

Characterization of Antibiotic-Resistance Antarctic Pseudomonas That Produce Bacteriocin-like Compounds
 ,
,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Origin of the Antarctic Bacteria
2.2. Morphological and Biochemical Characterization of C1-4-7, D2-4-6, and M1-4-11 Antarctic Bacteria
2.3. Antibiotic Susceptibility Profile of Antarctic Bacteria
2.4. Molecular Identification of Antarctic Bacteria
2.5. Genotyping of Antarctic Bacteria by Repetitive Elements PCR Amplification
2.6. Screening for Antimicrobial Activity of the Antarctic Bacteria and Their Cell-Free Culture Supernatants
2.7. Exclusion Membrane Assay
2.8. Sensitivity of the Antimicrobial Agents to Proteinase K Enzyme
2.9. Effect of Temperature and Culture Medium on the Growth and Bacteriocin-like Production of the Antarctic Bacteria
2.10. Statistical Analysis
3. Results
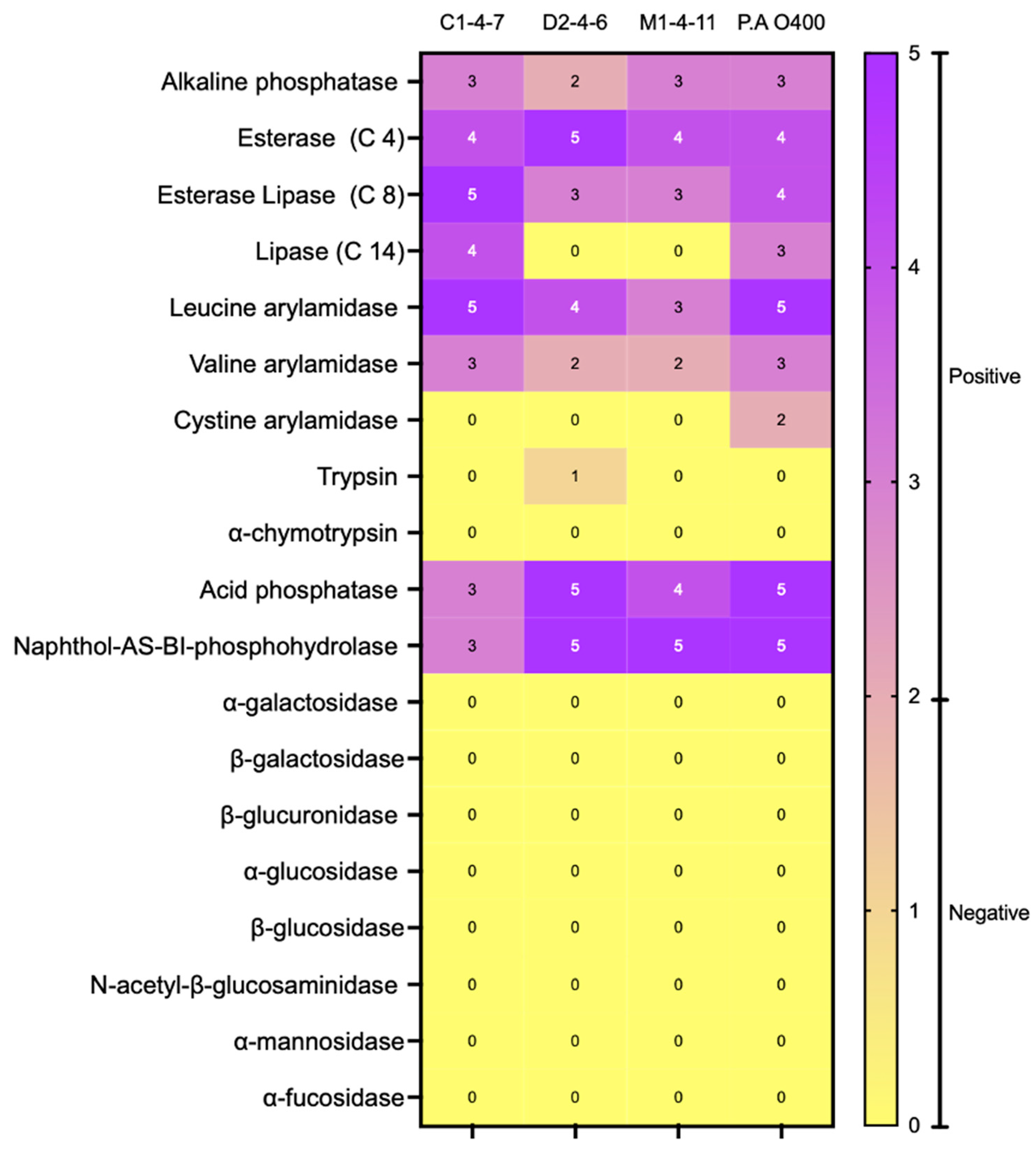
3.1. Morphological and Biochemical Characterization of Antarctic Bacteria
3.2. Molecular Identification of Antarctic Bacteria
3.3. Genotyping of Antarctic Bacteria by Detection of Repetitive Elements Amplified by PCR
3.4. Antibiotic Susceptibility Profile of Antarctic Pseudomonas sp. C1-4-7, D2-4-6, and M1-4-11
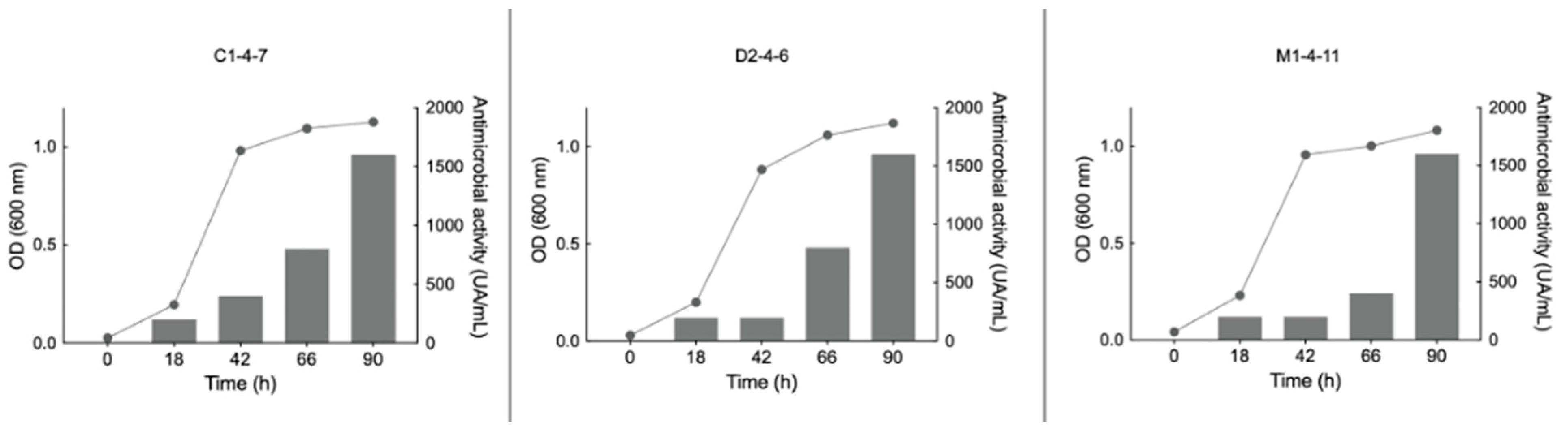
3.5. Effect of Temperature and Culture Medium on the Growth of the Antarctic Bacteria C1-4-7, D2-4-6, and M1-4-11
3.6. Screening for Antimicrobial Activity of the Antarctic Bacteria C1-4-7, D2-4-6, and M1-4-11 and Their Cell-Free Culture Supernatants Concentrated at 80 °C
3.7. Exclusion Membrane Assay
3.8. Sensitivity of the Antimicrobial Agents to Proteinase K Enzyme
3.9. Effect of Temperature and Culture Medium on the Bacteriocin-like Production of the Antarctic Bacteria C1-4-7, D2-4-6, and M1-4-11
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cole, S.T. Who Will Develop New Antibacterial Agents? Phil. Trans. R. Soc. B 2014, 369, 20130430. [Google Scholar] [CrossRef] [PubMed]
- Baricz, A.; Teban, A.; Chiriac, C.M.; Szekeres, E.; Farkas, A.; Nica, M.; Dascălu, A.; Oprișan, C.; Lavin, P.; Coman, C. Investigating the Potential Use of an Antarctic Variant of Janthinobacterium Lividum for Tackling Antimicrobial Resistance in a One Health Approach. Sci. Rep. 2018, 8, 15272. [Google Scholar] [CrossRef] [PubMed]
- Joerger, R. Alternatives to Antibiotics: Bacteriocins, Antimicrobial Peptides and Bacteriophages. Poult. Sci. 2003, 82, 640–647. [Google Scholar] [CrossRef]
- Yang, S.-C.; Lin, C.-H.; Sung, C.T.; Fang, J.-Y. Antibacterial Activities of Bacteriocins: Application in Foods and Pharmaceuticals. Front. Microbiol. 2014, 5, 241. [Google Scholar] [CrossRef]
- Mishra, B.; Reiling, S.; Zarena, D.; Wang, G. Host Defense Antimicrobial Peptides as Antibiotics: Design and Application Strategies. Curr. Opin. Chem. Biol. 2017, 38, 87–96. [Google Scholar] [CrossRef]
- Ahmad, V.; Jamal, Q.M.S.; Siddiqui, M.U.; Shukla, A.K.; Alzohairy, M.A.; Al Karaawi, M.A.; Kamal, M.A. Methods of Screening-Purification and Antimicrobial Potentialities of Bacteriocin in Health Care. Curr. Drug Metab. 2017, 18, 814–830. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuji, T.; Gallo, R.L. Antimicrobial Peptides: Old Molecules with New Ideas. J. Investig. Dermatol. 2012, 132, 887–895. [Google Scholar] [CrossRef]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A Viable Alternative to Antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Microbial Production of Bacteriocins: Latest Research Development and Applications. Biotechnol. Adv. 2018, 36, 2187–2200. [Google Scholar] [CrossRef]
- Silva, T.R.; Duarte, A.W.F.; Passarini, M.R.Z.; Ruiz, A.L.T.G.; Franco, C.H.; Moraes, C.B.; de Melo, I.S.; Rodrigues, R.A.; Fantinatti-Garboggini, F.; Oliveira, V.M. Bacteria from Antarctic Environments: Diversity and Detection of Antimicrobial, Antiproliferative, and Antiparasitic Activities. Polar Biol. 2018, 41, 1505–1519. [Google Scholar] [CrossRef]
- Katz, L.; Baltz, R.H. Natural Product Discovery: Past, Present, and Future. J. Ind. Microbiol. Biotechnol. 2016, 43, 155–176. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Rendon, D.; Passari, A.K.; Ruiz-Villafán, B.; Rodríguez-Sanoja, R.; Sánchez, S.; Demain, A.L. Impact of Novel Microbial Secondary Metabolites on the Pharma Industry. Appl. Microbiol. Biotechnol. 2022, 106, 1855–1878. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Montero, K.; Lamilla, C.; Abanto, M.; Maruyama, F.; Jorquera, M.A.; Santos, A.; Martinez-Urtaza, J.; Barrientos, L. Antarctic Streptomyces Fildesensis So13.3 Strain as a Promising Source for Antimicrobials Discovery. Sci. Rep. 2019, 9, 7488. [Google Scholar] [CrossRef] [PubMed]
- Asencio, G.; Lavin, P.; Alegría, K.; Domínguez, M.; Bello, H.; González-Rocha, G.; González-Aravena, M. Antibacterial Activity of the Antarctic Bacterium Janthinobacterium Sp. SMN 33.6 against Multi-Resistant Gram-Negative Bacteria. Electron. J. Biotechnol. 2014, 17, 1–5. [Google Scholar] [CrossRef]
- Maida, I.; Fondi, M.; Papaleo, M.C.; Perrin, E.; Orlandini, V.; Emiliani, G.; De Pascale, D.; Parrilli, E.; Tutino, M.L.; Michaud, L.; et al. Phenotypic and Genomic Characterization of the Antarctic Bacterium Gillisia Sp. CAL575, a Producer of Antimicrobial Compounds. Extremophiles 2014, 18, 35–49. [Google Scholar] [CrossRef]
- Tedesco, P.; Maida, I.; Palma Esposito, F.; Tortorella, E.; Subko, K.; Ezeofor, C.C.; Zhang, Y.; Tabudravu, J.; Jaspars, M.; Fani, R.; et al. Antimicrobial Activity of Monoramnholipids Produced by Bacterial Strains Isolated from the Ross Sea (Antarctica). Mar. Drugs 2016, 14, 83. [Google Scholar] [CrossRef] [PubMed]
- Tomova, I.; Stoilova-Disheva, M.; Lazarkevich, I.; Vasileva-Tonkova, E. Antimicrobial Activity and Resistance to Heavy Metals and Antibiotics of Heterotrophic Bacteria Isolated from Sediment and Soil Samples Collected from Two Antarctic Islands. Front. Life Sci. 2015, 8, 348–357. [Google Scholar] [CrossRef]
- Baquero, F.; Lanza, V.F.; Baquero, M.-R.; del Campo, R.; Bravo-Vázquez, D.A. Microcins in Enterobacteriaceae: Peptide Antimicrobials in the Eco-Active Intestinal Chemosphere. Front. Microbiol. 2019, 10, 2261. [Google Scholar] [CrossRef]
- Indira, M.; Venkateswarulu, T.C.; Vidya Prabhakar, K.; Abraham Peele, K.; Krupanidhi, S. Isolation and Characterization of Bacteriocin Producing Enterococcus Casseliflavus and Its Antagonistic Effect on Pseudomonas Aeruginosa. Karbala Int. J. Mod. Sci. 2018, 4, 361–368. [Google Scholar] [CrossRef]
- Kumariya, R.; Garsa, A.K.; Rajput, Y.S.; Sood, S.K.; Akhtar, N.; Patel, S. Bacteriocins: Classification, Synthesis, Mechanism of Action and Resistance Development in Food Spoilage Causing Bacteria. Microb. Pathog. 2019, 128, 171–177. [Google Scholar] [CrossRef]
- Calisto, N.; Navarro, L.; Orellana, P.; Wiese, G.; Gómez, C.; Cortés-Cortés, P.; Gutiérrez-Moraga, A.; Gidekel, M.; Corsini, G. Resistencia a antibióticos y actividad antimicrobiana de aislados bacterianos de suelo antártico. An. Inst. Patagon. 2021, 49, 1–15. [Google Scholar] [CrossRef]
- Holt, J.G.; Krieg, N.R.; Sneath, P.H.A.; Staley, J.T.; Williams, S.T. Bergey’s Manual of Determinative Bacteriology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 1994; ISBN 978-0-683-00603-2. [Google Scholar]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent Updates and New Developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed]
- Koeuth, T.; Versalovic, J.; Lupski, J.R. Differential Subsequence Conservation of Interspersed Repetitive Streptococcus Pneumoniae BOX Elements in Diverse Bacteria. Genome Res. 1995, 5, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Corsini, G.; Karahanian, E.; Tello, M.; Fernandez, K.; Rivero, D.; Saavedra, J.M.; Ferrer, A. Purification and Characterization of the Antimicrobial Peptide Microcin N: Properties of the Antimicrobial Peptide Microcin N. FEMS Microbiol. Lett. 2010, 312, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Daba, G.M.; Mostafa, F.A.; Saleh, S.A.A.; Elkhateeb, W.A.; Awad, G.; Nomiyama, T.; Zendo, T.; El-Dein, A.N. Purification, Amino Acid Sequence, and Characterization of Bacteriocin GA15, a Novel Class IIa Bacteriocin Secreted by Lactiplantibacillus Plantarum GCNRC_GA15. Int. J. Biol. Macromol. 2022, 213, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Montero, K.; Barrientos, L. Advances in Antarctic Research for Antimicrobial Discovery: A Comprehensive Narrative Review of Bacteria from Antarctic Environments as Potential Sources of Novel Antibiotic Compounds Against Human Pathogens and Microorganisms of Industrial Importance. Antibiotics 2018, 7, 90. [Google Scholar] [CrossRef]
- Silva, M.B.; Feitosa, A.O.; Lima, I.G.O.; Bispo, J.R.S.; Santos, A.C.M.; Moreira, M.S.A.; Câmara, P.E.A.S.; Rosa, L.H.; Oliveira, V.M.; Duarte, A.W.F.; et al. Antarctic Organisms as a Source of Antimicrobial Compounds: A Patent Review. Acad. Bras. Cienc. 2022, 94, e20210840. [Google Scholar] [CrossRef]
- Lugtenberg, B.; Kamilova, F. Plant-Growth-Promoting Rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef]
- Rodríguez-Rojas, F.; Tapia, P.; Castro-Nallar, E.; Undabarrena, A.; Muñoz-Díaz, P.; Arenas-Salinas, M.; Díaz-Vásquez, W.; Valdés, J.; Vásquez, C. Draft Genome Sequence of a Multi-Metal Resistant Bacterium Pseudomonas Putida ATH-43 Isolated from Greenwich Island, Antarctica. Front. Microbiol. 2016, 7, 1777. [Google Scholar] [CrossRef]
- See-Too, W.S.; Salazar, S.; Ee, R.; Convey, P.; Chan, K.-G.; Peix, Á. Pseudomonas Versuta Sp. Nov., Isolated from Antarctic Soil. Syst. Appl. Microbiol. 2017, 40, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Spiers, A.J.; Buckling, A.; Rainey, P.B. The Causes of Pseudomonas Diversity. Microbiology 2000, 146, 2345–2350. [Google Scholar] [CrossRef] [PubMed]
- Ghequire, M.G.K.; De Mot, R. Ribosomally Encoded Antibacterial Proteins and Peptides from Pseudomonas. FEMS Microbiol. Rev. 2014, 38, 523–568. [Google Scholar] [CrossRef]
- Smith, J.J.; Tow, L.A.; Stafford, W.; Cary, C.; Cowan, D.A. Bacterial Diversity in Three Different Antarctic Cold Desert Mineral Soils. Microb. Ecol. 2006, 51, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.; Tam, H.; Alias, S.; González, M.; González-Rocha, G.; Domínguez-Yévenes, M. Pseudomonas and Pedobacter Isolates from King George Island Inhibited the Growth of Foodborne Pathogens. Pol. Polar Res. 2011, 32, 3–14. [Google Scholar] [CrossRef]
- García-Echauri, S.A.; Gidekel, M.; Gutiérrez-Moraga, A.; Santos, L.; De León-Rodríguez, A. Isolation and Phylogenetic Classification of Culturable Psychrophilic Prokaryotes from the Collins Glacier in the Antarctica. Folia Microbiol. 2011, 56, 209–214. [Google Scholar] [CrossRef]
- De Souza, M.-J.; Nair, S.; Loka Bharathi, P.A.; Chandramohan, D. Metal and Antibiotic-Resistance in Psychrotrophic Bacteria from Antarctic Marine Waters. Ecotoxicology 2006, 15, 379–384. [Google Scholar] [CrossRef]
- Marcoleta, A.E.; Arros, P.; Varas, M.A.; Costa, J.; Rojas-Salgado, J.; Berríos-Pastén, C.; Tapia-Fuentes, S.; Silva, D.; Fierro, J.; Canales, N.; et al. The Highly Diverse Antarctic Peninsula Soil Microbiota as a Source of Novel Resistance Genes. Sci. Total Environ. 2022, 810, 152003. [Google Scholar] [CrossRef]
- Na, G.; Zhang, W.; Gao, H.; Wang, C.; Li, R.; Zhao, F.; Zhang, K.; Hou, C. Occurrence and Antibacterial Resistance of Culturable Antibiotic-Resistant Bacteria in the Fildes Peninsula, Antarctica. Mar. Pollut. Bull. 2021, 162, 111829. [Google Scholar] [CrossRef]
- Lo Giudice, A.; Brilli, M.; Bruni, V.; De Domenico, M.; Fani, R.; Michaud, L. Bacterium-Bacterium Inhibitory Interactions among Psychrotrophic Bacteria Isolated from Antarctic Seawater (Terra Nova Bay, Ross Sea): Antagonism among Psychrotrophic Antarctic Marine Bacteria. FEMS Microbiol. Ecol. 2007, 60, 383–396. [Google Scholar] [CrossRef]
- Bratchkova, A.; Ivanova, V. Bioactive Metabolites Produced by Microorganisms Collected in Antarctica and the Arctic. Biotechnol. Biotechnol. Equip. 2011, 25, 1–7. [Google Scholar] [CrossRef]
- Orellana, P.; Pavón, A.; Calisto, N.; Wiese, G.; Navarro, L.; Cortés-Cortés, P.; Gidekel, M.; Gutiérrez-Moraga, A.; Corsini, G. Identificación de bacterias antárticas con actividad antimicrobiana aisladas de la rizósfera de Deschampsia antarctica Desv. An. Inst. Patagon. 2022, 50, 11. [Google Scholar] [CrossRef]
- O’Brien, A.; Sharp, R.; Russell, N.J.; Roller, S. Antarctic Bacteria Inhibit Growth of Food-Borne Microorganisms at Low Temperatures. FEMS Microbiol. Ecol. 2004, 48, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Gradisteanu Pircalabioru, G.; Popa, L.I.; Marutescu, L.; Gheorghe, I.; Popa, M.; Czobor Barbu, I.; Cristescu, R.; Chifiriuc, M.-C. Bacteriocins in the Era of Antibiotic Resistance: Rising to the Challenge. Pharmaceutics 2021, 13, 196. [Google Scholar] [CrossRef]
- Nishie, M.; Nagao, J.-I.; Sonomoto, K. Antibacterial Peptides “Bacteriocins”: An Overview of Their Diverse Characteristics and Applications. Biocontrol. Sci. 2012, 17, 1–16. [Google Scholar] [CrossRef]
- Walsh, C.J.; Guinane, C.M.; Hill, C.; Ross, R.P.; O’Toole, P.W.; Cotter, P.D. In Silico Identification of Bacteriocin Gene Clusters in the Gastrointestinal Tract, Based on the Human Microbiome Project’s Reference Genome Database. BMC Microbiol. 2015, 15, 183. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biochemical Test | C1-4-7, D2-4-6, and M1-4-11 |
|---|---|
| Catalase | + |
| Oxidase | + |
| Urease | + |
| Use of citrate as a carbon source | + |
| Growth on Cetrimide | + |
| Glucose Fermentation | − |
| Fructose Fermentation | − |
| Sucrose Fermentation | − |
| Lactose Fermentation | − |
| Antarctic Bacteria | Closest Related Species | Identity | Accession ID |
|---|---|---|---|
| C1-4-7 ID: OR839092 | Pseudomonas marginalis pv. Marginalis | 99.93% | MK123849.1 |
| Pseudomonas fluorescens NBRC 15840 | AB680979.1 | ||
| Pseudomonas rhodesiae BDNA-E25 | MN636657.1 | ||
| Pseudomonas sp. G.S.34 | MT890191.1 | ||
| D2-4-6 ID: OR839091 | Pseudomonas sp. J380 | 100% | CP043060.1 |
| Pseudomonas azotoformans P20-L3 | LC769466.1 | ||
| Pseudomonas carnis UCD_MED7 | ON595689.1 | ||
| Pseudomonas azotoformans PCRB17a | ON564745.1 | ||
| M1-4-11 ID: OR839093 | Pseudomonas marginalis pv. Marginalis | 99.93% | MK123849.1 |
| Pseudomonas fluorescens NBRC 15840 | AB680979.1 | ||
| Pseudomonas rhodesiae BDNA-E25 | MN636657.1 | ||
| Pseudomonas sp. W15Feb39 | EU681022.1 |
| Antibiotics | μg | C1-4-7 | D2-4-6 | M1-4-11 | |
|---|---|---|---|---|---|
| Penicillins | Ampicillin | 10 | R | R | R |
| Amoxicillin-Clavulanic Acid | 10–20 | R | R | R | |
| Piperacillin-Tazobactam | 10–100 | S | S | S | |
| Cephalosporins | Cefuroxime | 30 | R | R | R |
| Ceftazidime | 30 | S | S | S | |
| Cefepime | 30 | S | I | I | |
| Cefotaxime | 30 | R | R | R | |
| Ceftriaxone | 30 | R | S | R | |
| Aminoglycosides | Amikacin | 30 | S | S | S |
| Gentamicin | 10 | S | S | S | |
| Kanamycin | 30 | S | S | S | |
| Streptomycin | 10 | I | R | S | |
| Quinolones | Ciprofloxacin | 5 | S | S | S |
| Nalidixic acid | 30 | S | I | S | |
| Levofloxacin | 5 | S | S | S | |
| Sulfonamides | Sulfafurazole | 300 | R | R | I |
| Cotrimoxazole | 25 | R | I | R | |
| Carbapenems | Ertapenem | 10 | R | R | R |
| Meropenem | 10 | S | R | S | |
| Imipenem | 10 | S | S | S | |
| Others | Trimethoprim | 5 | R | R | R |
| Chloramphenicol | 30 | R | R | R | |
| Tetracycline | 30 | S | I | S | |
| Bacitracin | 0.04 U | R | R | R |
| Pathogenic Bacteria | C1-4-7 | D2-4-6 | M1-4-11 | ||||
|---|---|---|---|---|---|---|---|
| Bacteria | Cell-Free Supernatant * | Bacteria | Cell-Free Supernatant * | Bacteria | Cell-Free Supernatant * | ||
| Gram positive | Staphylococcus saprophyticus | − | − | + | + | + | + |
| Micrococcus sp. | − | − | + | + | + | + | |
| Bacillus cereus | − | − | − | − | + | + | |
| Staphylococcus aureus ATCC 25923 | − | − | + | + | + | + | |
| Staphylococcus aureus ATCC 6835 | + | − | − | − | + | + | |
| Staphylococcus epidermidis | − | − | − | − | + | + | |
| Enterococcus faecium | − | − | − | − | + | + | |
| Streptococcus agalactiae | − | + | + | + | + | + | |
| Gram negative | Salmonella enterica serotype Enteritidis | + | + | − | − | + | + |
| Klebsiella oxytocic | + | − | − | − | + | + | |
| Vibrio parahaemolyticus VpKX | + | + | − | − | + | + | |
| Klebsiella pneumoniae | + | + | − | − | + | + | |
| Proteus vulgaris | − | − | − | + | + | + | |
| Pseudomonas aeruginosa O400 | − | + | − | + | + | + | |
| Escherichia coli XL-1 Blue | + | + | − | + | + | + | |
| Pseudomonas aeruginosa PAO1 | − | + | − | + | + | + | |
| Enterobacter aerogenes | + | − | − | − | + | + | |
| Citrobacter freundii | − | − | − | − | + | + | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calisto, N.; Navarro, L.; Iribarren, C.; Orellana, P.; Gómez, C.; Salazar, L.; Gutiérrez, A.; Aranda, C.; González, A.R.; Tello, M.; et al. Characterization of Antibiotic-Resistance Antarctic Pseudomonas That Produce Bacteriocin-like Compounds. Microorganisms 2024, 12, 530. https://doi.org/10.3390/microorganisms12030530
Calisto N, Navarro L, Iribarren C, Orellana P, Gómez C, Salazar L, Gutiérrez A, Aranda C, González AR, Tello M, et al. Characterization of Antibiotic-Resistance Antarctic Pseudomonas That Produce Bacteriocin-like Compounds. Microorganisms. 2024; 12(3):530. https://doi.org/10.3390/microorganisms12030530
Chicago/Turabian StyleCalisto, Nancy, Laura Navarro, Cristian Iribarren, Paz Orellana, Claudio Gómez, Lorena Salazar, Ana Gutiérrez, Carlos Aranda, Alex R. González, Mario Tello, and et al. 2024. "Characterization of Antibiotic-Resistance Antarctic Pseudomonas That Produce Bacteriocin-like Compounds" Microorganisms 12, no. 3: 530. https://doi.org/10.3390/microorganisms12030530
APA StyleCalisto, N., Navarro, L., Iribarren, C., Orellana, P., Gómez, C., Salazar, L., Gutiérrez, A., Aranda, C., González, A. R., Tello, M., Cortés-Cortés, P., Gidekel, M., & Corsini, G. (2024). Characterization of Antibiotic-Resistance Antarctic Pseudomonas That Produce Bacteriocin-like Compounds. Microorganisms, 12(3), 530. https://doi.org/10.3390/microorganisms12030530