
In Vitro Biofilm-Mediated Biodegradation of Pesticides and Dye-Contaminated Effluents Using Bacterial Biofilms


 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Pollutants
2.2. Soil Sampling and Isolation of Bacterial Strains
2.3. Congo Red Assay (Qualitative Biofilm Production Assay)
2.4. Morphological and Biochemical Characterization
2.5. Genetic Identification of Bacterial Strains
2.6. Biofilm Time Kinetics
2.7. Biofilm-Mediated Biodegradation Assay
2.8. Fourier Transform Infrared Spectroscopy
2.9. Thin-Layer Chromatography(TLC)
2.10. Statistical Analysis
3. Results
3.1. Screening of Biofilm Producers
3.2. Morphological and Biochemical Identification
3.3. 16S rRNA Sequencing Study
3.4. Time Kinetics of Biofilm Formation
3.5. UV–VIS Spectroscopic Analysis of CYP and IMI before and after Biofilm Treatment
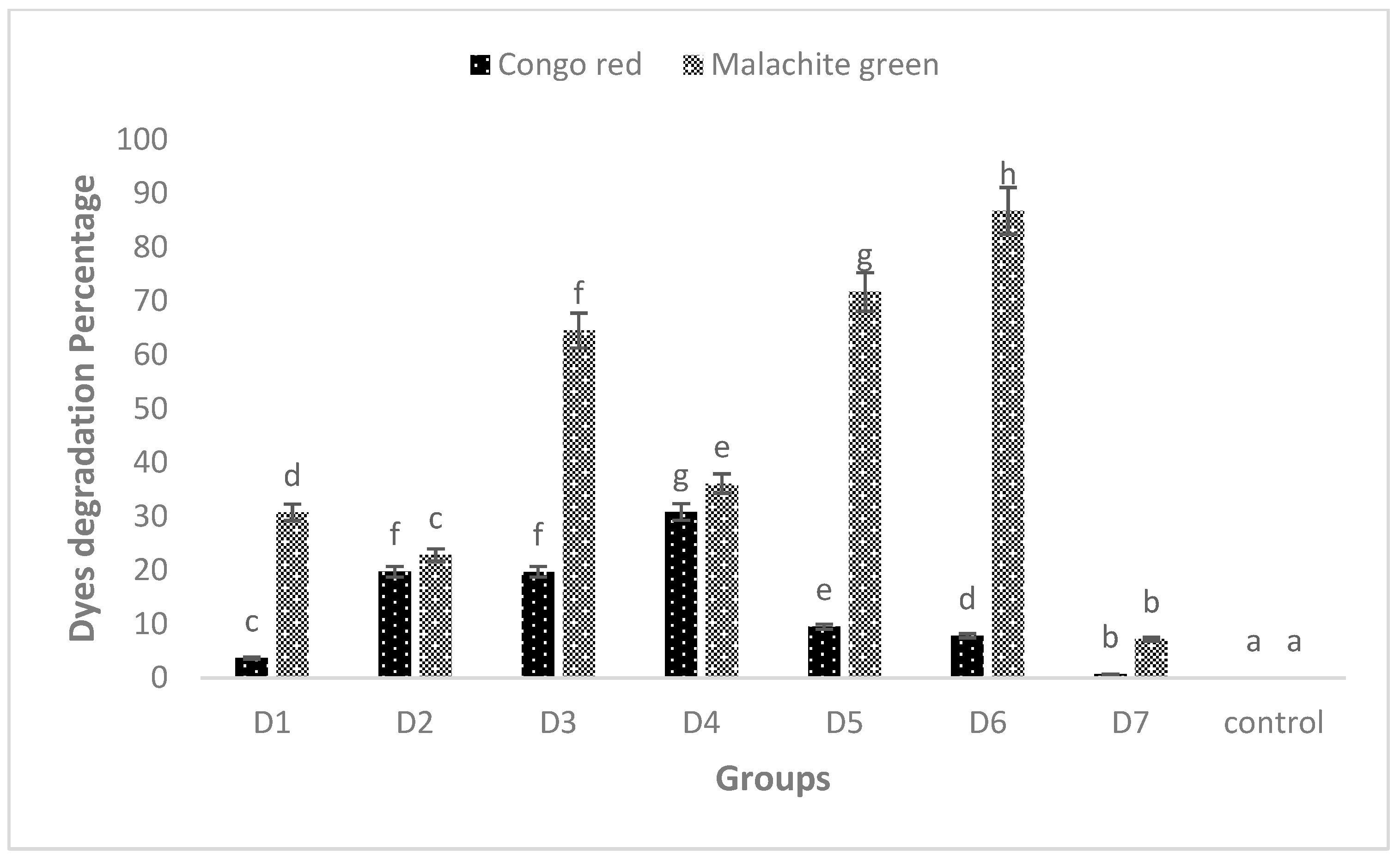
3.6. UV–VIS Spectroscopic Analysis of MG and CR before and after Biofilm Treatment
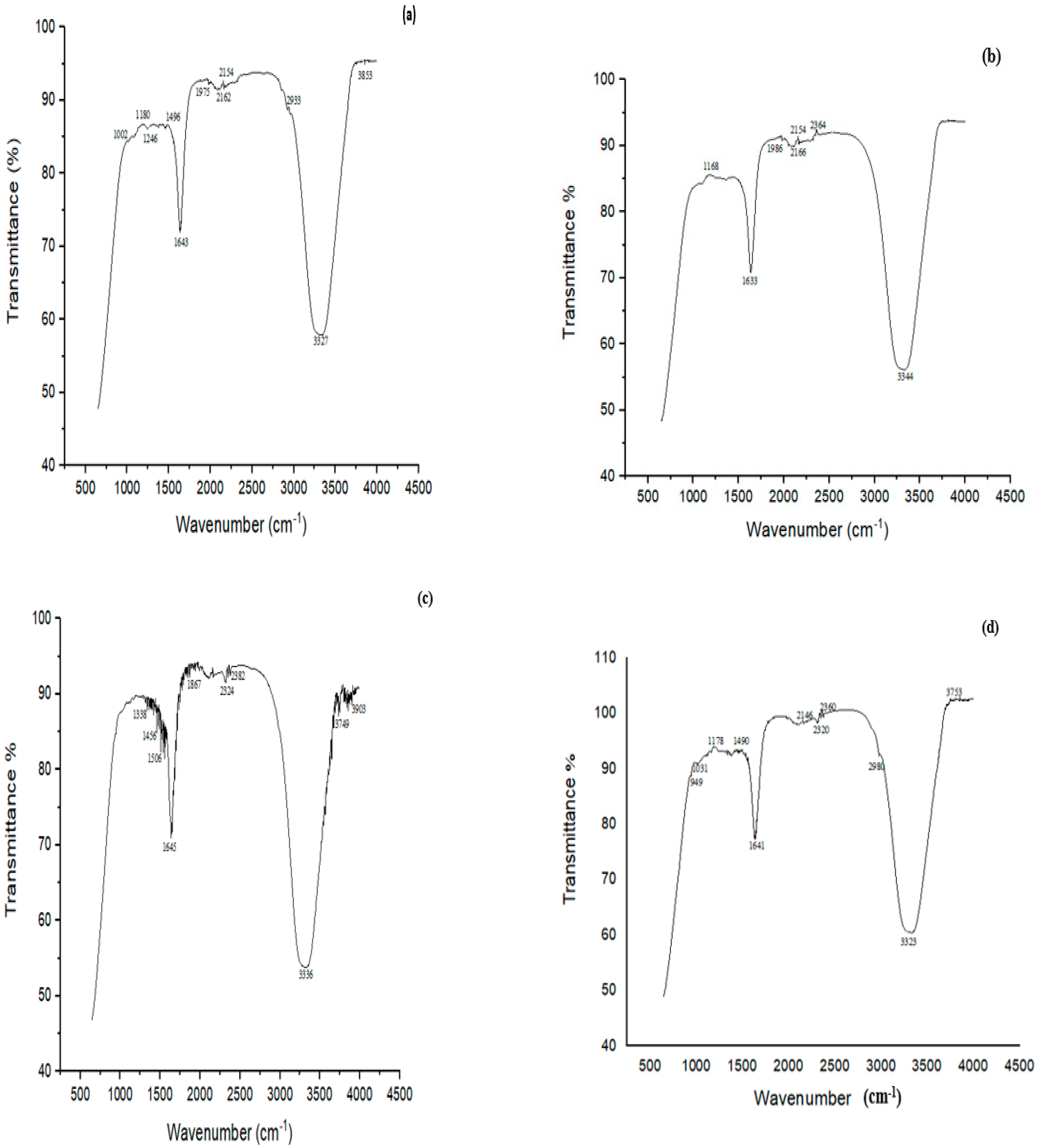
3.7. FTIR Analysis of CYP and IMI before and after Biodegradation
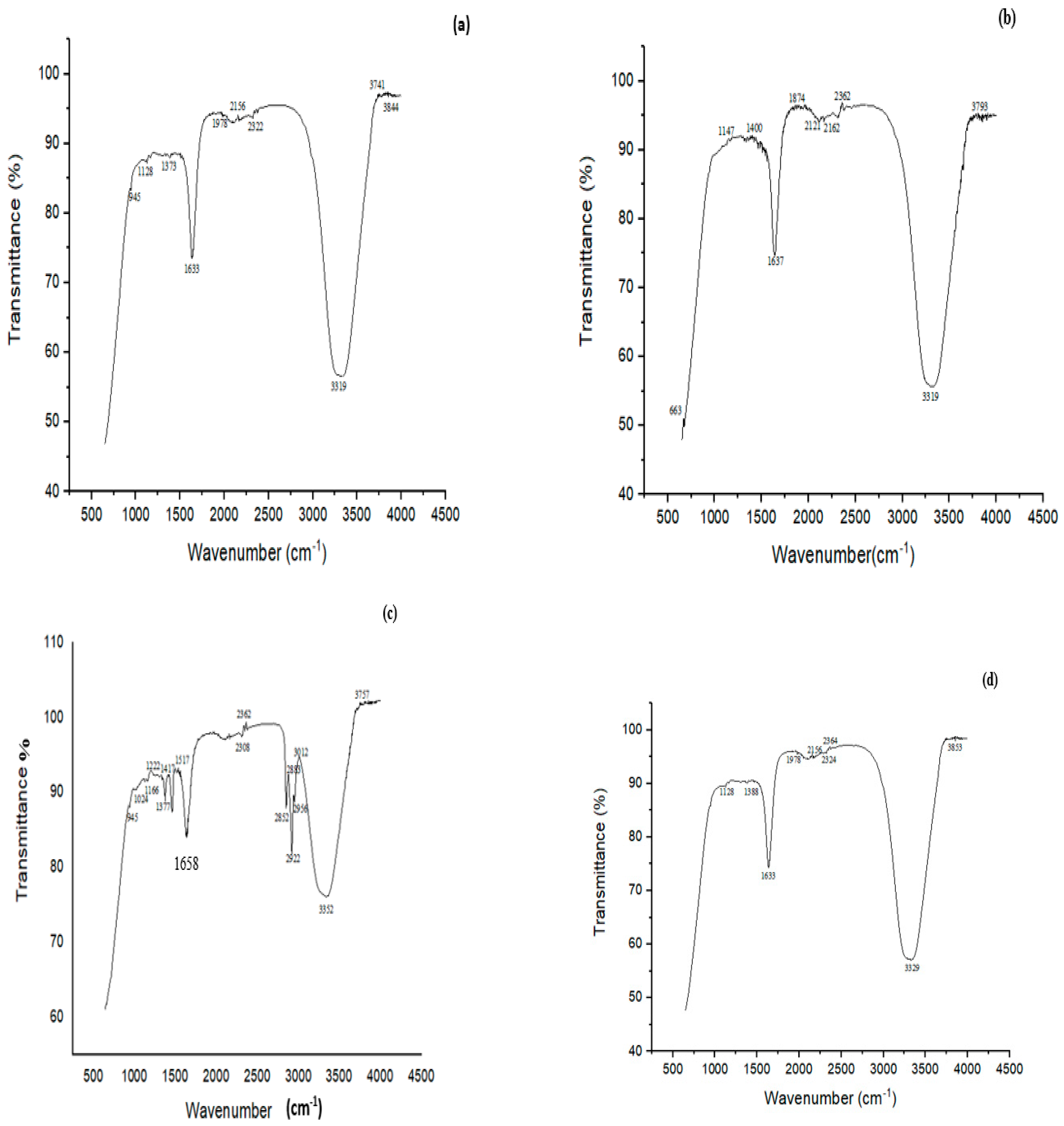
3.8. FTIR Analysis of MG and CR before and after Biodegradation
3.9. TLC Analysis of CYP, IMI, MG and CR before and after Biodegradation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khalid, M.Z.; Ahmed, S.; Al-Ashkar, I.; El Sabagh, A.; Liu, L.; Zhong, G. Evaluation of resistance development in Bemisia tabaci Genn.(Homoptera: Aleyrodidae) in cotton against different insecticides. Insects 2021, 12, 996. [Google Scholar] [CrossRef] [PubMed]
- Hayat, N.; Hussain, A.; Lohano, H.D. Eco-labeling and sustainability: A case of textile industry in Pakistan. J. Clean. Prod. 2020, 252, 119807. [Google Scholar] [CrossRef]
- Barathi, S.; Aruljothi, K.; Karthik, C.; Padikasan, I.A.; Ashokkumar, V. Biofilm mediated decolorization and degradation of reactive red 170 dye by the bacterial consortium isolated from the dyeing industry wastewater sediments. Chemosphere 2022, 286, 131914. [Google Scholar] [CrossRef] [PubMed]
- Talha, M.A.; Goswami, M.; Giri, B.; Sharma, A.; Rai, B.; Singh, R. Bioremediation of Congo red dye in immobilized batch and continuous packed bed bioreactor by Brevibacillus parabrevis using coconut shell bio-char. Bioresour. Technol. 2018, 252, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Tariq, M.I.; Afzal, S.; Hussain, I.; Sultana, N. Pesticides exposure in Pakistan: A review. Environ. Int. 2007, 33, 1107–1122. [Google Scholar] [CrossRef] [PubMed]
- Razaq, M.; Suhail, A.; Arif, M.; Aslam, M.; Sayyed, A. Effect of rotational use of insecticides on pyrethroids resistance in Helicoverpa armigera (Lep.: Noctuidae). J. Appl. Entomol. 2007, 131, 460–465. [Google Scholar] [CrossRef]
- Whangchai, K.; Van Hung, T.; Al-Rashed, S.; Narayanan, M.; Kandasamy, S.; Pugazhendhi, A. Biodegradation competence of Streptomyces toxytricini D2 isolated from leaves surface of the hybrid cotton crop against β cypermethrin. Chemosphere 2021, 276, 130152. [Google Scholar] [CrossRef]
- Huang, Y.; Xiao, L.; Li, F.; Xiao, M.; Lin, D.; Long, X.; Wu, Z. Microbial degradation of pesticide residues and an emphasis on the degradation of cypermethrin and 3-phenoxy benzoic acid: A review. Molecules 2018, 23, 2313. [Google Scholar] [CrossRef]
- Ali, H.F. Cellular Mechanism involved in cypermethrin induced neurotoxicity. Rec. Pharm. Biomed. Sci. 2020, 4, 32–39. [Google Scholar] [CrossRef]
- Singh, Z.; Kaur, J.; Kaur, R.; Hundal, S.S. Toxic effects of organochlorine pesticides: A review. Am. J. BioSci. 2016, 4, 11. [Google Scholar] [CrossRef]
- Phugare, S.S.; Kalyani, D.C.; Gaikwad, Y.B.; Jadhav, J.P. Microbial degradation of imidacloprid and toxicological analysis of its biodegradation metabolites in silkworm (Bombyx mori). Chem. Eng. J. 2013, 230, 27–35. [Google Scholar] [CrossRef]
- Baig, S.A.; Akhter, N.A.; Ashfaq, M.; Asi, M.R.; Ashfaq, U. Imidacloprid residues in vegetables, soil and water in the southern Punjab, Pakistan. J. Agric. Technol. 2012, 8, 903–916. [Google Scholar]
- Rafique, N.; Tariq, S.R.; Ahmed, D. Monitoring and distribution patterns of pesticide residues in soil from cotton/wheat fields of Pakistan. Environ. Monit. Assess. 2016, 188, 695. [Google Scholar] [CrossRef]
- Pang, S.; Lin, Z.; Zhang, W.; Mishra, S.; Bhatt, P.; Chen, S. Insights into the microbial degradation and biochemical mechanisms of neonicotinoids. Front. Microbiol. 2020, 11, 868. [Google Scholar] [CrossRef]
- Noman, M.; Shahid, M.; Ahmed, T.; Niazi, M.B.K.; Hussain, S.; Song, F.; Manzoor, I. Use of biogenic copper nanoparticles synthesized from a native Escherichia sp. as photocatalysts for azo dye degradation and treatment of textile effluents. Environ. Pollut. 2020, 257, 113514. [Google Scholar] [CrossRef] [PubMed]
- Daneshvar, N.; Khataee, A.; Rasoulifard, M.; Pourhassan, M. Biodegradation of dye solution containing Malachite Green: Optimization of effective parameters using Taguchi method. J. Hazard. Materi. 2007, 143, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Parshetti, G.; Kalme, S.; Saratale, G.; Govindwar, S. Biodegradation of Malachite Green by Kocuria rosea MTCC 1532. Acta Chim. Slov. 2006, 53, 492–498. [Google Scholar]
- Alaya, V.; Kodi, R.K.; Ninganna, E.; Gowda, B.; Shivanna, M. Decolorization of Malachite green dye by Stenotrophomonas maltophilia a compost bacterium. Bull. Natl. Res. Cent. 2021, 45, 81. [Google Scholar] [CrossRef]
- Tatarko, M.; Bumpus, J.A. Biodegradation of congo red by Phanerochaete chrysosporium. Water Res. 1998, 32, 1713–1717. [Google Scholar] [CrossRef]
- Rai, M.S.; Bhat, P.R.; Prajna, P.; Jayadev, K.; Rao, P.V. Degradation of malachite green and congo red using Aloe barabadensis mill extract. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 330–340. [Google Scholar]
- Adane, T.; Adugna, A.T.; Alemayehu, E. Textile industry effluent treatment techniques. J. Chem. 2021, 2021, 5314404. [Google Scholar] [CrossRef]
- Selvaraj, V.; Karthika, T.S.; Mansiya, C.; Alagar, M. An over review on recently developed techniques, mechanisms and intermediate involved in the advanced azo dye degradation for industrial applications. J. Mol. Struct. 2021, 1224, 129195. [Google Scholar] [CrossRef]
- Dash, D.M.; Osborne, W.J. Rapid biodegradation and biofilm-mediated bioremoval of organophosphorus pesticides using an indigenous Kosakonia oryzae strain-VITPSCQ3 in a Vertical-flow Packed Bed Biofilm Bioreactor. Ecotoxicol. Environ. Saf. 2020, 192, 110290. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.M.; Haque, M.A.; Mosharaf, M.K.; Islam, M.S.; Islam, M.M.; Hasan, M.; Molla, A.H.; Haque, M.A. Biofilm-mediated decolorization, degradation and detoxification of synthetic effluent by novel biofilm-producing bacteria isolated from textile dyeing effluent. Environ. Pollut. 2022, 314, 120237. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, J.; Thakur, A.; Goyal, A. Industrial Wastewater and Its Toxic Effects; Royal Society of Chemistry: London, UK, 2021. [Google Scholar]
- Maurya, A.; Raj, A. Recent advances in the application of biofilm in bioremediation of industrial wastewater and organic pollutants. In Microorganisms for Sustainable Environment and Health; Springer: Berlin/Heidelberg, Germany, 2020; pp. 81–118. [Google Scholar]
- Sharma, S.; Singh, B.; Gupta, V. Assessment of imidacloprid degradation by soil-isolated Bacillus alkalinitrilicus. Environ. Monit. Assess. 2014, 186, 7183–7193. [Google Scholar] [CrossRef]
- Gupta, M.; Mathur, S.; Sharma, T.K.; Rana, M.; Gairola, A.; Navani, N.K.; Pathania, R. A study on metabolic prowess of Pseudomonas sp. RPT 52 to degrade imidacloprid, endosulfan and coragen. J. Hazard. Mater. 2016, 301, 250–258. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, Y.; Chen, W.-J.; Wu, S.; Lei, Q.; Zhou, Z.; Zhang, W.; Mishra, S.; Bhatt, P.; Chen, S. Environmental occurrence, toxicity concerns, and biodegradation of neonicotinoid insecticides. Environ. Res. 2023, 218, 114953. [Google Scholar] [CrossRef]
- Gautam, P.; Pandey, A.K.; Dubey, S.K. Multi-omics approach reveals elevated potential of bacteria for biodegradation of imidacloprid. Environ. Res. 2023, 221, 115271. [Google Scholar] [CrossRef]
- Gautam, P.; Pandey, A.K.; Gupta, A.; Dubey, S.K. Microcosm-omics centric investigation reveals elevated bacterial degradation of imidacloprid. Environ. Pollut. 2023, 324, 121402. [Google Scholar] [CrossRef]
- Sharma, A.; Gangola, S.; Khati, P.; Kumar, G.; Srivastava, A. Novel pathway of cypermethrin biodegradation in a Bacillus sp. strain SG2 isolated from cypermethrin-contaminated agriculture field. 3 Biotech 2016, 6, 45. [Google Scholar]
- Qu, W.; Hong, G.; Zhao, J. Degradation of malachite green dye by Tenacibaculum sp. HMG1 isolated from Pacific deep-sea sediments. Acta Oceanol. Sin. 2018, 37, 104–111. [Google Scholar] [CrossRef]
- Kishor, R.; Purchase, D.; Saratale, G.D.; Ferreira, L.F.R.; Bilal, M.; Iqbal, H.M.; Bharagava, R.N. Environment friendly degradation and detoxification of Congo red dye and textile industry wastewater by a newly isolated Bacillus cohnni (RKS9). Environ. Technol. Innov. 2021, 22, 101425. [Google Scholar] [CrossRef]
- Firoozeh, F.; Shahamat, Y.D.; Rodríguez-Couto, S.; Kouhsari, E.; Niknejad, F. Bioremediation for the Decolorization of Textile Dyes by Bacterial Strains Isolated from Dyeing Wastewater. Jordan J. Biolol. Sci. 2022, 15, 219–225. [Google Scholar]
- Liaqat, I.; Bachmann, R.T.; Edyvean, R.G. Type 2 quorum sensing monitoring, inhibition and biofilm formation in marine microrganisms. Curr. Microbiol. 2014, 68, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Liaqat, I.; Muhammad, N.; Ara, C.; Hanif, U.; Andleeb, S.; Arshad, M.; Aftab, M.N.; Raza, C.; Mubin, M. Bioremediation of heavy metals polluted environment and decolourization of black liquor using microbial biofilms. Mol. Biol. Rep. 2023, 50, 3985–3997. [Google Scholar] [CrossRef] [PubMed]
- Holt, J.G.; Krieg, N.R.; Sneath, P.H.; Staley, J.T.; Williams, S.T. Bergey’s Manual of Determinate Bacteriology. 1994. Available online: https://bsu.libguides.com/c.php?g=41359&p=263398 (accessed on 13 March 2022.).
- Zhi, X.-Y.; Li, W.-J.; Stackebrandt, E. An update of the structure and 16S rRNA gene sequence-based definition of higher ranks of the class Actinobacteria, with the proposal of two new suborders and four new families and emended descriptions of the existing higher taxa. Int. J. Syst. Evol. Microbiol. 2009, 59, 589–608. [Google Scholar] [CrossRef]
- Liaqat, I.; Sumbal, F.; Sabri, A.N. Tetracycline and chloramphenicol efficiency against selected biofilm forming bacteria. Curr. Microbiol. 2009, 59, 212–220. [Google Scholar] [CrossRef]
- Narayanan, M.; Murugan, J.M.; Kandasamy, G.; Kandasamy, S.; Nasif, O.; Rajendran, M.; Pugazhendhi, A. The biotransformation potential of Bacillus cereus on β-cypermethrin to protect the earthworm (Perionyx excavatus) on insecticide-contaminated soil. Arch. Agron. Soil Sci. 2020, 68, 944–955. [Google Scholar] [CrossRef]
- Tahir, U.; Nawaz, S.; Hassan, K.U.; Yasmin, A. Assessment of biodecolorization potentials of biofilm forming bacteria from two different genera for Mordant Black 11 dye. Bioremed. J. 2021, 25, 252–270. [Google Scholar] [CrossRef]
- Kalyani, D.; Patil, P.S.; Jadhav, J.P.; Govindwar, S.P. Biodegradation of reactive textile dye Red BLI by an isolated bacterium Pseudomonas sp. SUK1. Bioresour. Technol. 2008, 99, 4635–4641. [Google Scholar] [CrossRef]
- Sandhya, M.; Huang, Y.; Li, J.; Wu, X.; Zhou, Z.; Lei, Q.; Bhatt, P.; Chen, S. Biofilm-mediated bioremediation is a powerful tool for the removal of environmental pollutants. Chemosphere 2022, 294, 133609. [Google Scholar]
- Zhang, L.; Dong, D.; Hua, X.; Guo, Z. Inhibitory effects of extracellular polymeric substances on ofloxacin sorption by natural biofilms. Sci. Total Environ. 2018, 625, 178–184. [Google Scholar] [CrossRef]
- Ambalam, P.; Kondepudi, K.K.; Nilsson, I.; Wadström, T.; Ljungh, Å. Bile stimulates cell surface hydrophobicity, Congo red binding and biofilm formation of Lactobacillus strains. FEMS Microbiol. Lett. 2012, 333, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Liaqat, I.; Bachmann, R.T.; Sabri, A.N.; Edyvean, R.G. Isolate-specific effects of patulin, penicillic Acid and EDTA on biofilm formation and growth of dental unit water line biofilm isolates. Curr. Microbiol. 2010, 61, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Sarwan, J. Role of isolates of Bacillus species for biodegradation of multiple contaminants. J. Sustain. Environ. Manag. 2022, 1, 292–298. [Google Scholar] [CrossRef]
- Hossen, M.Z.; Hussain, M.E.; Hakim, A.; Islam, K.; Uddin, M.N.; Azad, A.K. Biodegradation of reactive textile dye Novacron Super Black G by free cells of newly isolated Alcaligenes faecalis AZ26 and Bacillus spp. obtained from textile effluents. Heliyon 2019, 5, e02068. [Google Scholar] [CrossRef]
- Chen, S.; Deng, Y.; Chang, C.; Lee, J.; Cheng, Y.; Cui, Z.; Zhou, J.; He, F.; Hu, M.; Zhang, L.-H. Pathway and kinetics of cyhalothrin biodegradation by Bacillus thuringiensis strain ZS-19. Sci. Rep. 2015, 5, srep08784. [Google Scholar] [CrossRef]
- Xiao, Y.; Chen, S.; Gao, Y.; Hu, W.; Hu, M.; Zhong, G. Isolation of a novel beta-cypermethrin degrading strain Bacillus subtilis BSF01 and its biodegradation pathway. Appl. Microbiol. Biotechnol. 2015, 99, 2849–2859. [Google Scholar] [CrossRef]
- Nour, E.H.; Elsayed, T.R.; Springael, D.; Smalla, K. Comparable dynamics of linuron catabolic genes and IncP-1 plasmids in biopurification systems (BPSs) as a response to linuron spiking. Appl. Microbiol. Biotechnol. 2017, 101, 4815–4825. [Google Scholar] [CrossRef]
- Gangola, S.; Sharma, A.; Joshi, S.; Bhandari, G.; Prakash, O.; Govarthanan, M.; Kim, W.; Bhatt, P. Novel mechanism and degradation kinetics of pesticides mixture using Bacillus sp. strain 3C in contaminated sites. Pestic Biochem. Phys. 2022, 181, 104996. [Google Scholar] [CrossRef]
- Gangola, S.; Joshi, S.; Kumar, S.; Sharma, B.; Sharma, A. Differential proteomic analysis under pesticides stress and normal conditions in Bacillus cereus 2D. PLoS ONE 2021, 16, e0253106. [Google Scholar] [CrossRef] [PubMed]
- Kaur, P.; Sharma, A.; Parihar, L. In vitro study of mycoremediation of cypermethrin-contaminated soils in different regions of Punjab. Ann. Microbiol. 2015, 65, 1949–1959. [Google Scholar] [CrossRef]
- Kitsiou, V.; Filippidis, N.; Mantzavinos, D.; Poulios, I. Heterogeneous and homogeneous photocatalytic degradation of the insecticide imidacloprid in aqueous solutions. Appl. Catal. 2009, 86, 27–35. [Google Scholar] [CrossRef]
- Anhalt, J.C.; Moorman, T.B.; Koskinen, W.C. Biodegradation of imidacloprid by an isolated soil microorganism. J. Environ. Sci. Health B 2007, 42, 509–514. [Google Scholar] [CrossRef]
- Akoijam, R.; Singh, B. Biodegradation of imidacloprid in sandy loam soil by Bacillus aerophilus. Int. J. Environ. Anal Chem. 2015, 95, 730–743. [Google Scholar] [CrossRef]
- Pang, S.; Lin, Z.; Zhang, Y.; Zhang, W.; Alansary, N.; Mishra, S.; Bhatt, P.; Chen, S. Insights into the toxicity and degradation mechanisms of imidacloprid via physicochemical and microbial approaches. Toxics 2020, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Tripathi, P.; Mohan, D.; Singh, R.S. Imidacloprid biodegradation using novel bacteria Tepidibacillus decaturensis strain ST1 in batch and in situ microcosm study. Environ. Sci. Pollut. Res. 2023, 30, 61562–61572. [Google Scholar] [CrossRef]
- Sabourmoghaddam, N.; Zakaria, M.P.; Omar, D. Evidence for the microbial degradation of imidacloprid in soils of Cameron Highlands. J. Saudi. Soc. Agric. Sci. 2015, 14, 182–188. [Google Scholar] [CrossRef]
- Joshi, P.A.; Mhatre, K.J. Microbial efficiency to degrade Carbol fuchsin and Malachite green dyes. Adv. Appl. Sci. Res. 2015, 6, 85–88. [Google Scholar]
- Etezad, S.; Sadeghi-Kiakhani, M. Decolorization of malachite green dye solution by bacterial biodegradation. Prog. Color. Color. Coat. 2021, 14, 79–87. [Google Scholar]
- Asad, S.; Amoozegar, M.; Pourbabaee, A.A.; Sarbolouki, M.; Dastgheib, S. Decolorization of textile azo dyes by newly isolated halophilic and halotolerant bacteria. Bioresour. Technol. 2007, 98, 2082–2088. [Google Scholar] [CrossRef] [PubMed]
- Abo-State, M.; Saleh, Y.; Hazaa, H. Decolorization of Congo Red dye by bacterial isolates. J. Ecol. Health Environ. 2017, 5, 41–48. [Google Scholar]
- Saied, E.; Fouda, A.; Alemam, A.M.; Sultan, M.H.; Barghoth, M.G.; Radwan, A.A.; Desouky, S.G.; Azab, I.H.E.; Nahhas, N.E.; Hassan, S.E.-D. Evaluate the toxicity of pyrethroid insecticide cypermethrin before and after biodegradation by Lysinibacillus cresolivuorans strain HIS7. Plants 2021, 10, 1903. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; Bankole, P.O.; Sadasivam, S.K. Biodecolorization and degradation of textile azo dyes using Lysinibacillus sphaericus MTCC 9523. Front. Environ. Sci. 2022, 10, 990855. [Google Scholar] [CrossRef]
- Gomaa, O.M.; Abd El Kareem, H.; Selim, N. Nitrate modulation of Bacillus sp. biofilm components: A proposed model for sustainable bioremediation. Biotechnol. Lett. 2021, 43, 2185–2197. [Google Scholar] [CrossRef]
- Makino, Y.; Oshita, S.; Murayama, Y.; Mori, M.; Kawagoe, Y.; Sakai, K. Nondestructive analysis of chlorpyrifos on apple skin using UV reflectance. Trans. ASABE 2009, 52, 1955–1960. [Google Scholar] [CrossRef]
- Sinha, S.; Chattopadhyay, P.; Pan, I.; Chatterjee, S.; Chanda, P.; Bandyopadhyay, D.; Das, K.; Sen, S.K. Microbial transformation of xenobiotics for environmental bioremediation. Afr. J. Biotechnol. 2009, 8, 66090. [Google Scholar]
- Armenta, S.; Quintás, G.; Garrigues, S.; de la Guardia, M. A validated and fast procedure for FTIR determination of cypermethrin and chlorpyrifos. Talanta 2005, 67, 634–639. [Google Scholar] [CrossRef]
- Tallur, P.N.; Megadi, V.B.; Ninnekar, H.Z. Biodegradation of cypermethrin by Micrococcus sp. strain CPN 1. Biodegradation 2008, 19, 77–82. [Google Scholar] [CrossRef]
- Lin, Q.; Chen, S.; Hu, M.; Haq, M.; Yang, L.; Li, H. Biodegradation of cypermethrin by a newly isolated actinomycetes HU-S-01 from wastewater sludge. Int. J. Env. Sci. Technol. 2011, 8, 45–56. [Google Scholar] [CrossRef]
- Negi, G.; Srivastava, A.; Sharma, A. In situ biodegradation of endosulfan, imidacloprid, and carbendazim using indigenous bacterial cultures of agriculture fields of Uttarakhand, India. Int. J. Bioeng. Life Sci. 2014, 8, 973–981. [Google Scholar]
- Telke, A.A.; Joshi, S.M.; Jadhav, S.U.; Tamboli, D.P.; Govindwar, S.P. Decolorization and detoxification of Congo red and textile industry effluent by an isolated bacterium Pseudomonas sp. SU-EBT. Biodegradation 2010, 21, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.; Mosharaf, M.K.; Marcus, P.K. Novel bacterial biofilm consortia that degrade and detoxify the carcinogenic diazo dye Congo red. Arch. Microbiol. 2021, 203, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, M.; Kumarasamy, S.; Ranganathan, M.; Kandasamy, S.; Kandasamy, G.; Gnanavel, K. Enzyme and metabolites attained in degradation of chemical pesticides β Cypermethrin by Bacillus cereus. Mater. Today Proc. 2020, 33, 3640–3645. [Google Scholar] [CrossRef]
- Tripathi, A.; Srivastava, S. Biodegradation of orange G by a novel isolated bacterial strain Bacillus megaterium ITBHU01 using response surface methodology. Afr. J. Biotechnol. 2012, 11, 1768–1781. [Google Scholar] [CrossRef]
- Phugare, S.S.; Kalyani, D.C.; Patil, A.V.; Jadhav, J.P. Textile dye degradation by bacterial consortium and subsequent toxicological analysis of dye and dye metabolites using cytotoxicity, genotoxicity and oxidative stress studies. J. Hazards Mater. 2011, 186, 713–723. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liaqat, I.; Khalid, A.; Rubab, S.; Rashid, F.; Latif, A.A.; Naseem, S.; Bibi, A.; Khan, B.N.; Ansar, W.; Javed, A.; et al. In Vitro Biofilm-Mediated Biodegradation of Pesticides and Dye-Contaminated Effluents Using Bacterial Biofilms. Microorganisms 2023, 11, 2163. https://doi.org/10.3390/microorganisms11092163
Liaqat I, Khalid A, Rubab S, Rashid F, Latif AA, Naseem S, Bibi A, Khan BN, Ansar W, Javed A, et al. In Vitro Biofilm-Mediated Biodegradation of Pesticides and Dye-Contaminated Effluents Using Bacterial Biofilms. Microorganisms. 2023; 11(9):2163. https://doi.org/10.3390/microorganisms11092163
Chicago/Turabian StyleLiaqat, Iram, Awais Khalid, Saima Rubab, Farzana Rashid, Asma Abdul Latif, Sajida Naseem, Asia Bibi, Bushra Nisar Khan, Waiza Ansar, Arshad Javed, and et al. 2023. "In Vitro Biofilm-Mediated Biodegradation of Pesticides and Dye-Contaminated Effluents Using Bacterial Biofilms" Microorganisms 11, no. 9: 2163. https://doi.org/10.3390/microorganisms11092163
APA StyleLiaqat, I., Khalid, A., Rubab, S., Rashid, F., Latif, A. A., Naseem, S., Bibi, A., Khan, B. N., Ansar, W., Javed, A., Afzaal, M., Summer, M., Majid, S., Ali, S., & Aftab, M. N. (2023). In Vitro Biofilm-Mediated Biodegradation of Pesticides and Dye-Contaminated Effluents Using Bacterial Biofilms. Microorganisms, 11(9), 2163. https://doi.org/10.3390/microorganisms11092163