, S.; Poole, E.J.; Handel, I.G.; Bronsvoort, B.M.d.C.; Hanotte, O.; et al. Bluetongue and Epizootic Haemorrhagic Disease Virus in Local Breeds of Cattle in Kenya. Res. Vet. Sci. 2013, 94, 769–773. [Google Scholar] [CrossRef] [PubMed]
- Lundervold, M.; Milner-Gulland, E.J.; O’Callaghan, C.J.; Hamblin, C.; Corteyn, A.; Macmillan, A.P. A Serological Survey of Ruminant Livestock in Kazakhstan during Post-Soviet Transitions in Farming and Disease Control. Acta Vet. Scand. 2004, 45, 211. [Google Scholar] [CrossRef] [PubMed]
- Favero, C.M.; Matos, A.C.D.; Campos, F.S.; Cândido, M.V.; Costa, É.A.; Heinemann, M.B.; Barbosa-Stancioli, E.F.; Lobato, Z.I.P. Epizootic Hemorrhagic Disease in Brocket Deer, Brazil. Emerg. Infect. Dis. 2013, 19, 346–348. [Google Scholar] [CrossRef] [PubMed]
- Inaba, Y. Ibaraki Disease and Its Relationship to Bluetongue. Aust. Vet. J. 1975, 51, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Li, Z.; Wang, J.; Li, Z.; Yang, Z.; Liao, D.; Zhu, J.; Li, H. Novel Serotype of Epizootic Hemorrhagic Disease Virus, China. Emerg. Infect. Dis. 2020, 26, 3081–3083. [Google Scholar] [CrossRef]
- Wright, I.M. Serological and Genetic Characterisation of Putative New Serotypes of Bluetongue Virus and Epizootic Haemorrhagic Disease Virus Isolated from an Alpaca. Master’s Thesis, North-West University, Potchefstroom, South Africa, 2014. [Google Scholar]
- Omori, T.; Inaba, Y.; Morimoto, T.; Tanaka, Y.; Ishitani, R. Ibaraki Virus, an Agent of Epizootic Disease of Cattle Resembling Bluetongue. I. Epidemiologic, Clinical and Pathologic Observations and Experimental Transmission to Calves. Jpn. J. Microbiol. 1969, 13, 139–157. [Google Scholar] [CrossRef]
- Garrett, E.F.; Po, E.; Bichi, E.R.; Hexum, S.K.; Melcher, R.; Hubner, A.M. Clinical Disease Associated with Epizootic Hemorrhagic Disease Virus in Cattle in Illinois. J. Am. Vet. Med. Assoc. 2015, 247, 190–195. [Google Scholar] [CrossRef]
- Bréard, E.; Sailleau, C.; Hamblin, C.; Graham, S.D.; Gourreau, J.M.; Zientara, S. Outbreak of Epizootic Haemorrhagic Disease on the Island of Réunion. Vet. Rec. 2004, 155, 422–423. [Google Scholar] [CrossRef]
- Allison, A.B.; Goekjian, V.H.; Potgieter, A.C.; Wilson, W.C.; Johnson, D.J.; Mertens, P.P.C.; Stallknecht, D.E. Detection of a Novel Reassortant Epizootic Hemorrhagic Disease Virus (EHDV) in the USA Containing RNA Segments Derived from Both Exotic (EHDV-6) and Endemic (EHDV-2) Serotypes. J. Gen. Virol. 2010, 91, 430–439. [Google Scholar] [CrossRef]
- Golender, N.; Khinich, Y.; Gorohov, A.; Abramovitz, I.; Bumbarov, V. Epizootic Hemorrhagic Disease Virus Serotype 6 Outbreak in Israeli Cattle in 2015. J. Vet. Diagn. Investig. 2017, 29, 885–888. [Google Scholar] [CrossRef]
- Golender, N.; Bumbarov, V.; Kovtunenko, A.; David, D.; Guini-Rubinstein, M.; Sol, A.; Beer, M.; Eldar, A.; Wernike, K. Identification and Genetic Characterization of Viral Pathogens in Ruminant Gestation Abnormalities, Israel, 2015–2019. Viruses 2021, 13, 2136. [Google Scholar] [CrossRef]
- Behar, A.; Friedgut, O.; Rotenberg, D.; Zalesky, O.; Izhaki, O.; Yulzary, A.; Rot, A.; Wolkomirsky, R.; Zamir, L.; Hmd, F.; et al. Insights on Transmission, Spread, and Possible Endemization of Selected Arboviruses in Israel—Interim Results from Five-Year Surveillance. Vet. Sci. 2022, 9, 65. [Google Scholar] [CrossRef] [PubMed]
- Kedmi, M.; Van Straten, M.; Ezra, E.; Galon, N.; Klement, E. Assessment of the Productivity Effects Associated with Epizootic Hemorrhagic Disease in Dairy Herds. J. Dairy Sci. 2010, 93, 2486–2495. [Google Scholar] [CrossRef]
- Shapiro, J.L.; Wiegers, A.; Dulac, G.C.; Bouffard, A.; Afshar, A.; Myers, D.J.; Dubuc, C.; Martin, M.W.; Koller, M. A Survey of Cattle for Antibodies against Bluetongue and Epizootic Hemorrhagic Disease of Deer Viruses in British Columbia and Southwestern Alberta in 1987. Can. J. Vet. Res. 1991, 55, 203–204. [Google Scholar]
- Allen, S.E.; Rothenburger, J.L.; Jardine, C.M.; Ambagala, A.; Hooper-McGrevy, K.; Colucci, N.; Furukawa-Stoffer, T.; Vigil, S.; Ruder, M.; Nemeth, N.M. Epizootic Hemorrhagic Disease in White-Tailed Deer, Canada. Emerg. Infect. Dis. 2019, 25, 832–834. [Google Scholar] [CrossRef]
- Allen, S.E.; Jardine, C.M.; Hooper-McGrevy, K.; Ambagala, A.; Bosco-Lauth, A.M.; Kunkel, M.R.; Mead, D.G.; Nituch, L.; Ruder, M.G.; Nemeth, N.M. Serologic Evidence of Arthropod-Borne Virus Infections in Wild and Captive Ruminants in Ontario, Canada. Am. J. Trop. Med. Hyg. 2020, 103, 2100–2107. [Google Scholar] [CrossRef] [PubMed]
- Stallknecht, D.E.; Allison, A.B.; Park, A.W.; Phillips, J.E.; Goekjian, V.H.; Nettles, V.F.; Fischer, J.R. Apparent Increase of Reported Hemorrhagic Disease in the Midwestern and Northeastern USA. J. Wildl. Dis. 2015, 51, 348–361. [Google Scholar] [CrossRef]
- Zientara, S.; Bréard, E.; Vitour, D.; Sailleau, C. Emergence of Epizootic Hemorrhagic Disease in Europe. Virologie 2023, 27, 16–17. [Google Scholar] [CrossRef] [PubMed]
- Lorusso, A.; Cappai, S.; Loi, F.; Pinna, L.; Ruiu, A.; Puggioni, G.; Guercio, A.; Purpari, G.; Vicari, D.; Sghaier, S.; et al. First Detection of Epizootic Haemorrhagic Disease Virus in the European Union, Italy-2022. bioRxiv 2022. [Google Scholar] [CrossRef]
- Allison, A.B.; Holmes, E.C.; Potgieter, A.C.; Wright, I.M.; Sailleau, C.; Breard, E.; Ruder, M.G.; Stallknecht, D.E. Segmental Configuration and Putative Origin of the Reassortant Orbivirus, Epizootic Hemorrhagic Disease Virus Serotype 6, Strain Indiana. Virology 2012, 424, 67–75. [Google Scholar] [CrossRef]
- Rodrigues, T.C.S.; Viadanna, P.H.O.; Subramaniam, K.; Hawkins, I.K.; Jeon, A.B.; Loeb, J.C.; Krauer, J.M.C.; Lednicky, J.A.; Wisely, S.M.; Waltzek, T.B. Characterization of a Novel Reassortant Epizootic Hemorrhagic Disease Virus Serotype 6 Strain Isolated from Diseased White-Tailed Deer (Odocoileus virginianus) on a Florida Farm. Viruses 2022, 14, 1012. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lanka, S.; Cassout, D.; Mateus-Pinilla, N.E.; Li, G.; Wilson, W.C.; Yoo, D.; Shelton, P.; Fredrickson, R.L. Inter-Serotype Reassortment among Epizootic Haemorrhagic Disease Viruses in the United States. Transbound. Emerg. Dis. 2019, 66, 1809–1820. [Google Scholar] [CrossRef]
- Anbalagan, S.; Cooper, E.; Klumper, P.; Simonson, R.R.; Hause, B.M. Whole Genome Analysis of Epizootic Hemorrhagic Disease Virus Identified Limited Genome Constellations and Preferential Reassortment. J. Gen. Virol. 2014, 95, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Foster, N.M.; Breckon, R.D.; Luedke, A.J.; Jones, R.H. Transmission of Two Strains of Epizootic Hemorrhagic Disease Virus in Deer by Culicoides variipennis. J. Wildl. Dis. 1977, 13, 9–16. [Google Scholar] [CrossRef]
- Carpenter, S.; Wilson, A.; Barber, J.; Veronesi, E.; Mellor, P.; Venter, G.; Gubbins, S. Temperature Dependence of the Extrinsic Incubation Period of Orbiviruses in Culicoides Biting Midges. PLoS ONE 2011, 6, e27987. [Google Scholar] [CrossRef] [PubMed]
- McGregor, B.L.; Erram, D.; Alto, B.W.; Lednicky, J.A.; Wisely, S.M.; Burkett-Cadena, N.D. Vector Competence of Florida Culicoides Insignis (Diptera: Ceratopogonidae) for Epizootic Hemorrhagic Disease Virus Serotype-2. Viruses 2021, 13, 410. [Google Scholar] [CrossRef] [PubMed]
- Savini, G.; Afonso, A.; Mellor, P.; Aradaib, I.; Yadin, H.; Sanaa, M.; Wilson, W.; Monaco, F.; Domingo, M. Epizootic Haemorragic Disease. Res. Vet. Sci. 2011, 91, 1–17. [Google Scholar] [CrossRef] [PubMed]
- McGregor, B.L.; Shults, P.T.; McDermott, E.G. A Review of the Vector Status of North American Culicoides (Diptera: Ceratopogonidae) for Bluetongue Virus, Epizootic Hemorrhagic Disease Virus, and Other Arboviruses of Concern. Curr. Trop. Med. Rep. 2022, 9, 130–139. [Google Scholar] [CrossRef] [PubMed]
- McGregor, B.L.; Sloyer, K.E.; Sayler, K.A.; Goodfriend, O.; Krauer, J.M.C.; Acevedo, C.; Zhang, X.; Mathias, D.; Wisely, S.M.; Burkett-Cadena, N.D. Field Data Implicating Culicoides Stellifer and Culicoides Venustus (Diptera: Ceratopogonidae) as Vectors of Epizootic Hemorrhagic Disease Virus. Parasites Vectors 2019, 12, 258. [Google Scholar] [CrossRef]
- Whitehorn, J.; Yacoub, S. Global Warming and Arboviral Infections. Clin. Med. 2019, 19, 149–152. [Google Scholar] [CrossRef]
- Rocklöv, J.; Dubrow, R. Climate Change: An Enduring Challenge for Vector-Borne Disease Prevention and Control. Nat. Immunol. 2020, 21, 479–483. [Google Scholar] [CrossRef] [PubMed]
- Climate Change: Global Temperature|NOAA Climate.gov. Available online: http://www.climate.gov/news-features/understanding-climate/climate-change-global-temperature (accessed on 25 March 2023).
- Wittmann, E.J.; Mello, P.S.; Baylis, M. Effect of Temperature on the Transmission of Orbiviruses by the Biting Midge, Culicoides Sonorensis. Med. Vet. Entomol. 2002, 16, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Wellby, M.P.; Baylis, M.; Rawlings, P.; Mellor, P.S. Effect of Temperature on Survival and Rate of Virogenesis of African Horse Sickness Virus in Culicoides variipennis Sonorensis (Diptera: Ceratopogonidae) and Its Significance in Relation to the Epidemiology of the Disease. Bull. Entomol. Res. 1996, 86, 715–720. [Google Scholar] [CrossRef]
- Gao, H.; Wang, L.; Ma, J.; Gao, X.; Xiao, J.; Wang, H. Modeling the Current Distribution Suitability and Future Dynamics of Culicoides Imicola under Climate Change Scenarios. PeerJ 2021, 9, e12308. [Google Scholar] [CrossRef]
- Castillo-Olivares, J. African Horse Sickness in Thailand: Challenges of Controlling an Outbreak by Vaccination. Equine Vet. J. 2021, 53, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Sellers, R.F. Bluetongue, African Horse Sickness, and Related Orbiviruses: Weather, Culicoides, and the Distribution and Spread of Bluetongue and African Horse Sickness Viruses; CRC Press: Boca Raton, FL, USA, 1992; p. 1042. [Google Scholar]
- Hendrickx, G.; Gilbert, M.; Staubach, C.; Elbers, A.; Mintiens, K.; Gerbier, G.; Ducheyne, E. A Wind Density Model to Quantify the Airborne Spread of Culicoides Species during North-Western Europe Bluetongue Epidemic, 2006. Prev. Vet. Med. 2008, 87, 162–181. [Google Scholar] [CrossRef]
- Maurer, L.M.; Paslaru, A.; Torgerson, P.R.; Veronesi, E.; Mathis, A. Vector Competence of Culicoides Biting Midges from Switzerland for African Horse Sickness Virus and Epizootic Haemorrhagic Disease Virus. Schweiz. Arch. Tierheilkd. 2022, 164, 66–70. [Google Scholar] [CrossRef]
- Federici, V.; Ippoliti, C.; Goffredo, M.; Catalani, M.; Di Provvido, A.; Santilli, A.; Quaglia, M.; Mancini, G.; Di Nicola, F.; Di Gennaro, A.; et al. Epizootic Haemorrhagic Disease in Italy: Vector Competence of Indigenous Culicoides Species and Spatial Multicriteria Evaluation of Vulnerability. Vet. Ital. 2016, 52, 271–279. [Google Scholar] [CrossRef]
- Howerth, E.W.; Stallknecht, D.E.; Kirkland, P.D. Bluetongue, Epizootic Hemorrhagic Disease, and Other Orbivirus-Related Diseases. In Infectious Diseases of Wild Mammals; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2001; pp. 77–97. ISBN 978-0-470-34488-0. [Google Scholar]
- Gaydos, J.K.; Davidson, W.R.; Elvinger, F.; Howerth, E.W.; Murphy, M.; Stallknecht, D.E. Cross-Protection between Epizootic Hemorrhagic Disease Virus Serotypes 1 and 2 in White-Tailed Deer. J. Wildl. Dis. 2002, 38, 720–728. [Google Scholar] [CrossRef]
- Brodie, S.J.; Bardsley, K.D.; Diem, K.; Mecham, J.O.; Norelius, S.E.; Wilson, W.C. Epizootic Hemorrhagic Disease: Analysis of Tissues by Amplification and in Situ Hybridization Reveals Widespread Orbivirus Infection at Low Copy Numbers. J. Virol. 1998, 72, 3863–3871. [Google Scholar] [CrossRef]
- Noon, T.H.; Wesche, S.L.; Heffelfinger, J.; Fuller, A.; Bradley, G.A.; Reggiardo, C. Hemorrhagic Disease in Deer in Arizona. J. Wildl. Dis 2002, 38, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Roug, A.; Shannon, J.; Hersey, K.; Heaton, W.; van Wettere, A. Investigation into Causes of Antler Deformities in Mule Deer (Odocoileus hemionus) Bucks in Southern Utah, USA. J. Wildl. Dis. 2022, 58, 222–227. [Google Scholar] [CrossRef]
- Raabis, S.M.; Byers, S.R.; Han, S.; Callan, R.J. Epizootic Hemorrhagic Disease in a Yak. Can. Vet. J. 2014, 55, 369–372. [Google Scholar] [PubMed]
- Kedmi, M.; Levi, S.; Galon, N.; Bomborov, V.; Yadin, H.; Batten, C.; Klement, E. No Evidence for Involvement of Sheep in the Epidemiology of Cattle Virulent Epizootic Hemorrhagic Disease Virus. Vet. Microbiol. 2011, 148, 408–412. [Google Scholar] [CrossRef] [PubMed]
- Nol, P.; Kato, C.; Reeves, W.K.; Rhyan, J.; Spraker, T.; Gidlewski, T.; VerCauteren, K.; Salman, M. Epizootic Hemorrhagic Disease Outbreak in a Captive Facility Housing White-Tailed Deer (Odocoileus virginianus), Bison (Bison bison), Elk (Cervus elaphus), Cattle (Bos taurus), and Goats (Capra hircus) in Colorado, U.S.A. J. Zoo Wildl. Med. 2010, 41, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Mills, M.K.; Ruder, M.G.; Nayduch, D.; Michel, K.; Drolet, B.S. Dynamics of Epizootic Hemorrhagic Disease Virus Infection within the Vector, Culicoides Sonorensis (Diptera: Ceratopogonidae). PLoS ONE 2017, 12, e0188865. [Google Scholar] [CrossRef]
- Darpel, K.E.; Langner, K.F.A.; Nimtz, M.; Anthony, S.J.; Brownlie, J.; Takamatsu, H.-H.; Mellor, P.S.; Mertens, P.P.C. Saliva Proteins of Vector Culicoides Modify Structure and Infectivity of Bluetongue Virus Particles. PLoS ONE 2011, 6, e17545. [Google Scholar] [CrossRef]
- McLaughlin, B.E.; DeMaula, C.D.; Wilson, W.C.; Boyce, W.M.; MacLachlan, N.J. Replication of Bluetongue Virus and Epizootic Hemorrhagic Disease Virus in Pulmonary Artery Endothelial Cells Obtained from Cattle, Sheep, and Deer. Am. J. Vet. Res. 2003, 64, 860–865. [Google Scholar] [CrossRef]
- Sharma, P.; Stallknecht, D.E.; Howerth, E.W. Epizootic Haemorrhagic Disease Virus Induced Apoptosis in Bovine Carotid Artery Endothelium Is P53 Independent. Vet. Ital. 2016, 52, 363–368. [Google Scholar] [CrossRef]
- Shai, B.; Schmukler, E.; Yaniv, R.; Ziv, N.; Horn, G.; Bumbarov, V.; Yadin, H.; Smorodinsky, N.I.; Bacharach, E.; Pinkas-Kramarski, R.; et al. Epizootic Hemorrhagic Disease Virus Induces and Benefits from Cell Stress, Autophagy, and Apoptosis. J. Virol. 2013, 87, 13397–13408. [Google Scholar] [CrossRef]
- Sharma, P.; Stallknecht, D.E.; Murphy, M.D.; Howerth, E.W. Expression of Interleukin-1 Beta and Interleukin-6 in White-Tailed Deer Infected with Epizootic Hemorrhagic Disease Virus. Vet. Ital. 2015, 51, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Howerth, E.W. Cytokine Release and Endothelial Dysfunction: A Perfect Storm in Orbivirus Pathogenesis. Vet. Ital. 2015, 51, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Cordón, P.J.; Pedrera, M.; Risalde, M.A.; Molina, V.; Rodríguez-Sánchez, B.; Núñez, A.; Sánchez-Vizcaíno, J.M.; Gómez-Villamandos, J.C. Potential Role of Proinflammatory Cytokines in the Pathogenetic Mechanisms of Vascular Lesions in Goats Naturally Infected with Bluetongue Virus Serotype 1. Transbound. Emerg. Dis. 2013, 60, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Quist, C.F.; Howerth, E.W.; Stallknecht, D.E.; Brown, J.; Pisell, T.; Nettles, V.F. Host Defense Responses Associated with Experimental Hemorrhagic Disease in White-Tailed Deer. J. Wildl. Dis. 1997, 33, 584–599. [Google Scholar] [CrossRef] [PubMed]
- Wessels, J.E.; Ishida, Y.; Rivera, N.A.; Stirewalt, S.L.; Brown, W.M.; Novakofski, J.E.; Roca, A.L.; Mateus-Pinilla, N.E. The Impact of Variation in the Toll-like Receptor 3 Gene on Epizootic Hemorrhagic Disease in Illinois Wild White-Tailed Deer (Odocoileus virginianus). Genes 2023, 14, 426. [Google Scholar] [CrossRef]
- Stallknecht, D.E.; Howerth, E.W.; Kellogg, M.L.; Quist, C.F.; Pisell, T. In Vitro Replication of Epizootic Hemorrhagic Disease and Bluetongue Viruses in White-Tailed Deer Peripheral Blood Mononuclear Cells and Virus-Cell Association during in Vivo Infections. J. Wildl. Dis. 1997, 33, 574–583. [Google Scholar] [CrossRef]
- Maclachlan, N.J.; Zientara, S.; Wilson, W.C.; Richt, J.A.; Savini, G. Bluetongue and Epizootic Hemorrhagic Disease Viruses: Recent Developments with These Globally Re-Emerging Arboviral Infections of Ruminants. Curr. Opin. Virol. 2019, 34, 56–62. [Google Scholar] [CrossRef]
- Anbalagan, S.; Hause, B.M. Characterization of Epizootic Hemorrhagic Disease Virus from a Bovine with Clinical Disease with High Nucleotide Sequence Identity to White-Tailed Deer Isolates. Arch. Virol. 2014, 159, 2737–2740. [Google Scholar] [CrossRef]
- Gibbs, E.P.J.; Lawman, M.J.P. Infection of British Deer and Farm Animals with Epizootic Haemorrhagic Disease of Deer Virus. J. Comp. Pathol. 1977, 87, 335–343. [Google Scholar] [CrossRef]
- Fletch, A.L.; Karstad, L.H. Studies on the Pathogenesis of Experimental Epizootic Hemorrhagic Disease of White-Tailed Deer. Can. J. Comp. Med. 1971, 35, 224–229. [Google Scholar]
- Shope, R.E.; Macnamara, L.G.; Mangold, R. A virus-induced epizootic hemorrhagic disease of the virginia white-tailed deer (Odocoileus virginianus). J. Exp. Med. 1960, 111, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Ruder, M.G.; Stallknecht, D.E.; Allison, A.B.; Mead, D.G.; Carter, D.L.; Howerth, E.W. Host and Potential Vector Susceptibility to an Emerging Orbivirus in the United States: Epizootic Hemorrhagic Disease Virus Serotype 6. Vet. Pathol. 2016, 53, 574–584. [Google Scholar] [CrossRef] [PubMed]
- Ruder, M.G.; Allison, A.B.; Stallknecht, D.E.; Mead, D.G.; McGraw, S.M.; Carter, D.L.; Kubiski, S.V.; Batten, C.A.; Klement, E.; Howerth, E.W. Susceptibility of White-Tailed Deer (Odocoileus virginianus) to Experimental Infection with Epizootic Hemorrhagic Disease Virus Serotype 7. J. Wildl. Dis. 2012, 48, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Gaydos, J.K.; Davidson, W.R.; Elvinger, F.; Mead, D.G.; Howerth, E.W.; Stallknecht, D.E. Innate Resistance to Epizootic Hemorrhagic Disease in White-Tailed Deer. J. Wildl. Dis. 2002, 38, 713–719. [Google Scholar] [CrossRef]
- Sharma, P.; Stallknech, D.E.; Quist, C.F.; Howerth, E.W. Tumor Necrosis Factor-Alpha Expression in White-Tailed Deer (Odocoileus virginianus) Infected with Epizootic Haemorrhagic Disease Virus. Vet. Ital. 2016, 52, 369–374. [Google Scholar] [CrossRef]
- Gaydos, J.K.; Stallknecht, D.E.; Kavanaugh, D.; Olson, R.J.; Fuchs, E.R. Dynamics of Maternal Antibodies to Hemorrhagic Disease Viruses (Reoviridae: Orbivirus) in White-Tailed Deer. J. Wildl. Dis. 2002, 38, 253–257. [Google Scholar] [CrossRef]
- Stilwell, N.K.; Clarke, L.L.; Howerth, E.W.; Kienzle-Dean, C.; Fojtik, A.; Hollander, L.P.; Carter, D.; Osborn, D.A.; D’Angelo, G.J.; Miller, K.V.; et al. The Effect of Maternal Antibodies on Clinical Response to Infection with Epizootic Hemorrhagic Disease Virus in White-Tailed Deer (Odocoileus virginianus) Fawns. J. Wildl. Dis. 2021, 57, 189–193. [Google Scholar] [CrossRef]
- Quist, C.F.; Howerth, E.W.; Bounous, D.I.; Stallknecht, D.E. Cell-Mediated Immune Response and IL-2 Production in White-Tailed Deer Experimentally Infected with Hemorrhagic Disease Viruses. Vet. Immunol. Immunopathol. 1997, 56, 283–297. [Google Scholar] [CrossRef]
- Ruder, M.G.; Howerth, E.W.; Stallknecht, D.E.; Allison, A.B.; Carter, D.L.; Drolet, B.S.; Klement, E.; Mead, D.G. Vector Competence of Culicoides Sonorensis (Diptera: Ceratopogonidae) to Epizootic Hemorrhagic Disease Virus Serotype 7. Parasites Vectors 2012, 5, 236. [Google Scholar] [CrossRef]
- Smith, K.E.; Stallknecht, D.E.; Nettles, V.F. Experimental Infection of Culicoides Lahillei (Diptera: Ceratopogonidae) with Epizootic Hemorrhagic Disease Virus Serotype 2 (Orbivirus: Reoviridae). J. Med. Entomol. 1996, 33, 117–122. [Google Scholar] [CrossRef]
- Ruder, M.G.; Stallknecht, D.E.; Howerth, E.W.; Carter, D.L.; Pfannenstiel, R.S.; Allison, A.B.; Mead, D.G. Effect of Temperature on Replication of Epizootic Hemorrhagic Disease Viruses in Culicoides Sonorensis (Diptera: Ceratopogonidae). J. Med. Entomol. 2015, 52, 1050–1059. [Google Scholar] [CrossRef] [PubMed]
- Mendiola, S.Y.; Mills, M.K.; Maki, E.; Drolet, B.S.; Wilson, W.C.; Berghaus, R.; Stallknecht, D.E.; Breitenbach, J.; McVey, D.S.; Ruder, M.G. EHDV-2 Infection Prevalence Varies in Culicoides Sonorensis after Feeding on Infected White-Tailed Deer over the Course of Viremia. Viruses 2019, 11, 371. [Google Scholar] [CrossRef] [PubMed]
- Gaydos, J.K.; Allison, A.B.; Hanson, B.A.; Yellin, A.S. Oral and Fecal Shedding of Epizootic Hemorrhagic Disease Virus, Serotype 1 from Experimentally Infected White-Tailed Deer. J. Wildl. Dis. 2002, 38, 166–168. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V.; Cox, R.J.; Waters, W.R.; Thacker, T.C.; Whipple, D.L. Using White-Tailed Deer (Odocoileus virginianus) in Infectious Disease Research. J. Am. Assoc. Lab. Anim. Sci. 2017, 56, 350–360. [Google Scholar] [PubMed]
- Aradaib, I.E.; Sawyer, M.M.; Osburn, B.I. Experimental Epizootic Hemorrhagic Disease Virus Infection in Calves: Virologic and Serologic Studies. J. Vet. Diagn. Investig. 1994, 6, 489–492. [Google Scholar] [CrossRef] [PubMed]
- Bowen, R.A. Serologic Responses of Calves to Sequential Infections with Epizootic Hemorrhagic Disease Virus Serotypes. Am. J. Vet. Res. 1987, 48, 1449–1452. [Google Scholar]
- Abdy, M.J.; Howerth, E.E.; Stallknecht, D.E. Experimental Infection of Calves with Epizootic Hemorrhagic Disease Virus. Am. J. Vet. Res. 1999, 60, 621–626. [Google Scholar]
- Batten, C.A.; Edwards, L.; Bin-Tarif, A.; Henstock, M.R.; Oura, C.A.L. Infection Kinetics of Epizootic Haemorrhagic Disease Virus Serotype 6 in Holstein-Friesian Cattle. Vet. Microbiol. 2011, 154, 23–28. [Google Scholar] [CrossRef]
- Breard, E.; Belbis, G.; Viarouge, C.; Riou, M.; Desprat, A.; Moreau, J.; Laloy, E.; Martin, G.; Sarradin, P.; Vitour, D.; et al. Epizootic Hemorrhagic Disease Virus Serotype 6 Experimentation on Adult Cattle. Res. Vet. Sci. 2013, 95, 794–798. [Google Scholar] [CrossRef]
- Eschbaumer, M.; Wernike, K.; Batten, C.A.; Savini, G.; Edwards, L.; Di Gennaro, A.; Teodori, L.; Oura, C.A.L.; Beer, M.; Hoffmann, B. Epizootic Hemorrhagic Disease Virus Serotype 7 in European Cattle and Sheep: Diagnostic Considerations and Effect of Previous BTV Exposure. Vet. Microbiol. 2012, 159, 298–306. [Google Scholar] [CrossRef]
- Sailleau, C.; Breard, E.; Viarouge, C.; Belbis, G.; Lilin, T.; Vitour, D.; Zientara, S. Experimental Infection of Calves with Seven Serotypes of Epizootic Hemorrhagic Disease Virus: Production and Characterization of Reference Sera. Vet. Ital. 2019, 55, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Hastings, A.K.; Uraki, R.; Gaitsch, H.; Dhaliwal, K.; Stanley, S.; Sproch, H.; Williamson, E.; MacNeil, T.; Marin-Lopez, A.; Hwang, J.; et al. Aedes Aegypti NeSt1 Protein Enhances Zika Virus Pathogenesis by Activating Neutrophils. J. Virol. 2019, 93, e00395-19. [Google Scholar] [CrossRef] [PubMed]
- Ruder, M.G.; Mead, D.G.; Stallknecht, D.E.; Kedmi, M.; Klement, E.; Brown, J.D.; Carter, D.L.; Howerth, E.W. Experimental Infection of Holstein Cows and Calves with EHDV-7 and Preliminary Evaluation of Different Inoculation Methods. Vet. Ital. 2015, 51, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Marín-Lopez, A.; Calvo-Pinilla, E.; Moreno, S.; Utrilla-Trigo, S.; Nogales, A.; Brun, A.; Fikrig, E.; Ortego, J. Modeling Arboviral Infection in Mice Lacking the Interferon Alpha/Beta Receptor. Viruses 2019, 11, 35. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Pinilla, E.; Rodríguez-Calvo, T.; Anguita, J.; Sevilla, N.; Ortego, J. Establishment of a Bluetongue Virus Infection Model in Mice That Are Deficient in the Alpha/Beta Interferon Receptor. PLoS ONE 2009, 4, e5171. [Google Scholar] [CrossRef]
- Marín-López, A.; Bermúdez, R.; Calvo-Pinilla, E.; Moreno, S.; Brun, A.; Ortego, J. Pathological Characterization of IFNAR(-/-) Mice Infected with Bluetongue Virus Serotype 4. Int. J. Biol. Sci. 2016, 12, 1448–1460. [Google Scholar] [CrossRef]
- Castillo-Olivares, J.; Calvo-Pinilla, E.; Casanova, I.; Bachanek-Bankowska, K.; Chiam, R.; Maan, S.; Nieto, J.M.; Ortego, J.; Mertens, P.P.C. A Modified Vaccinia Ankara Virus (MVA) Vaccine Expressing African Horse Sickness Virus (AHSV) VP2 Protects against AHSV Challenge in an IFNAR−/− Mouse Model. PLoS ONE 2011, 6, e16503. [Google Scholar] [CrossRef]
- Jiménez-Cabello, L.; Utrilla-Trigo, S.; Calvo-Pinilla, E.; Moreno, S.; Nogales, A.; Ortego, J.; Marín-López, A. Viral Vector Vaccines against Bluetongue Virus. Microorganisms 2021, 9, 42. [Google Scholar] [CrossRef]
- Calvo-Pinilla, E.; Marín-López, A.; Moreno, S.; Lorenzo, G.; Utrilla-Trigo, S.; Jiménez-Cabello, L.; Benavides, J.; Nogales, A.; Blasco, R.; Brun, A.; et al. A Protective Bivalent Vaccine against Rift Valley Fever and Bluetongue. NPJ Vaccines 2020, 5, 70. [Google Scholar] [CrossRef]
- Utrilla-Trigo, S.; Jiménez-Cabello, L.; Alonso-Ravelo, R.; Calvo-Pinilla, E.; Marín-López, A.; Moreno, S.; Lorenzo, G.; Benavides, J.; Gilbert, S.; Nogales, A.; et al. Heterologous Combination of ChAdOx1 and MVA Vectors Expressing Protein NS1 as Vaccination Strategy to Induce Durable and Cross-Protective CD8+ T Cell Immunity to Bluetongue Virus. Vaccines 2020, 8, 346. [Google Scholar] [CrossRef]
- Utrilla-Trigo, S.; Jiménez-Cabello, L.; Calvo-Pinilla, E.; Marín-López, A.; Lorenzo, G.; Sánchez-Cordón, P.; Moreno, S.; Benavides, J.; Gilbert, S.; Nogales, A.; et al. The Combined Expression of the Non-Structural Protein NS1 and the N-Terminal Half of NS2 (NS21-180) by ChAdOx1 and MVA Confers Protection against Clinical Disease in Sheep upon Bluetongue Virus Challenge. J. Virol. 2021, 96, e01614-21. [Google Scholar] [CrossRef] [PubMed]
- Mettler, N.E.; Macnamara, L.G.; Shope, R.E. The Propagation of the Virus of Epizootic Hemorrhagic Disease of Deer in Newborn Mice and HeLa Cells. J. Exp. Med. 1962, 116, 665–678. [Google Scholar] [CrossRef] [PubMed]
- Shope, R.E.; MacNamara, L.G.; Mettler, N.E. The Attenuation of The Virus of Epizootic Hemorrhagic Disease of Deer by Its Serial Passage in the Brains of Newborn Mice. J. Exp. Med. 1963, 118, 421–424. [Google Scholar] [CrossRef]
- Willems, F.; Vollstedt, S.; Suter, M. Phenotype and Function of Neonatal DC. Eur. J. Immunol. 2009, 39, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Eschbaumer, M.; Keller, M.; Beer, M.; Hoffmann, B. Epizootic Hemorrhagic Disease Virus Infection of Type I Interferon Receptor Deficient Mice. Vet. Microbiol. 2012, 155, 417–419. [Google Scholar] [CrossRef]
- Spedicato, M.; Carmine, I.; Teodori, L.; Leone, A.; Portanti, O.; Marini, V.; Pisciella, M.; Lorusso, A.; Savini, G. Innocuity of a Commercial Live Attenuated Vaccine for Epizootic Hemorrhagic Disease Virus Serotype 2 in Late-Term Pregnant Cows. Vaccine 2016, 34, 1430–1435. [Google Scholar] [CrossRef]
- McVey, D.S.; MacLachlan, N.J. Vaccines for Prevention of Bluetongue and Epizootic Hemorrhagic Disease in Livestock: A North American Perspective. Vector Borne Zoonotic Dis. 2015, 15, 385–396. [Google Scholar] [CrossRef]
- van Rijn, P.A. Prospects of Next-Generation Vaccines for Bluetongue. Front. Vet. Sci. 2019, 6, 407. [Google Scholar] [CrossRef]
- Patta, C.; Giovannini, A.; Rolesu, S.; Nannini, D.; Savini, G.; Calistri, P.; Santucci, U.; Caporale, V. Bluetongue Vaccination in Europe: The Italian Experience. Vet. Ital. 2004, 40, 601–610. [Google Scholar]
- Sunwoo, S.Y.; Noronha, L.E.; Morozov, I.; Trujillo, J.D.; Kim, I.J.; Schirtzinger, E.E.; Faburay, B.; Drolet, B.S.; Urbaniak, K.; McVey, D.S.; et al. Evaluation of A Baculovirus-Expressed VP2 Subunit Vaccine for the Protection of White-Tailed Deer (Odocoileus virginianus) from Epizootic Hemorrhagic Disease. Vaccines 2020, 8, 59. [Google Scholar] [CrossRef]
- Jiménez-Cabello, L.; Utrilla-Trigo, S.; Barreiro-Piñeiro, N.; Pose-Boirazian, T.; Martínez-Costas, J.; Marín-López, A.; Ortego, J. Nanoparticle- and Microparticle-Based Vaccines against Orbiviruses of Veterinary Importance. Vaccines 2022, 10, 1124. [Google Scholar] [CrossRef] [PubMed]
- Alshaikhahmed, K.; Roy, P. Generation of Virus-like Particles for Emerging Epizootic Haemorrhagic Disease Virus: Towards the Development of Safe Vaccine Candidates. Vaccine 2016, 34, 1103–1108. [Google Scholar] [CrossRef]
- Fay, P.C.; Mohd Jaafar, F.; Batten, C.; Attoui, H.; Saunders, K.; Lomonossoff, G.P.; Reid, E.; Horton, D.; Maan, S.; Haig, D.; et al. Serological Cross-Reactions between Expressed VP2 Proteins from Different Bluetongue Virus Serotypes. Viruses 2021, 13, 1455. [Google Scholar] [CrossRef] [PubMed]
- Forzan, M.; Maan, S.; Mazzei, M.; Belaganahalli, M.N.; Bonuccelli, L.; Calamari, M.; Carrozza, M.L.; Cappello, V.; Di Luca, M.; Bandecchi, P.; et al. Generation of Virus like Particles for Epizootic Hemorrhagic Disease Virus. Res. Vet. Sci. 2016, 107, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Zhang, J.; Xu, Q.; Sun, E.; Li, J.; Lv, S.; Feng, Y.; Zhang, Q.; Wang, H.; Wang, H.; et al. Development of a Reverse Genetics System for Epizootic Hemorrhagic Disease Virus and Evaluation of Novel Strains Containing Duplicative Gene Rearrangements. J. Gen. Virol. 2015, 96, 2714–2720. [Google Scholar] [CrossRef]
- Matsuo, E.; Saeki, K.; Roy, P.; Kawano, J. Development of Reverse Genetics for Ibaraki Virus to Produce Viable VP6-Tagged IBAV. FEBS Open Bio 2015, 5, 445–453. [Google Scholar] [CrossRef]
- van Rijn, P.A.; Maris-Veldhuis, M.A.; van Gennip, R.G.P. The Bluetongue Disabled Infectious Single Animal (DISA) Vaccine Platform Based on Deletion NS3/NS3a Protein Is Safe and Protective in Cattle and Enables DIVA. Viruses 2021, 13, 857. [Google Scholar] [CrossRef]
- Calvo-Pinilla, E.; Marín-López, A.; Utrilla-Trigo, S.; Jiménez-Cabello, L.; Ortego, J. Reverse Genetics Approaches: A Novel Strategy for African Horse Sickness Virus Vaccine Design. Curr. Opin. Virol. 2020, 44, 49–56. [Google Scholar] [CrossRef]
- van Rijn, P.A.; Maris-Veldhuis, M.A.; Boonstra, J.; van Gennip, R.G.P. Diagnostic DIVA Tests Accompanying the Disabled Infectious Single Animal (DISA) Vaccine Platform for African Horse Sickness. Vaccine 2018, 36, 3584–3592. [Google Scholar] [CrossRef]
- Tacken, M.G.J.; Daus, F.J.; Feenstra, F.; van Gennip, R.G.P.; van Rijn, P.A. Development of a Competitive ELISA for NS3 Antibodies as DIVA Test Accompanying the Novel Disabled Infectious Single Animal (DISA) Vaccine for Bluetongue. Vaccine 2015, 33, 5539–5545. [Google Scholar] [CrossRef]
- Matsuo, E.; Celma, C.C.P.; Boyce, M.; Viarouge, C.; Sailleau, C.; Dubois, E.; Bréard, E.; Thiéry, R.; Zientara, S.; Roy, P. Generation of Replication-Defective Virus-Based Vaccines That Confer Full Protection in Sheep against Virulent Bluetongue Virus Challenge. J. Virol. 2011, 85, 10213–10221. [Google Scholar] [CrossRef] [PubMed]
- van Rijn, P.A.; Maris-Veldhuis, M.A.; Spedicato, M.; Savini, G.; van Gennip, R.G.P. Pentavalent Disabled Infectious Single Animal (DISA)/DIVA Vaccine Provides Protection in Sheep and Cattle against Different Serotypes of Bluetongue Virus. Vaccines 2021, 9, 1150. [Google Scholar] [CrossRef] [PubMed]
- Celma, C.C.; Stewart, M.; Wernike, K.; Eschbaumer, M.; Gonzalez-Molleda, L.; Breard, E.; Schulz, C.; Hoffmann, B.; Haegeman, A.; De Clercq, K.; et al. Replication-Deficient Particles: New Insights into the Next Generation of Bluetongue Virus Vaccines. J. Virol. 2016, 91, e01892-16. [Google Scholar] [CrossRef] [PubMed]
- Celma, C.C.P.; Boyce, M.; van Rijn, P.A.; Eschbaumer, M.; Wernike, K.; Hoffmann, B.; Beer, M.; Haegeman, A.; De Clercq, K.; Roy, P. Rapid Generation of Replication-Deficient Monovalent and Multivalent Vaccines for Bluetongue Virus: Protection against Virulent Virus Challenge in Cattle and Sheep. J. Virol. 2013, 87, 9856–9864. [Google Scholar] [CrossRef] [PubMed]
- Russell, B.L.; Parbhoo, N.; Gildenhuys, S. Analysis of Conserved, Computationally Predicted Epitope Regions for VP5 and VP7 Across Three Orbiviruses. Bioinform. Biol. Insights 2018, 12, 1177932218755348. [Google Scholar] [CrossRef]
- Armillotta, G.; Di Febo, T.; Ulisse, S.; Laguardia, C.; Iorio, M.; Krasteva, I.; Tittarelli, M.; Mercante, M.T.; Luciani, M. Production and Characterization of Monoclonal Antibodies against the VP7 Protein of Epizootic Hemorrhagic Disease Virus. Monoclon. Antib. Immunodiagn. Immunother. 2022, 41, 181–187. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez-Cabello, L.; Utrilla-Trigo, S.; Lorenzo, G.; Ortego, J.; Calvo-Pinilla, E. Epizootic Hemorrhagic Disease Virus: Current Knowledge and Emerging Perspectives. Microorganisms 2023, 11, 1339. https://doi.org/10.3390/microorganisms11051339
Jiménez-Cabello L, Utrilla-Trigo S, Lorenzo G, Ortego J, Calvo-Pinilla E. Epizootic Hemorrhagic Disease Virus: Current Knowledge and Emerging Perspectives. Microorganisms. 2023; 11(5):1339. https://doi.org/10.3390/microorganisms11051339
Chicago/Turabian StyleJiménez-Cabello, Luis, Sergio Utrilla-Trigo, Gema Lorenzo, Javier Ortego, and Eva Calvo-Pinilla. 2023. "Epizootic Hemorrhagic Disease Virus: Current Knowledge and Emerging Perspectives" Microorganisms 11, no. 5: 1339. https://doi.org/10.3390/microorganisms11051339
APA StyleJiménez-Cabello, L., Utrilla-Trigo, S., Lorenzo, G., Ortego, J., & Calvo-Pinilla, E. (2023). Epizootic Hemorrhagic Disease Virus: Current Knowledge and Emerging Perspectives. Microorganisms, 11(5), 1339. https://doi.org/10.3390/microorganisms11051339