
Multi-Locus Phylogeny and Morphology Reveal Two New Species of Hypoxylon (Hypoxylaceae, Xylariales) from Motuo, China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological Studies
2.2. DNA Extraction and Sequencing
2.3. Phylogenetic Analysis

{kind=link}
{kind=link}
{kind=link}
| Species Name | Specimen No. | Locality | GenBank Accession No. | References | |||
|---|---|---|---|---|---|---|---|
| ITS | LSU | RPB2 | β-tubulin | ||||
| Annulohypoxylon annulatum | CBS 140775 | USA | KU604559 | KY610418 | KY624263 | KX376353 | [33,46,47] |
| A. truncatum | CBS 140778 | USA | KX376329 | KY610419 | KY624277 | KX376352 | [33,47] |
| Biscogniauxia nummularia | MUCL 51395 | France | KY610382 | KY610427 | KY624236 | KX271241 | [33] |
| Hypomontagnella barbarensis | STMA 14081 | Argentina | MK131720 | MK131718 | MK135891 | MK135893 | [34] |
| Hy. monticulosa | MUCL 54604 | Guiana | KY610404 | KY610487 | KY624305 | KX271273 | [33] |
| Hy. submonticulosa | CBS 115280 | France | KC968923 | KY610457 | KY624226 | KC977267 | [24,33] |
| Hypoxylon addis | MUCL 52797 | Ethiopia | KC968931 | - | - | KC977287 | [24] |
| H. anthochroum | YMJ 9 | Mexico | JN660819 | - | - | AY951703 | [24] |
| H. aveirense | CMG 29 | Portugal | MN053021 | - | - | MN066636 | [48] |
| H. baihualingense | FCATAS 477 | China | MG490190 | - | - | MH790276 | [37] |
| H. baruense | UCH 9545 | Panama | MN056428 | - | - | MK908142 | [49] |
| H. begae | YMJ 215 | USA | JN660820 | - | - | AY951704 | [32] |
| H. bellicolor | UCH 9543 | Panama | MN056425 | - | - | MK908139 | [49] |
| H. brevisporum | YMJ 36 | Puerto Rico | JN660821 | - | - | AY951705 | [32] |
| H. carneum | MUCL 54177 | France | KY610400 | KY610480 | KY624297 | KX271270 | [33] |
| H. cercidicola | CBS 119009 | France | KC968908 | KY610444 | KY624254 | KX271270 | [24,33] |
| H. chrysalidosporum | FCATAS 2710 | China | OL467294 | OL615106 | OL584222 | OL584229 | [35] |
| H. crocopeplum | CBS 119004 | France | KC968907 | KY610445 | KY624255 | KC977268 | [33] |
| H. cyclobalanopsidis | FCATAS 2714 | China | OL467298 | OL615108 | OL584225 | OL584232 | [35] |
| H. damuense | FCATAS 4207 | China | ON075427 | ON075433 | ON093251 | ON093245 | [19] |
| H. dieckmannii | YMJ 89041203 | China | JN979413 | - | - | AY951713 | [32] |
| H. diperithecium | FCATAS 4226 | China | ON178671 | ON350864 | ON365561 | ON365565 | This study |
| H. diperithecium | FCATAS 4323 | China | ON178672 | ON350865 | ON365562 | ON365566 | This study |
| H. duranii | YMJ 85 | China | JN979414 | - | - | AY951714 | [32] |
| H. erythrostroma | YMJ 90080602 | China | JN979416 | - | - | AY951716 | [32] |
| H. eurasiaticum | MUCL 57720 | Iran | MW367851 | - | MW373852 | MW373861 | [50] |
| H. fendleri | DSM 107927 | USA | MK287533 | MK287545 | MK287558 | MK287571 | [51] |
| H. ferrugineum | CBS 141259 | Austria | KX090079 | - | - | KX090080 | [52] |
| H. fragiforme | MUCL 51264 | Germany | KM186294 | KM186295 | KM186296 | KM186293 | [51] |
| H. fraxinophilum | MUCL 54176 | France | KC968938 | - | - | KC977301 | [24] |
| H. fulvosulphureum | MFLUCC 13-0589 | Thailand | KP401576 | - | - | KP401584 | [53] |
| H. fuscum | CBS 113049 | France | KY610401 | KY610482 | KY624299 | KX271271 | [33] |
| H. gibriacense | MUCL 52698 | Germany | KC968930 | - | - | - | [24] |
| H. greiderae | BRIP 72533 | USA | NR 182619 | OP598062 | - | - | [54] |
| H. griseobrunneum | CBS 331.73 | India | KY610402 | MH872399 | KY624300 | KC977303 | [24,33,55] |
| H. guilanense | MUCL 57726 | Iran | MT214997 | MT214992 | MT212235 | MT212239 | [56] |
| H. haematostroma | MUCL 53301 | Martinique | KC968911 | KY610484 | KY624301 | KC977291 | [34] |
| H. hainanense | FCATAS 2712 | China | OL467296 | OL616132 | OL584224 | OL584231 | [35] |
| H. hinnuleum | MUCL 3621 | USA | MK287537 | MK287549 | MK287562 | MK287575 | [51] |
| H. howeanum | MUCL 47599 | Germany | AM749928 | KY610448 | KY624258 | KC977277 | [24,33,57] |
| H. hypomiltum | MUCL 51845 | Guadeloupe | KY610403 | KY610449 | KY624302 | KX271249 | [33] |
| H. invadens | MUCL 51475 | France | MT809133 | MT809132 | MT813037 | MT813038 | [58] |
| H. investiens | CBS 118183 | Malaysia | KC968925 | KY610450 | KY624259 | KC977270 | [24,33] |
| H. isabellinum | STMA 10247 | Martinique | KC968935 | - | - | KC977295 | [24] |
| H. jaklitschii | JF13037 | Sri Lanka | KM610290 | - | - | KM610304 | [24] |
| H. jecorinum | YMJ 39 | Mexico | JN979429 | - | - | AY951731 | [32] |
| H. jianfengense | FACATAS845 | China | MW984546 | MZ029707 | MZ047260 | MZ047264 | [36] |
| H. larissae | FACATAS844 | China | MW984548 | MZ029706 | MZ047258 | MZ047262 | [36] |
| H. laschii | MUCL 52796 | Germany | JX658525 | - | - | - | [59] |
| H. lateripigmentum | MUCL 53304 | Martinique | KC968933 | KY610486 | KY624304 | KC977290 | [24,33] |
| H. lenormandii | CBS 135869 | Cameroon | KY610390 | KY610453 | KY624262 | KM610295 | [33,60] |
| H. liviae | CBS 115282 | Norway | NR155154 | - | - | KC977265 | [24] |
| H. lividicolor | YMJ 70 | China | JN979432 | - | - | AY951734 | [32] |
| H. lividipigmentum | YMJ 233 | Mexico | JN979433 | - | - | AY951735 | [32] |
| H. macrosporum | YMJ 47 | Canada | JN979434 | - | - | AY951736 | [32] |
| H. medogense | FCATAS 4061 | China | ON075425 | ON075431 | ON093249 | ON093243 | [19] |
| H. munkii | YMJ 90080403 | China | JN979436 | - | - | AY951738 | [32] |
| H. musceum | MUCL 53765 | Guadeloupe | KC968926 | KY610488 | KY624306 | KC977280 | [24,33] |
| H. notatum | YMJ 250 | USA | JQ009305 | - | - | AY951739 | [32] |
| H. olivaceopigmentum | DSM 10792 | USA | MK287530 | MK287542 | MK287555 | MK287568 | [51] |
| H. perforatum | CBS 115281 | France | KY610391 | KY610455 | KY624224 | KX271250 | [33] |
| H. petriniae | CBS 114746 | France | NR155185 | KY610491 | KY624279 | KX271274 | [33] |
| H. pilgerianum | STMA 13455 | Martinique | KY610412 | - | KY624308 | KY624315 | [33] |
| H. porphyreum | CBS 119022 | France | KC968921 | KY610456 | KY624225 | KC977264 | [24,33] |
| H. pseudofendleri | MFLUCC 11-0639 | Thailand | KU940156 | KU863144 | - | - | [61] |
| H. pseudofuscum | 18264 | Germany | MW367857 | MW367848 | MW373858 | MW373867 | [50] |
| H. pulicicidum | CBS 122622 | Martinique | JX183075 | KY610492 | KY624280 | JX183072 | [33,62] |
| H. rickii | MUCL 53309 | Martinique | KC968932 | KY610416 | KY624281 | KC977288 | [33] |
| H. rubiginosum | MUCL 52887 | Germany | KC477232 | KY610469 | KY624266 | KY624311 | [33,63] |
| H. rutilum | YMJ 181 | France | - | - | - | AY951752 | [32] |
| H. samuelsii | MUCL 51843 | Guadeloupe | KC968916 | KY610466 | KY624269 | KC977286 | [24,33] |
| H. shearii | YMJ 29 | Mexico | EF026142 | - | - | AY951753 | [32] |
| H. spegazzinianum | STMA 14082 | Argentina | KU604573 | - | - | KU604582 | [64] |
| H. sporistriatatunicum | UCH 9542 | Panama | MN056426 | - | - | MK908140 | [49] |
| H. subgilvum | YMJ 88113007 | China | JQ009315 | - | - | AY951755 | [32] |
| H. sublenormandii | JF 13026 | Sri Lanka | KM610291 | - | - | KM610303 | [60] |
| H. teeravasati | PUFD4 | India | KY863509 | MF385274 | MG986895 | MG986894 | [65] |
| H. texense | DSM 107933 | USA | MK287536 | MK287548 | MK287561 | MK287574 | [51] |
| H. tibeticum | FCATAS4022 | China | OR654146 | OR654303 | ON254302 | ON230084 | This study |
| H. tibeticum | FCATAS4371 | China | OR654263 | OR654304 | QQ303928 | QQ303964 | This study |
| H. tibeticum | FCATAS4212 | China | OR654264 | OR654305 | ON254308 | ON254275 | This study |
| H. tibeticum | FCATAS4373 | China | OR654265 | OR654306 | QQ303933 | QQ303965 | This study |
| H. ticinense | CBS 115271 | France | JQ009317 | KY610471 | KY624272 | AY951757 | [32,33] |
| H. trugodes | MUCL 54794 | Sri Lanka | KF234422 | NG066380 | KY624282 | KF300548 | [24,33] |
| H. ulmophilum | YMJ 350 | Russia | JQ009320 | - | - | AY951760 | [32] |
| H. vinosopulvinatum | YMJ 90080707 | China | JQ009321 | - | - | AY951761 | [32] |
| H. vogesiacum | CBS 115273 | France | KC968920 | KY610417 | KY624283 | KX271275 | [33] |
| H. wujiangense | GMBC0213 | China | MT568854 | MT568853 | MT585802 | MT572481 | [66] |
| H. wuzhishanense | FCATAS 2708 | China | OL467292 | OL615104 | OL584220 | OL584227 | [35] |
| H. zangii | FCATAS 6092 | China | OQ316425 | OQ348528 | OQ303910 | OQ303948 | [19] |
| Jackrogersella cohaerens | CBS 119126 | Germany | KY610396 | KY610497 | KY624270 | KY624314 | [33] |
| J. multiformis | CBS 119016 | Germany | KC477234 | KY610473 | KY624290 | KX271262 | [24,33] |
| Parahypoxylon papillatum | ATCC 58729 | USA | NR155153 | KY610454 | KY624223 | KC977258 | [24,33] |
| Pyrenopolyporus hunteri | MUCL 52673 | Ivory Coast | KY610421 | KY610472 | KY624309 | KU159530 | [33,47] |
| Py.laminosus | MUCL 53305 | Martinique | KC968934 | KY610485 | KY624303 | KC977292 | [24,33] |
| Py. nicaraguensis | CBS 117739 | Burkina Faso | AM749922 | KY610489 | KY624307 | KC977272 | [24,33,57] |
| Rhopalostroma angolense | CBS 126414 | Ivory Coast | KY610420 | KY610459 | KY624228 | KX271277 | [33] |
| Thamnomyces dendroidea | CBS 123578 | FrenchGuiana | FN428831 | KY610467 | KY624232 | KY624313 | [33,67] |
| Xylaria hypoxylon | CBS 122620 | Sweden | KY610407 | KY610495 | KY624231 | KX271279 | [33] |
3. Results
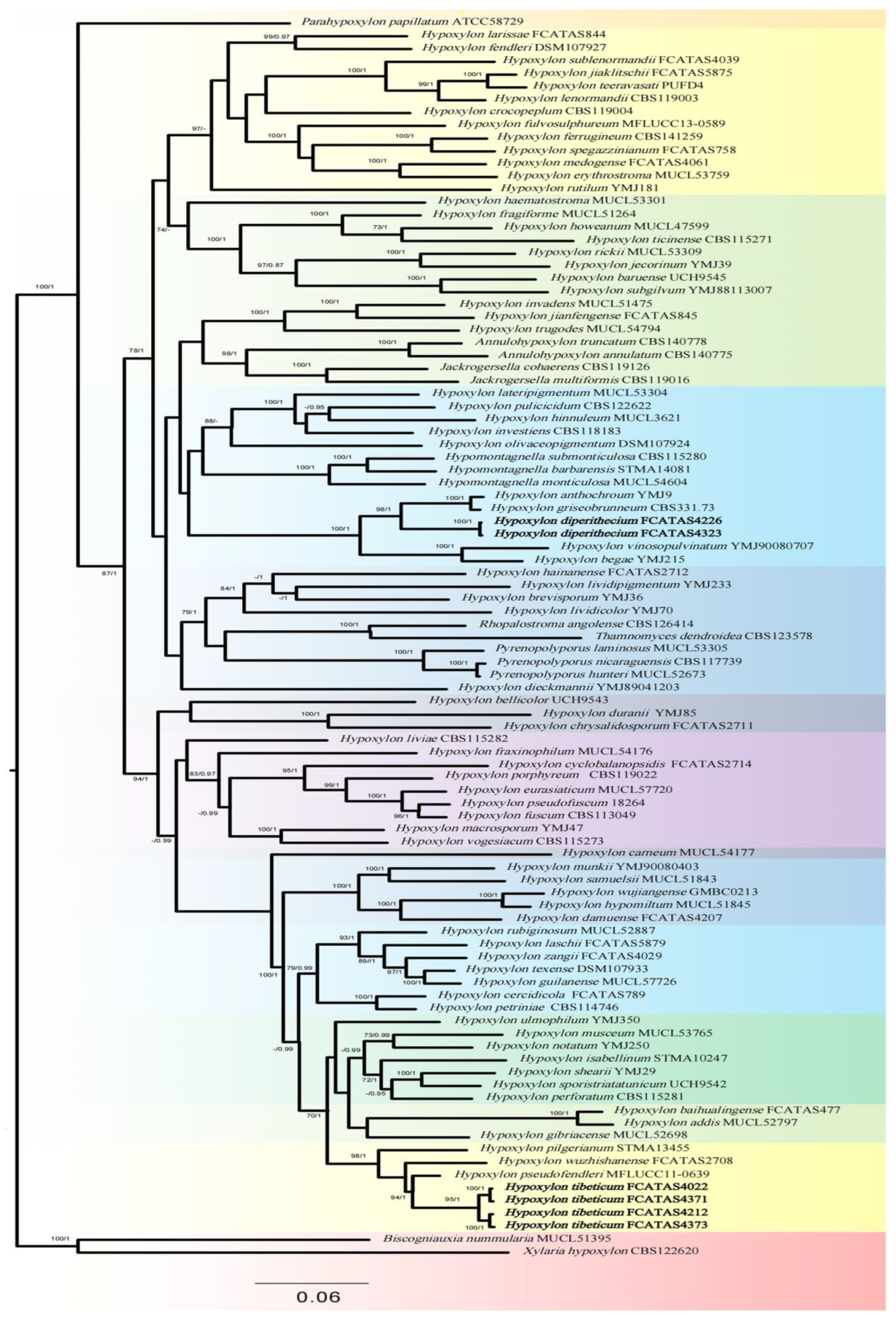
3.1. Phylogenetic Analysis
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fu, D.F.; Song, Q.G.; Li, D.J. Analysis of forest vegetation biomass and carbon storage in Motuo county of Tibet. Hunan For. Sci. Technol. 2015, 42, 67–72. [Google Scholar] [CrossRef]
- Peng, B.Z. Some problems of vertical zonation in MT. Namjagbarwa area. Acta Geogr. Sin. 1986, 41, 51–58. [Google Scholar]
- Feng, M.; Zhu, R.J.; Zhao, G.F. Utilization of wild plant resources and development suggestions of agricultural industry in Motuo tropical area of Tibet. Chin. J. Trop. Agric. 2022, 42, 38–41. [Google Scholar] [CrossRef]
- Chen, P.; Li, B. Analysis of water vapour transport characteristics and implications in Southeast Tibetan Plateau. South China Agric. 2018, 12, 124–125. [Google Scholar] [CrossRef]
- Wang, J.; De, Q.; Dan, Z.; Wang, Y.; De, J.; Chen, G.Y. Characteristic and causal analysis of concentrated and frequently occurring intense rainfall in Tibet in 2012–2018. Meteorol. Sci. Technol. 2021, 49, 211–217. [Google Scholar] [CrossRef]
- Wu, Z.Y. Flora of Tibet; Science Press: Beijing, China, 1983; Volume 1, 791p. [Google Scholar]
- Wu, Z.Y. Flora of Tibet; Science Press: Beijing, China, 1985; Volume 2, 956p. [Google Scholar]
- Wu, Z.Y. Flora of Tibet; Science Press: Beijing, China, 1986; Volume 3, 1046p. [Google Scholar]
- Wu, Z.Y. Flora of Tibet; Science Press: Beijing, China, 1985; Volume 4, 1021p. [Google Scholar]
- Wu, Z.Y. Flora of Tibet; Science Press: Beijing, China, 1987; Volume 5, 955p. [Google Scholar]
- Wang, Y.C.; Zang, M.; Ma, Q.M.; Kong, H.Z.; Mao, X.L.; Qi, Z.T.; Sun, Z.M.; Shen, R.X.; Ying, J.Z.; Li, M.X.; et al. Fungi of Tibet; Science Press: Beijing, China, 1983; 224p. [Google Scholar]
- Mao, X.L.; Jiang, C.P.; Ouzhu, C.W. Economic Fungi of Tibet; Science Press: Beijing, China, 1993; 651p. [Google Scholar]
- Ge, Z.W.; Yang, Z.L.; Vellinga, E.C. The genus Macrolepiota (Agaricaceae, Basidiomycota) in China. Fungal Divers. 2010, 45, 81–98. [Google Scholar] [CrossRef]
- Li, G.J.; Zhao, D.; Li, S.F.; Wen, H.A.; Liu, X.Z. Russula chiui and R. pseudopectinatoides, two new species from southwestern China supported by morphological and molecular evidence. Mycol. Prog. 2015, 14, 1–14. [Google Scholar] [CrossRef]
- Wu, F.; Chen, J.J.; Ji, X.H.; Vlasák, J.; Dai, Y.C. Phylogeny and diversity of the morphologically similar polypore genera Rigidoporus, Physisporinus, Oxyporus, and Leucophellinus. Mycologia 2017, 109, 749–765. [Google Scholar] [CrossRef]
- Song, J.; Sun, Y.F.; Ji, X.; Dai, Y.C.; Cui, B.K. Phylogeny and taxonomy of Laetiporus (Basidiomycota, Polyporales) with descriptions of two new species from western China. MycoKeys 2018, 37, 57–71. [Google Scholar] [CrossRef]
- Wang, P.M.; Yang, Z.L. Two new taxa of the Auriscalpium vulgare species complex with substrate preferences. Mycol. Prog. 2019, 18, 641–652. [Google Scholar] [CrossRef]
- Liu, S.; Chen, Y.Y.; Sun, Y.F.; He, X.L.; Song, C.G.; Si, J.; Liu, D.M.; Gates, G.; Cui, B.K. Systematic classification and phylogenetic relationships of the brown-rot fungi within the Polyporales. Fungal Divers. 2023, 118, 1–94. [Google Scholar] [CrossRef]
- Song, Z.K.; Zhu, A.H.; Liu, Z.D.; Qu, Z.; Li, Y.; Ma, H.X. Three new species of Hypoxylon (Xylariales, Ascomycota) on a multigene phylogeny from Medog in Southwest China. J. Fungi 2022, 8, 500. [Google Scholar] [CrossRef] [PubMed]
- Zhu, A.H.; Gao, Y.; Tang, K.; Song, Z.K.; Yang, Z.E.; Ma, H.X. Newly recorded species of Hypoxylaceae (Ascomycota) from Medog in China. Chin. J. Trop. Crops 2022, 43, 2268–2274. [Google Scholar] [CrossRef]
- Ma, H.X.; Yang, Z.E.; Song, Z.K.; Qu, Z.; Li, Y.; Zhu, A.H. Taxonomic and phylogenetic contributions to Diatrypaceae from southeastern Tibet in China. Front. Microbiol. 2023, 14, 1073548. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.H. A Monograph of the World Species of Hypoxylon; University Georgia Press: Athens, Greece, 1961; 158p. [Google Scholar]
- Ju, Y.M.; Rogers, J.D. A Revision of the Genus Hypoxylon; American Phytopathological Society Press: St. Paul, MN, USA, 1996; 365p. [Google Scholar]
- Ma, H.X.; Qiu, J.Z.; Xu, B.; Li, Y. Two Hypoxylon species from Yunnan Province based on morphological and molecular characters. Phytotaxa 2018, 376, 27–36. [Google Scholar] [CrossRef]
- Kuhnert, E.; Fournier, J.; Peršoh, D.; Luangsa-ard, J.J.; Stadler, M. New Hypoxylon species from Martinique and new evidence on the molecular phylogeny of Hypoxylon based on ITS rDNA and b-tubulin data. Fungal Divers. 2014, 64, 181–203. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Rajeshkumar, K.C.; Hawksworth, D.L.; Madrid, H.; Kirk, P.M.; Braun, U.; Singh, R.V.; Crous, P.W.; Kukwa, M.; et al. Notes for genera: Ascomycota. Fungal Divers. 2017, 86, 1–594. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycets. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Whalley, A.J.S. The Xylariaceae: Some ecological considerations. Sydowia 1985, 38, 369–382. [Google Scholar]
- Rogers, J.D. Thoughts and musings on tropical Xylariaceae. Mycol. Res. 2000, 104, 1412–1420. [Google Scholar] [CrossRef]
- U’Ren, J.M.; Miadlikowska, J.; Zimmerman, N.B.; Lutzoni, F.; Stajich, J.E.; Arnold, A.E. Contributions of North American endophytes to the phylogeny, ecology, and taxonomy of Xylariaceae (Sordariomycetes, Ascomycota). Mol. Phylogenet. Evol. 2016, 98, 210–232. [Google Scholar] [CrossRef] [PubMed]
- Daranagama, D.A.; Hyde, K.D.; Sir, E.B.; Thambugala, K.M.; Tian, Q.; Samarakoon, M.C.; McKenzie, E.H.C.; Jayasiri, S.C.; Tibpromma, S.; Bhat, J.D.; et al. Towards a natural classification and backbone tree for Graphostromataceae, Hypoxylaceae, Lopadostomataceae and Xylariaceae. Fungal Divers. 2018, 88, 1–165. [Google Scholar] [CrossRef]
- Helaly, S.E.; Thongbai, B.; Stadler, M. Diversity of biologically active secondary metabolites from endophytic and saprotrophic fungi of the ascomycete order Xylariales. Nat. Prod. Rep. 2018, 35, 992–1014. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, H.; Ju, Y.M.; Rogers, J.D. Molecular phylogeny of Hypoxylon and closely related genera. Mycologia 2005, 97, 844–865. [Google Scholar] [CrossRef] [PubMed]
- Wendt, L.; Sir, E.B.; Kuhnert, E.; Heitkämper, S.; Lambert, C.; Hladki, A.I.; Romero, A.I.; Luangsaard, J.J.; Srikitikulchai, P.; Per, D.; et al. Resurrection and emendation of the Hypoxylaceae, recognised from a multigene phylogeny of the Xylariales. Mycol. Prog. 2018, 17, 115–154. [Google Scholar] [CrossRef]
- Lambert, C.; Wendt, L.; Hladki, A.I.; Stadler, M.; Sir, E.B. Hypomontagnella (Hypoxylaceae): A new genus segregated from Hypoxylon by a polyphasic taxonomic approach. Mycol. Prog. 2019, 18, 187–201. [Google Scholar] [CrossRef]
- Ma, H.X.; Song, Z.K.; Pan, X.Y.; Li, Y.; Yang, Z.E.; Qu, Z. Multi-gene phylogeny and taxonomy of Hypoxylon (Hypoxylaceae, Ascomycota) from China. Diversity 2022, 14, 37. [Google Scholar] [CrossRef]
- Song, Z.K.; Pan, X.Y.; Li, C.T.; Ma, H.X.; Li, Y. Two new species of Hypoxylon (Hypoxylaceae) from China based on morphological and DNA sequence data analyses. Phytotaxa 2022, 538, 213–224. [Google Scholar] [CrossRef]
- Rayner, R.W. A Mycological Colour Chart; Commonwealth Mycological Institute: London, UK, 1970. [Google Scholar]
- White, T.J.; Bruns, T.D.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics-science direct. PCR Protoc. 1990, 18, 315–322. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- O‘donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. Bioedit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Franois, J.; Higgins, D.G. The CLUSTAL X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hőhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Granmo, A.; Hammelev, D.; Knudsen, H.; Læssøe, T.; Whalley, A.J.S. The genera Biscogniauxia and Hypoxylon (Sphaeriales) in the Nordic countries. Opera Bot. 1989, 100, 59–84. [Google Scholar]
- Kuhnert, E.; Sir, E.B.; Lambert, C.; Hyde, K.D.; Hladki, A.I.; Romero, A.I.; Rohde, M.; Stadler, M. Phylogenetic and chemotaxonomic resolution of the genus Annulohypoxylon (Xylariaceae) including four new species. Fungal Divers. 2017, 85, 1–43. [Google Scholar] [CrossRef]
- Vicente, T.F.L.; Goncalves, M.F.M.; Brandão, C.; Fidalgo, C.; Alves, A. Diversity of fungi associated with macroalgae from an estuarine environment and description of Cladosporium rubrum sp. nov. and Hypoxylon aveirense sp. nov. Int. J. Syst. Evol. Micr. 2021, 71, 004630. [Google Scholar] [CrossRef]
- Cedeño-Sanchez, M.; Wendt, L.; Stadler, M.; Mejía, L.C. Three new species of Hypoxylon and new records of Xylariales from Panama. Mycosphere 2020, 11, 1457–1476. [Google Scholar] [CrossRef]
- Sir, E.B.; Becker, K.; Lambert, C.; Bills, G.F.; Kuhnert, E. Observations on Texas hypoxylons, including two new Hypoxylon species and widespread environmental isolates of the H. croceum complex identified by a polyphasic approach. Mycologia 2019, 11, 832–856. [Google Scholar] [CrossRef]
- Daranagama, D.A.; Camporesi, E.; Tian, Q.; Liu, X.Z.; Chamyuang, S.; Stadler, M.; Hyde, K.D. Anthostomella is polyphyletic comprising several genera in Xylariaceae. Fungal Divers. 2015, 73, 203–238. [Google Scholar] [CrossRef]
- Friebes, G.; Wendelin, I. Studies on Hypoxylon ferrugineum (Xylariaceae), a rarely reported species collected in the urban area of Graz (Austria). Ascomycete.org 2016, 8, 83–90. [Google Scholar]
- Sir, E.B.; Kuhnert, E.; Surup, F.; Hyde, K.D.; Stadler, M. Discovery of new mitorubrin derivatives from Hypoxylon fulvosulphureum sp. nov. (Ascomycota, Xylariales). Mycol. Prog. 2015, 14, 28. [Google Scholar] [CrossRef]
- Tan, Y.P.; Shivas, R.G. Index of Australian Fungi No. 1; Cardwell: Queensland, Australia, 2022. [Google Scholar]
- Vu, D.; Groenewald, M.; Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Pourmoghaddam, M.J.; Lambert, C.; Surup, F.; Khodaparast, S.A.; Krisai-Greilhuber, I.; Voglmayr, H.; Stadler, M. Discovery of a new species of the Hypoxylon rubiginosum complex from Iran and antagonistic activities of Hypoxylon spp. against the Ash Dieback pathogen, Hymenoscyphus fraxineus, in dual culture. MycoKeys 2020, 66, 105–133. [Google Scholar] [CrossRef] [PubMed]
- Bitzer, J.; Læssøe, T.; Fournier, J.; Kummer, V.; Decock, C.; Tichy, H.V.; Piepenbring, M.; Peršoh, D.; Stadler, M. Affinities of Phylacia and the daldinoid Xylariaceae, inferred from chemotypes of cultures and ribosomal DNA sequences. Mycol. Res. 2008, 112, 251–270. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Lambert, C.; Wieschhaus, J.; Stadler, M. Phylogenetic assignment of the fungicolous Hypoxylon invadens (Ascomycota, Xylariales) and investigation of its secondary metabolites. Microorganisms 2020, 8, 1397. [Google Scholar] [CrossRef] [PubMed]
- Stadler, M.; Læssøe, T.; Fournier, J.; Decock, C.; Schmieschek, B.; Tichy, H.V.; Peršoh, D. A polyphasic taxonomy of Daldinia (Xylariaceae). Stud. Mycol. 2014, 77, 1–143. [Google Scholar] [CrossRef]
- Kuhnert, E.; Surup, F.; Sir, E.B.; Lambert, C.; Hyde, K.D.; Hladki, A.I.; Romero, A.I.; Stadler, M. Lenormandins A–G, new azaphilones from Hypoxylon lenormandii and Hypoxylon jaklitschii sp. nov., recognised by chemotaxonomic data. Fungal Divers. 2015, 71, 165–184. [Google Scholar] [CrossRef]
- Dai, D.Q.; Phookamsak, R.; Wijayawardene, N.N.; Li, W.J.; Bhat, D.J.; Xu, J.C.; Taylor, J.E.; Hyde, K.D.; Chukeatirote, E. Bambusicolous fungi. Fungal Divers. 2017, 82, 1–105. [Google Scholar] [CrossRef]
- Bills, G.F.; González-Menéndez, V.; Martín, J.; Platas, G.; Fournier, J.; Peršoh, D.; Stadler, M. Hypoxylon pulicicidum sp. nov. (Ascomycota, Xylariales), a pantropical insecticide-producing endophyte. PLoS ONE 2012, 7, e46687. [Google Scholar] [CrossRef] [PubMed]
- Stadler, M.; Kuhnert, E.; Peršoh, D.; Fournier, J. The Xylariaceae as model example for a unified nomenclature following the “One Fungus-One Name” (1F1N) concept. Mycology 2013, 4, 5–21. [Google Scholar]
- Sir, E.B.; Kuhnert, E.; Lambert, C.; Hladki, A.I.; Romero, A.I.; Stadler, M. New species and reports of Hypoxylon from Argentina recognized by a polyphasic approach. Mycol. Prog. 2016, 15, 42. [Google Scholar] [CrossRef]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspé, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef]
- Pi, Y.H.; Zhang, X.; Liu, L.L.; Long, Q.D.; Shen, X.C.; Kang, Y.Q.; Hyde, K.D.; Boonmee, S.; Kang, J.C.; Li, Q.R. Contributions to species of Xylariales in China—4 Hypoxylon wujiangensis sp. nov. Phytotaxa 2020, 455, 21–30. [Google Scholar] [CrossRef]
- Stadler, M.; Fournier, J.; Laessøe, T.; Chlebicki, A.; Lechat, C.; Flessa, F.; Rambold, G.; Peršoh, D. Chemotaxonomic and phylogenetic studies of Thamnomyces (Xylariaceae). Mycoscience 2010, 51, 189–207. [Google Scholar] [CrossRef]
- Fournier, J.; Lechat, C.; Courtecuisse, R. The genus Hypoxylon (Xylariaceae) in Guadeloupe and Martinique (French West Indies). Ascomycete.org 2016, 7, 145–212. [Google Scholar]
- Hennings, P. Fungi Matogrossenses a Dr. R. Pilger collecti 1899. Beibl. Hedwig. 1900, 39, 134–139. [Google Scholar]
- Sanchez-Ballesteros, J.; Gonzalez, V.; Salazar, O.; Acero, J.; Portal, M.A.; Julián, M.; Rubio, V.; Bill, G.F.; Polishook, J.D.; Platas, G.; et al. Phylogenetic study of Hypoxylon and related genera based on ribosomal ITS sequences. Mycologia 2000, 92, 964–977. [Google Scholar] [CrossRef]
- Triebel, D.; Peršoh, D.; Wollweber, H.; Stadler, M. Phylogenetic relationships among Daldinia, Entonaema and Hypoxylon as inferred from ITS nrDNA analyses of Xylariales. Nova Hedwig. 2005, 80, 25–43. [Google Scholar] [CrossRef]
- Cedeño-Sanchez, M.; Charria-Girón, E.; Lambert, C.; Luangsa-ard, J.J.; Decock, C.; Franke, R.; Brönstrup, M.; Stadler, M. Segregation of the genus Parahypoxylon (Hypoxylaceae, Xylariales) from Hypoxylon by a polyphasic taxonomic approach. MycoKeys 2023, 95, 131–162. [Google Scholar] [CrossRef] [PubMed]
- Chi, S.Q.; Xu, J.; Lu, B.S. Three New Chinese Records of Hypoxylon. J. Fungal Res. 2016, 14, 218–221. [Google Scholar] [CrossRef]
- Guo, L. Flora Fungorum Sinicorum. Volume 64. Annulohypoxylon, Hypoxylon, Rosellinia; Science Press: Beijing, China, 2022; 155p. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, A.-H.; Song, Z.-K.; Wang, J.-F.; Guan, H.-W.; Ma, H.-X. Multi-Locus Phylogeny and Morphology Reveal Two New Species of Hypoxylon (Hypoxylaceae, Xylariales) from Motuo, China. Microorganisms 2024, 12, 72. https://doi.org/10.3390/microorganisms12010072
Zhu A-H, Song Z-K, Wang J-F, Guan H-W, Ma H-X. Multi-Locus Phylogeny and Morphology Reveal Two New Species of Hypoxylon (Hypoxylaceae, Xylariales) from Motuo, China. Microorganisms. 2024; 12(1):72. https://doi.org/10.3390/microorganisms12010072
Chicago/Turabian StyleZhu, An-Hong, Zi-Kun Song, Jun-Fang Wang, Hao-Wen Guan, and Hai-Xia Ma. 2024. "Multi-Locus Phylogeny and Morphology Reveal Two New Species of Hypoxylon (Hypoxylaceae, Xylariales) from Motuo, China" Microorganisms 12, no. 1: 72. https://doi.org/10.3390/microorganisms12010072
APA StyleZhu, A. -H., Song, Z. -K., Wang, J. -F., Guan, H. -W., & Ma, H. -X. (2024). Multi-Locus Phylogeny and Morphology Reveal Two New Species of Hypoxylon (Hypoxylaceae, Xylariales) from Motuo, China. Microorganisms, 12(1), 72. https://doi.org/10.3390/microorganisms12010072