
Active Microbiota of Penaeus stylirostris Larvae: Partially Shaped via Vertical and Horizontal Transmissions and Larval Ontogeny

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design, Samples Collection and Storage
2.2. Quotidian Determination of Larval Survival Rates and Larval Stages
2.3. RNA Extractions, Reverse Transcriptions, Sequencing and Sequence Processing
2.4. Downstream Microbial Analysis
3. Results
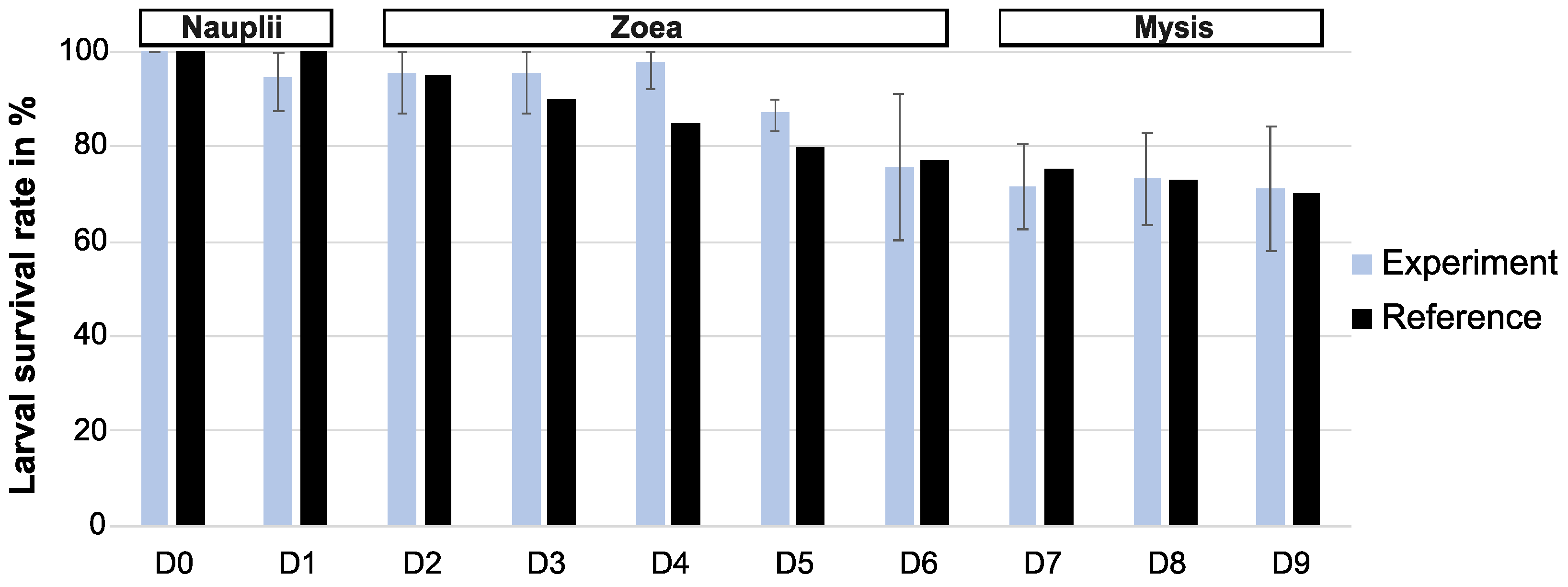
3.1. Zootechnical Parameters
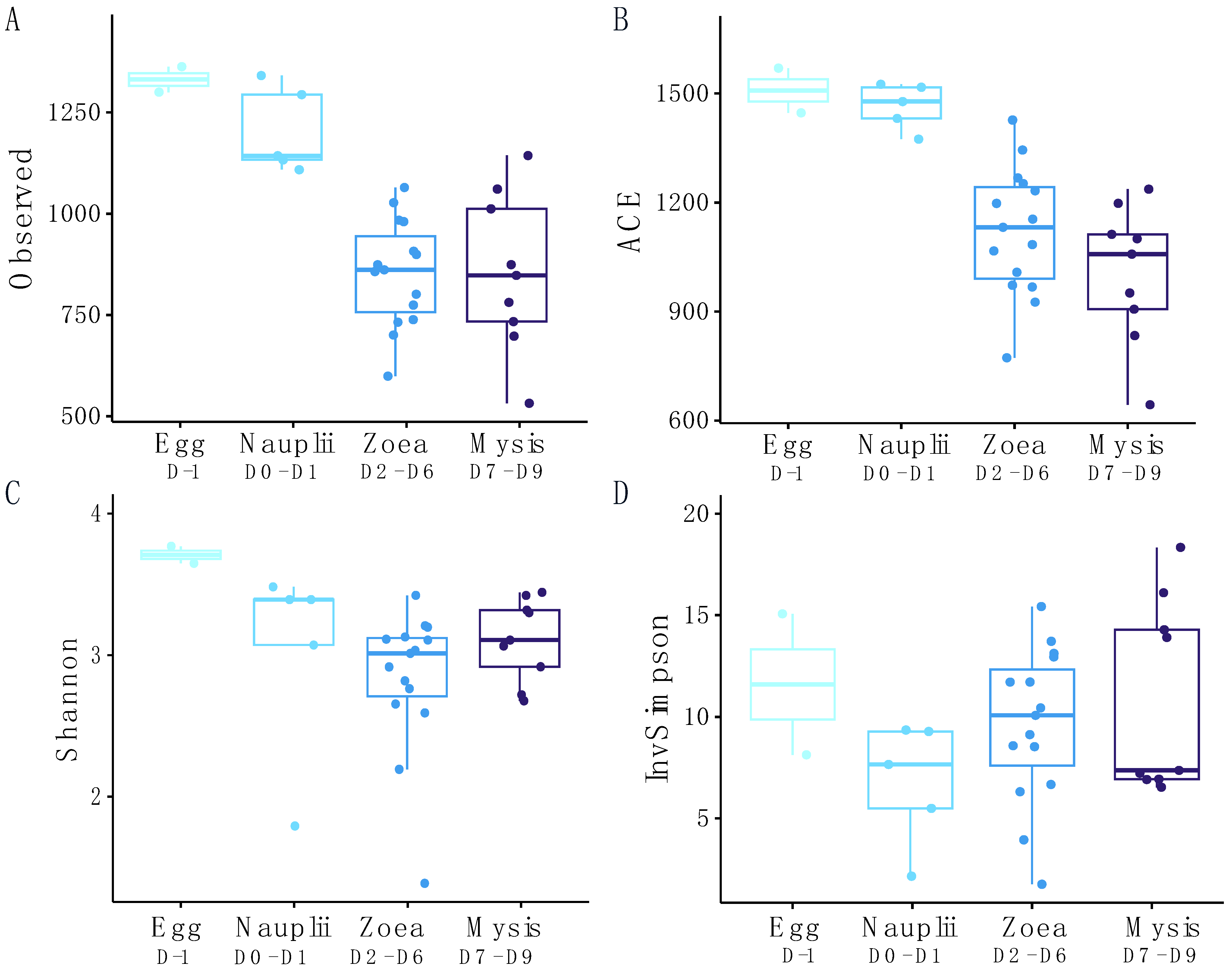
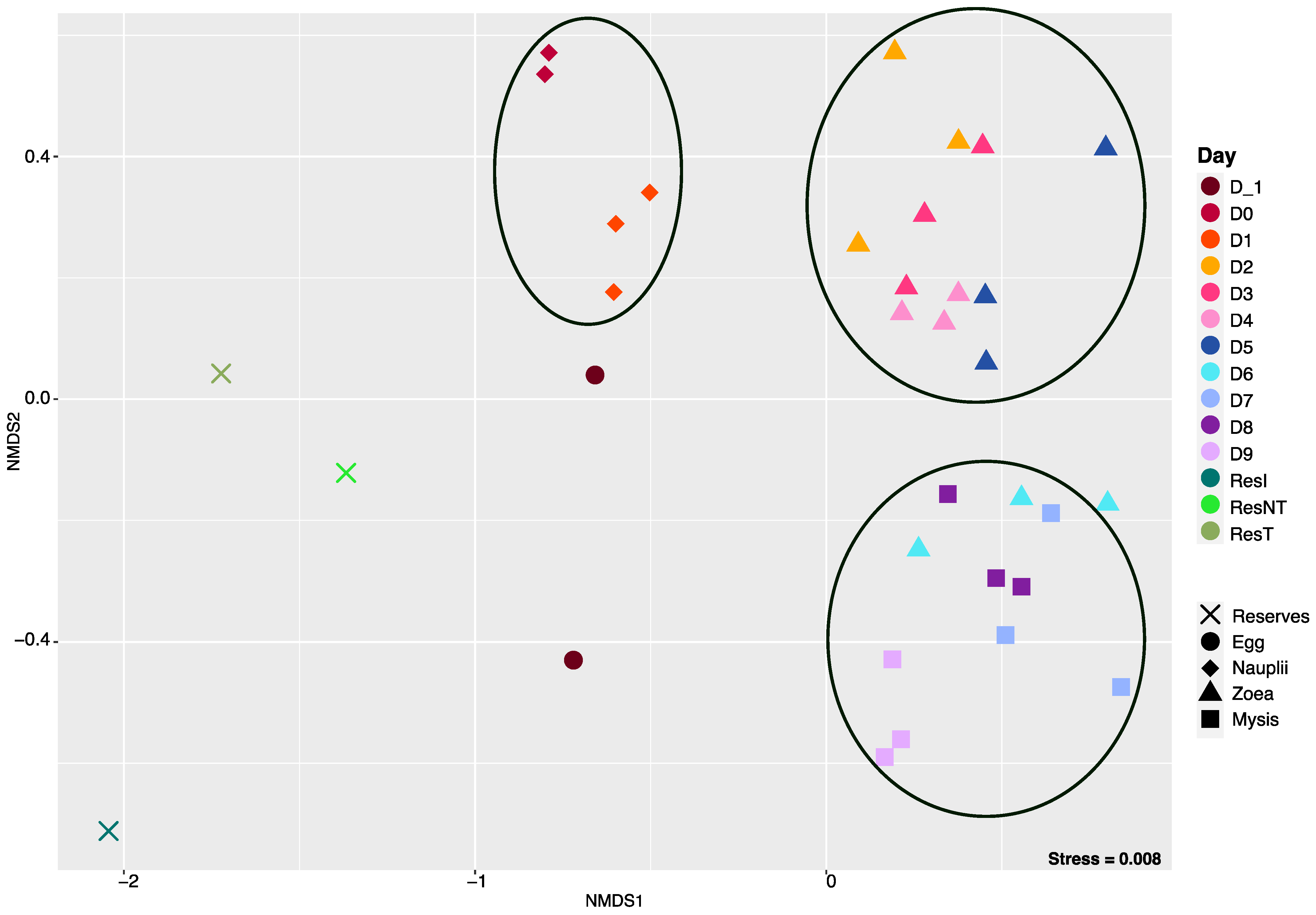
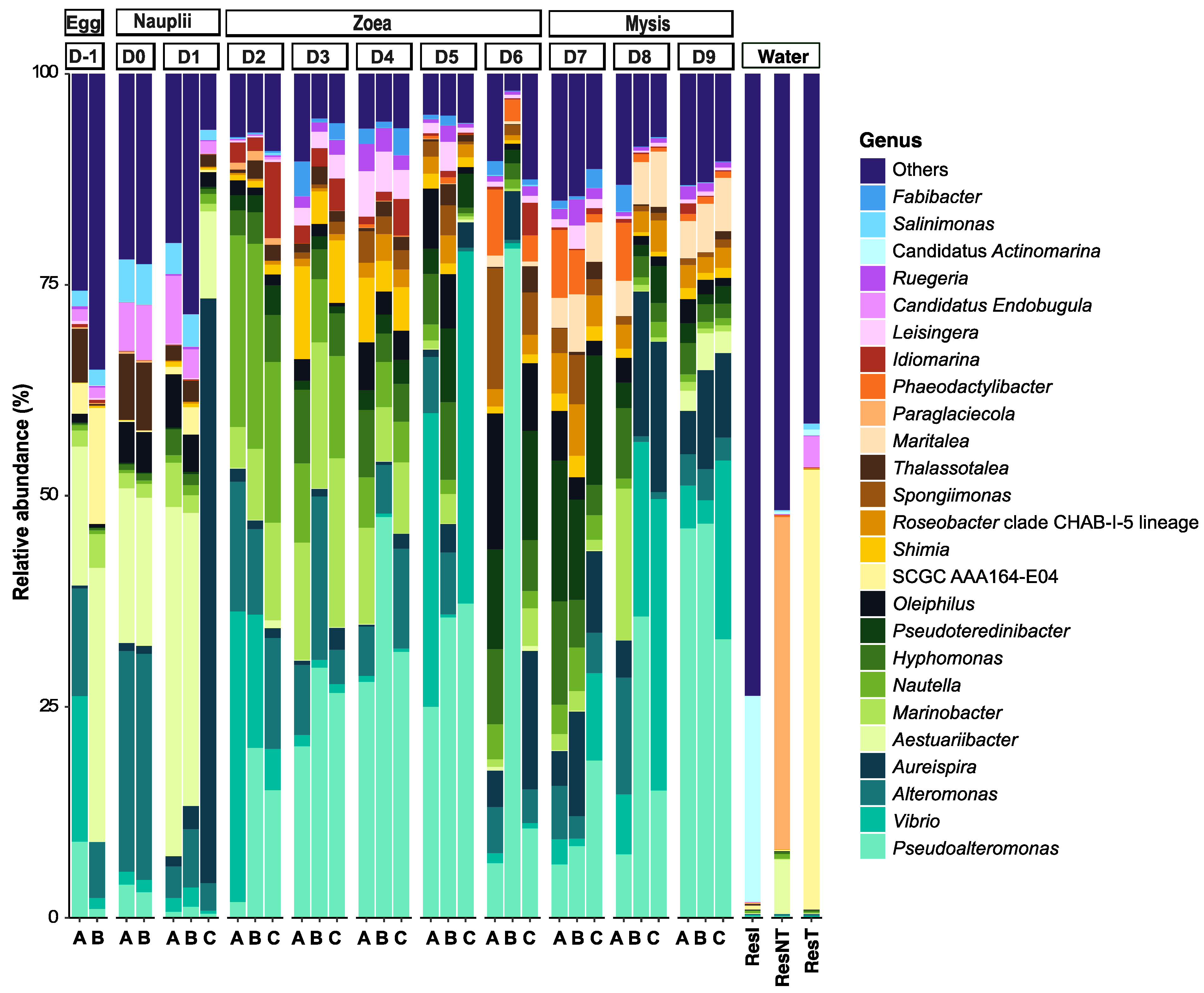
3.2. Dynamic of the Active Microbial Diversity Associated with the Larvae
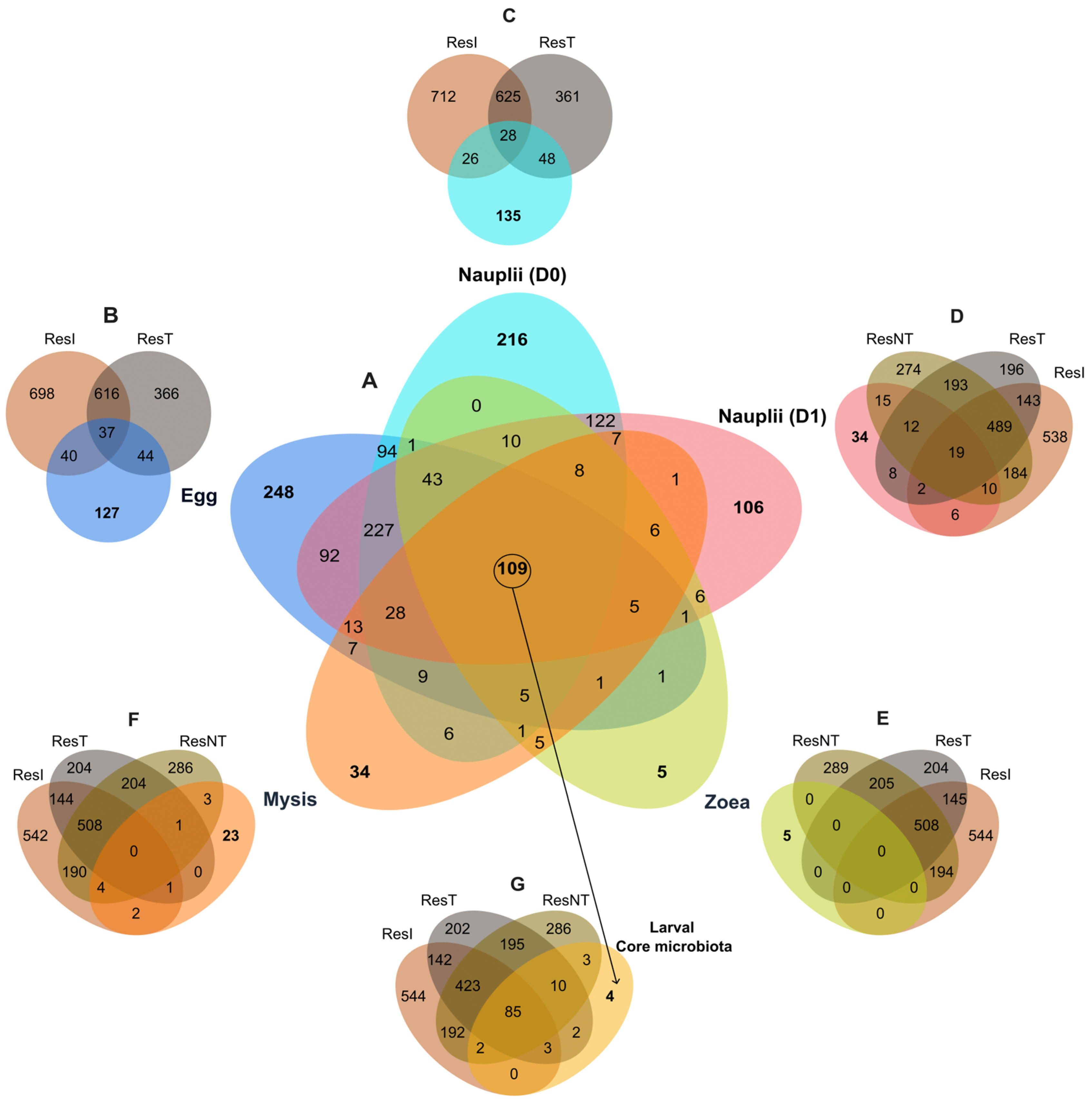
3.3. Specific and Core Microbiotas Associated with the Larval Stages and the Reservoirs
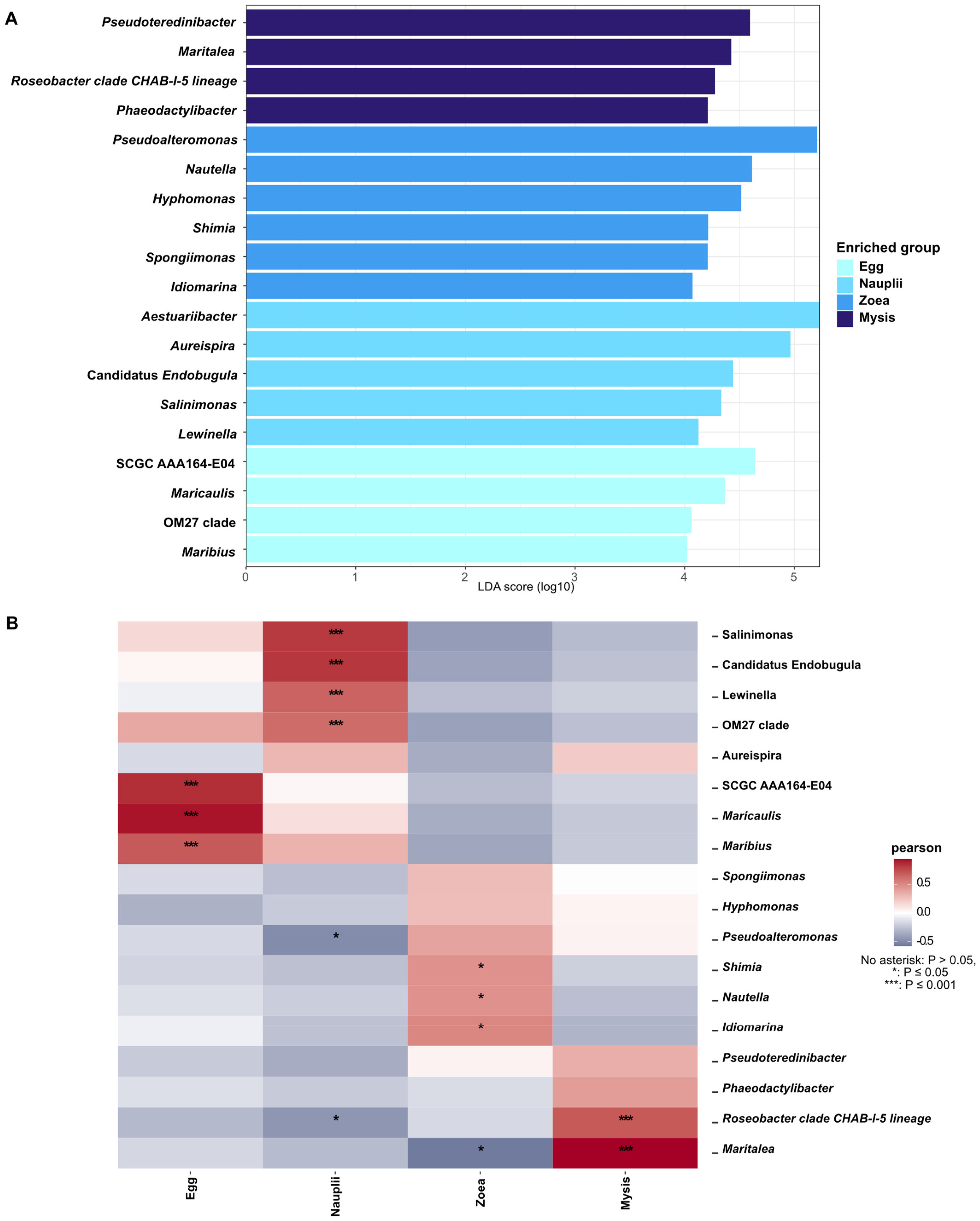
3.4. Biomarkers Associated with the Larval Stages
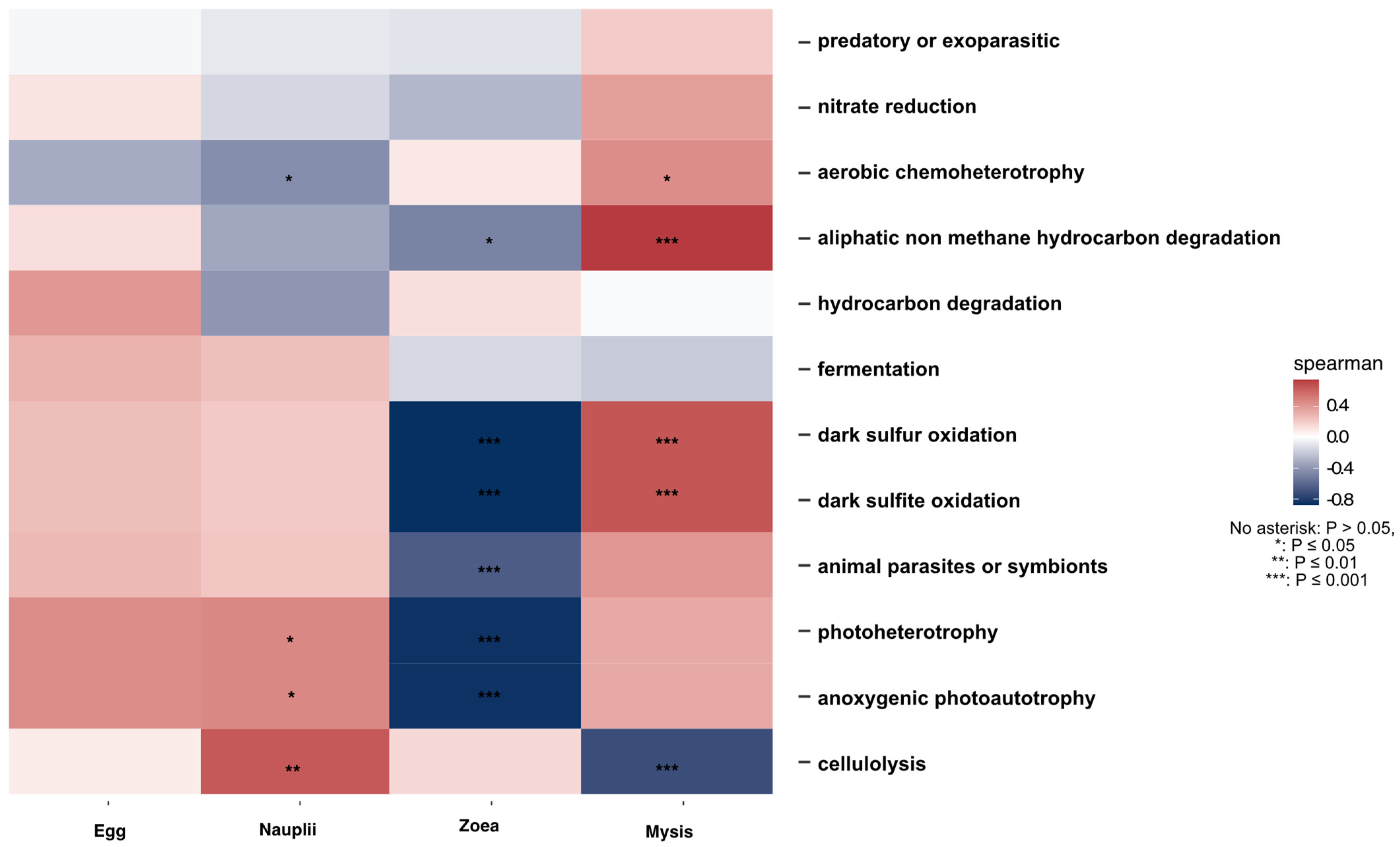
3.5. Putative Functions Associated with the Genera of the Specific and Core Microbiota of the Larvae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zheng, Y.; Yu, M.; Liu, J.; Qiao, Y.; Wang, L.; Li, Z.; Zhang, X.H.; Yu, M. Bacterial Community Associated with Healthy and Diseased Pacific White Shrimp (Litopenaeus vannamei) Larvae and Rearing Water across Different Growth Stages. Front. Microbiol. 2017, 8, 244804. [Google Scholar] [CrossRef]
- Sun, F.; Wang, Y.; Wang, C.; Zhang, L.; Tu, K.; Zheng, Z. Insights into the Intestinal Microbiota of Several Aquatic Organisms and Association with the Surrounding Environment. Aquaculture 2019, 507, 196–202. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, K.; Huang, L.; Dong, P.; Wang, S.; Chen, H.; Lu, Z.; Hou, D.; Zhang, D. Fine-Scale Succession Patterns and Assembly Mechanisms of Bacterial Community of Litopenaeus vannamei Larvae across the Developmental Cycle. Microbiome 2020, 8, 106. [Google Scholar] [CrossRef] [PubMed]
- Goarant, C.; Reynaud, Y.; Ansquer, D.; de Decker, S.; Saulnier, D.; Le Roux, F. Molecular Epidemiology of Vibrio nigripulchritudo, a Pathogen of Cultured Penaeid Shrimp (Litopenaeus stylirostris) in New Caledonia. Syst. Appl. Microbiol. 2006, 29, 570–580. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Gil, B.; Roque, A.; Turnbull, J.F. The Use and Selection of Probiotic Bacteria for Use in the Culture of Larval Aquatic Organisms. Aquaculture 2000, 191, 259–270. [Google Scholar] [CrossRef]
- Mookherjee, A.; Jurkevitch, E. Interactions between Bdellovibrio and like Organisms and Bacteria in Biofilms: Beyond Predator–Prey Dynamics. Environ. Microbiol. 2022, 24, 998–1011. [Google Scholar] [CrossRef]
- Sehnal, L.; Brammer-Robbins, E.; Wormington, A.M.; Blaha, L.; Bisesi, J.; Larkin, I.; Martyniuk, C.J.; Simonin, M.; Adamovsky, O. Microbiome Composition and Function in Aquatic Vertebrates: Small Organisms Making Big Impacts on Aquatic Animal Health. Front. Microbiol. 2021, 12, 567408. [Google Scholar] [CrossRef]
- Pratte, Z.A.; Besson, M.; Hollman, R.D.; Stewarta, F.J. The Gills of Reef Fish Support a Distinct Microbiome Influenced by Hostspecific Factors. Appl. Environ. Microbiol. 2018, 84, e00063-18. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, D.; Xu, W.; Tang, R.; Li, L. Microbiome of Co-Cultured Fish Exhibits Host Selection and Niche Differentiation at the Organ Scale. Front. Microbiol. 2019, 10, 481769. [Google Scholar] [CrossRef] [PubMed]
- Baedke, J.; Fábregas-Tejeda, A.; Nieves Delgado, A. The Holobiont Concept before Margulis. J. Exp. Zool. Part B Mol. Dev. Evol. 2020, 334, 149–155. [Google Scholar] [CrossRef]
- Singh, B.K.; Liu, H.; Trivedi, P. Eco-Holobiont: A New Concept to Identify Drivers of Host-Associated Microorganisms. Environ. Microbiol. 2020, 22, 564–567. [Google Scholar] [CrossRef]
- Ansaldo, E.; Farley, T.K.; Belkaid, Y. Control of Immunity by the Microbiota. Annu. Rev. Immunol. 2021, 39, 449–479. [Google Scholar] [CrossRef]
- Sommer, F.; Bäckhed, F. The Gut Microbiota-Masters of Host Development and Physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar] [CrossRef]
- Díaz-Sánchez, S.; Hernández-Jarguín, A.; Torina, A.; Fernández de Mera, I.G.; Estrada-Peña, A.; Villar, M.; La Russa, F.; Blanda, V.; Vicente, J.; Caracappa, S.; et al. Biotic and Abiotic Factors Shape the Microbiota of Wild-Caught Populations of the Arbovirus Vector Culicoides imicola. Insect Mol. Biol. 2018, 27, 847–861. [Google Scholar] [CrossRef]
- Agler, M.T.; Ruhe, J.; Kroll, S.; Morhenn, C.; Kim, S.T.; Weigel, D.; Kemen, E.M. Microbial Hub Taxa Link Host and Abiotic Factors to Plant Microbiome Variation. PLoS Biol. 2016, 14, e1002352. [Google Scholar] [CrossRef] [PubMed]
- Foster, K.R.; Schluter, J.; Coyte, K.Z.; Rakoff-Nahoum, S. The Evolution of the Host Microbiome as an Ecosystem on a Leash. Nature 2017, 548, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Garibay-Valdez, E.; Martínez-Porchas, M.; Calderón, K.; Vargas-Albores, F.; Gollas-Galván, T.; Martínez-Córdova, L. Taxonomic and Functional Changes in the Microbiota of the White Shrimp (Litopenaeus vannamei) Associated with Postlarval Ontogenetic Development. Aquaculture 2020, 518, 734842. [Google Scholar] [CrossRef]
- Imaizumi, K.; Tinwongger, S.; Kondo, H.; Hirono, I. Analysis of Microbiota in the Stomach and Midgut of Two Penaeid Shrimps during Probiotic Feeding. Sci. Rep. 2021, 11, 9936. [Google Scholar] [CrossRef]
- Giraud, C.; Callac, N.; Boulo, V.; Lam, J.-S.J.S.; Pham, D.; Selmaoui-Folcher, N.; Wabete, N. The Active Microbiota of the Eggs and the Nauplii of the Pacific Blue Shrimp Litopenaeus stylirostris Partially Shaped by a Potential Vertical Transmission. Front. Microbiol. 2022, 13, 886752. [Google Scholar] [CrossRef] [PubMed]
- Giraud, C.; Callac, N.; Beauvais, M.; Mailliez, J.-R.J.-R.; Ansquer, D.; Selmaoui-Folcher, N.; Pham, D.; Wabete, N.; Boulo, V. Potential Lineage Transmission within the Active Microbiota of the Eggs and the Nauplii of the Shrimp Litopenaeus stylirostris: Possible Influence of the Rearing Water and More. PeerJ 2021, 9, e12241. [Google Scholar] [CrossRef]
- Zhou, J.; Ning, D. Stochastic Community Assembly: Does It Matter in Microbial Ecology. Microbiol. Mol. Biol. Rev. 2017, 81, 10–128. [Google Scholar] [CrossRef]
- Van Der Gast, C.J.; Ager, D.; Lilley, A.K. Temporal Scaling of Bacterial Taxa Is Influenced by Both Stochastic and Deterministic Ecological Factors. Environ. Microbiol. 2008, 10, 1411–1418. [Google Scholar] [CrossRef]
- Zhang, W.; Zhu, Z.; Chen, J.; Qiu, Q.; Xiong, J. Quantifying the Importance of Abiotic and Biotic Factors Governing the Succession of Gut Microbiota Over Shrimp Ontogeny. Front. Microbiol. 2021, 12, 752750. [Google Scholar] [CrossRef] [PubMed]
- FAO the State of World Fisheries and Aquaculture 2022. Towards Blue Transformation. 2022. Available online: https://www.fao.org/documents/card/en/c/cc0461en (accessed on 19 December 2013).
- Goarant, C.; Merien, F. Quantification of Vibrio penaeicida, the Etiological Agent of Syndrome 93 in New Caledonian Shrimp, by Real-Time PCR Using SYBR Green I Chemistry. J. Microbiol. Methods 2006, 67, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Pham, D.; Ansquer, D.; Chevalier, A.; Dauga, C.; Peyramale, A.; Wabete, N.; Labreuche, Y. Selection and Characterization of Potential Probiotic Bacteria for Litopenaeus stylirostris Shrimp Hatcheries in New Caledonia. Aquaculture 2014, 432, 475–482. [Google Scholar] [CrossRef]
- Callac, N.; Boulo, V.; Giraud, C.; Beauvais, M.; Ansquer, D.; Ballan, V.; Maillez, J.-R.; Wabete, N.; Pham, D. Microbiota of the Rearing Water of Penaeus stylirostris Larvae Influenced by Lagoon Seawater and Specific Key Microbial Lineages of Larval Stage and Survival. Microbiol. Spectr. 2022, 10, e04241-22. [Google Scholar] [CrossRef] [PubMed]
- Callac, N.; Giraud, C.; Boulo, V.; Wabete, N.; Pham, D. Microbial Biomarker Detection in Shrimp Larvae Rearing Water as Putative Bio-Surveillance Proxies in Shrimp Aquaculture. PeerJ 2023, 11, e15201. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global Patterns of 16S RRNA Diversity at a Depth of Millions of Sequences per Sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef]
- Hugerth, L.W.; Wefer, H.A.; Lundin, S.; Jakobsson, H.E.; Lindberg, M.; Rodin, S.; Engstrand, L.; Andersson, A.F. DegePrime, a Program for Degenerate Primer Design for Broad-Taxonomic-Range PCR in Microbial Ecology Studies. Appl. Environ. Microbiol. 2014, 80, 5116–5123. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Wickham, H.; Chang, W. Package Ggplot2: An Implementation of the Grammar of Graphics. Creat. Elegant Data Vis. Using Gramm. Graph. 2016, 2, 1–189. [Google Scholar]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic Biomarker Discovery and Explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Dong, Q.; Wang, D.; Zhang, P.; Liu, Y.; Niu, C. Microbiome Marker: An R/Bioconductor package for microbiome marker identification and visualization. Bioinformatics 2022, 38, 4027–4029. [Google Scholar] [CrossRef]
- Liu, C.; Cui, Y.; Li, X.; Yao, M. Microeco: An R Package for Data Mining in Microbial Community Ecology. FEMS Microbiol. Ecol. 2021, 97, fiaa255. [Google Scholar] [CrossRef]
- Bardou, P.; Mariette, J.; Escudié, F.; Djemiel, C.; Klopp, C. SOFTWARE Open Access Jvenn: An Interactive Venn Diagram Viewer. BMC Bioinform. 2014, 15, 293. [Google Scholar] [CrossRef]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling Function and Taxonomy in the Global Ocean Microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef]
- Colette, M.; Guentas, L.; Patrona, L.D.; Ansquer, D.; Callac, N. Dynamic of Active Microbial Diversity in Rhizosphere Sediments of Halophytes Used for Bioremediation of Earthen Shrimp Ponds. Environ. Microbiome 2023, 18, 58. [Google Scholar] [CrossRef] [PubMed]
- Sansupa, C.; Wahdan, S.F.M.; Hossen, S.; Disayathanoowat, T.; Wubet, T.; Purahong, W. Can We Use Functional Annotation of Prokaryotic Taxa (FAPROTAX) to Assign the Ecological Functions of Soil Bacteria? Appl. Sci. 2021, 11, 688. [Google Scholar] [CrossRef]
- Cristescu, M.E. Can Environmental RNA Revolutionize Biodiversity Science? Trends Ecol. Evol. 2019, 34, 694–697. [Google Scholar] [CrossRef] [PubMed]
- Marshall, N.T.; Vanderploeg, H.A.; Chaganti, S.R. Environmental (e)RNA Advances the Reliability of EDNA by Predicting Its Age. Sci. Rep. 2021, 11, 2769. [Google Scholar] [CrossRef] [PubMed]
- Jesser, K.J.; Noble, R.T. Vibrio ecology in the Neuse River Estuary, North Carolina, characterized by next-generation amplicon sequencing of the gene encoding heat shock protein 60 (hsp60). Appl. Environ. Microbiol. 2018, 84, e00333-18. [Google Scholar] [CrossRef] [PubMed]
- Miyata, K.; Inoue, Y.; Amano, Y.; Nishioka, T.; Yamane, M.; Kawaguchi, T.; Morita, O.; Honda, H. Fish Environmental RNA Enables Precise Ecological Surveys with High Positive Predictivity. Ecol. Indic. 2021, 128, 107796. [Google Scholar] [CrossRef]
- Vinay, T.N.; Patil, P.K.; Aravind, R.; Anand, P.S.S.; Baskaran, V.; Balasubramanian, C.P. Microbial Community Composition Associated with Early Developmental Stages of the Indian White Shrimp, Penaeus indicus. Mol. Genet. Genom. 2022, 297, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Huang, J.; Wang, P.; Li, T. Insights into the Microbiota of Larval and Postlarval Pacific White Shrimp (Penaeus vannamei) along Early Developmental Stages: A Case in Pond Level. Mol. Genet. Genom. 2020, 295, 1517–1528. [Google Scholar] [CrossRef]
- Xue, M.; Wu, L.; He, Y.; Liang, H.; Wen, C. Biases during DNA Extraction Affect Characterization of the Microbiota Associated with Larvae of the Pacific White Shrimp, Litopenaeus vannamei. PeerJ 2018, 2018, e5257. [Google Scholar] [CrossRef]
- Chim, L.; Brun, H.L.; Le Moullac, G. Marine Shrimp Farming. Fish. Aquac. J. 2009, 4, 28–54. [Google Scholar]
- Methou, P.; Hernández-Ávila, I.; Aube, J.; Cueff-Gauchard, V.; Gayet, N.; Amand, L.; Shillito, B.; Pradillon, F.; Cambon-Bonavita, M.A. Is It First the Egg or the Shrimp?—Diversity and Variation in Microbial Communities Colonizing Broods of the Vent Shrimp Rimicaris exoculata during Embryonic Development. Front. Microbiol. 2019, 10, 441978. [Google Scholar] [CrossRef] [PubMed]
- Guri, M.; Durand, L.; Cueff-Gauchard, V.; Zbinden, M.; Crassous, P.; Shillito, B.; Cambon-Bonavita, M.A. Acquisition of Epibiotic Bacteria along the Life Cycle of the Hydrothermal Shrimp Rimicaris exoculata. ISME J. 2012, 6, 597–609. [Google Scholar] [CrossRef] [PubMed]
- Craig Cary, S.; Giovannoni, S.J. Transovarial Inheritance of Endosymbiotic Bacteria in Clams Inhabiting Deep-Sea Hydrothermal Vents and Cold Seeps. Proc. Natl. Acad. Sci. USA 1993, 90, 5695–5699. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, S.; Weisz, J.B.; Lindquist, N.; Hentschel, U. Vertical Transmission of a Phylogenetically Complex Microbial Consortium in the Viviparous Sponge Ircinia felix. Appl. Environ. Microbiol. 2007, 73, 2067–2078. [Google Scholar] [CrossRef] [PubMed]
- Nyholm, S.V. In the Beginning: Egg-Microbe Interactions and Consequences for Animal Hosts. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190593. [Google Scholar] [CrossRef] [PubMed]
- Lauzon, H.L.; Gudmundsdottir, S.; Petursdottir, S.K.; Reynisson, E.; Steinarsson, A.; Oddgeirsson, M.; Bjornsdottir, R.; Gudmundsdottir, B.K. Microbiota of Atlantic Cod (Gadus morhua L.) Rearing Systems at Pre- and Posthatch Stages and the Effect of Different Treatments. J. Appl. Microbiol. 2010, 109, 1775–1789. [Google Scholar] [CrossRef] [PubMed]
- Kesarcodi-Watson, A.; Kaspar, H.; Lategan, M.J.; Gibson, L. Alteromonas macleodii 0444 and Neptunomonas sp. 0536, Two Novel Probiotics for Hatchery-Reared GreenshellTM Mussel Larvae, Perna canaliculus. Aquaculture 2010, 309, 49–55. [Google Scholar] [CrossRef]
- Huang, F.; Pan, L.; Song, M.; Tian, C.; Gao, S. Microbiota Assemblages of Water, Sediment, and Intestine and Their Associations with Environmental Factors and Shrimp Physiological Health. Appl. Microbiol. Biotechnol. 2018, 102, 8585–8598. [Google Scholar] [CrossRef]
- Suantika, G.; Aditiawati, P.; Indriani Astuti, D.; Khotimah, Z.F. The Use of Indigenous Probiotic Halomonas aquamarina and Shewanella algae for White Shrimp (Litopenaeus vannamei Boone) Hatchery Productivity in Zero Water Discharge System. J. Aquac. Res. Dev. 2013, 4, 5. [Google Scholar] [CrossRef]
- Sorieul, L.; Rückert, C.; Al-Dilaimi, A.; Winkler, A.; Kalinowski, J.; Pham, D.; Wabete, N.; Boulo, V. Whole-Genome Sequence of Pseudoalteromonas sp. NC201, a Probiotic Strain for Litopenaeus stylirostris Hatcheries in New Caledonia. Microbiol. Resour. Announc. 2019, 8, e00477-19. [Google Scholar] [CrossRef]
- Gromek, S.M.; Suria, A.M.; Fullmer, M.S.; Garcia, J.L.; Gogarten, J.P.; Nyholm, S.V.; Balunas, M.J. Leisingera sp. JC1, a Bacterial Isolate from Hawaiian Bobtail Squid Eggs, Produces Indigoidine and Differentially Inhibits Vibrios. Front. Microbiol. 2016, 7, 210965. [Google Scholar] [CrossRef]
- Abd El-Rhman, A.M.; Khattab, Y.A.E.; Shalaby, A.M.E. Micrococcus luteus and Pseudomonas Species as Probiotics for Promoting the Growth Performance and Health of Nile Tilapia, Oreochromis niloticus. Fish Shellfish Immunol. 2009, 27, 175–180. [Google Scholar] [CrossRef]
- Koga, A.; Goto, M.; Hayashi, S.; Yamamoto, S.; Miyasaka, H. Probiotic Effects of a Marine Purple Non-Sulfur Bacterium, Rhodovulum sulfidophilum KKMI01, on Kuruma Shrimp (Marsupenaeus japonicus). Microorganisms 2022, 10, 244. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, R.; Miura, N.; Ito, M.; Takagi, T.; Yamashiro, H.; Nishikawa, Y.; Nishimura, Y.; Kobayashi, K.; Kataoka, M. Specific Detection of Coral-Associated Ruegeria, a Potential Probiotic Bacterium, in Corals and Subtropical Seawater. Mar. Biotechnol. 2021, 23, 576–589. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Jiang, C.; Yamano, R.; Koike, S.; Sakai, Y.; Mino, S.; Sawabe, T. Unveiling the Early Life Core Microbiome of the Sea Cucumber Apostichopus japonicus and the Unexpected Abundance of the Growth-Promoting Sulfitobacter. Anim. Microbiome 2023, 5, 54. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, G.; Hartzell, P.L.; Navaratnam, V. Calcium Is Required for Ixotrophy of Aureispira sp. CCB-QB1. Microbiology 2015, 161, 1933–1941. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Sun, X. Core Gut Microbiota of Shrimp Function as a Regulator to Maintain Immune Homeostasis in Response to WSSV Infection. Microbiol. Spectr. 2022, 10, e02465-21. [Google Scholar] [CrossRef]
- Hossain, M.S.; Dai, J.; Qiu, D. Dysbiosis of the Shrimp (Penaeus monodon) Gut Microbiome with AHPND Outbreaks Revealed by 16S RRNA Metagenomics Analysis. Aquac. Res. 2021, 52, 3336–3349. [Google Scholar] [CrossRef]
- Abraham, W. Maricaulis. In Bergey’s Manual of Systematics of Archaea and Bacteria; Wiley: Hoboken, NJ, USA, 2020; pp. 1–8. [Google Scholar] [CrossRef]
- Scott Quackenbusho, L. Yolk Synthesis in the Marine Shrimp, Penaeus vannamei. Am. Zool. 2001, 41, 458–464. [Google Scholar] [CrossRef]
- Palacios, E.; Palacios, E.; Racotta, I.S.; Racotta, I.S.; Heras, H.; Heras, H.; Moal, J.; Moal, J. Relation between Lipid and Fatty Acid Composition of Eggs and Larval Survival in White Paci c Shrimp (Penaeus vannamei, Boone, 1931). Aquac. Int. 2002, 9, 531–543. [Google Scholar] [CrossRef]
- Doberva, M.; Stien, D.; Sorres, J.; Hue, N.; Sanchez-Ferandin, S.; Eparvier, V.; Ferandin, Y.; Lebaron, P.; Lami, R. Large Diversity and Original Structures of Acyl-Homoserine Lactones in Strain MOLA 401, a Marine Rhodobacteraceae Bacterium. Front. Microbiol. 2017, 8, 254028. [Google Scholar] [CrossRef]
- Barnier, C.; Clerissi, C.; Lami, R.; Intertaglia, L.; Lebaron, P.; Grimaud, R.; Urios, L. Description of Palleronia rufa sp. nov., a Biofilm-Forming and AHL-Producing Rhodobacteraceae, Reclassification of Hwanghaeicola aestuarii as Palleronia aestuarii comb. nov., Maribius pontilimi as Palleronia pontilimi comb. nov., Maribius salinus as Palleronia pelagia comb. nov. and emended description of the genus Palleronia. Syst. Appl. Microbiol. 2020, 43, 126018. [Google Scholar] [CrossRef]
- Pajares, S. Unraveling the Distribution Patterns of Bacterioplankton in a Mesoscale Cyclonic Eddy Confined to an Oxygen-Depleted Basin. Aquat. Microb. Ecol. 2021, 87, 151–166. [Google Scholar] [CrossRef]
- Yoon, J.H.; Kang, S.J.; Lee, S.Y. Salinimonas lutimaris sp. nov., a Polysaccharide-Degrading Bacterium Isolated from a Tidal Flat. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2012, 101, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Fang, J.; Zhang, H.; Wei, Y.; Wang, L.; Liu, R.; Cao, J. Complete Genome Sequence of a Piezophilic Bacterium Salinimonas sediminis N102T, Isolated from Deep-Sea Sediment of the New Britain Trench. Mar. Genom. 2021, 56, 100807. [Google Scholar] [CrossRef]
- Gago, J.F.; Viver, T.; Urdiain, M.; Pastor, S.; Kämpfer, P.; Ferreira, E.; Rossello-Mora, R. Description of Three New Alteromonas Species Alteromonas antoniana sp. nov., Alteromonas lipotrueae sp. nov. and Alteromonas lipotrueiana sp. nov. Isolated from Marine Environments, and Proposal for Reclassification of the Genus Salinimonas as Alteromonas. Syst. Appl. Microbiol. 2021, 44, 126226. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, H.; Cao, L.; Chen, H.; Wang, M.; Lian, C.; Zhong, Z.; Li, C. Salinimonas iocasae sp. nov., a Halophilic Bacterium Isolated from a Polychaete Tube in a Hydrothermal Field. Int. J. Syst. Evol. Microbiol. 2020, 70, 3899–3904. [Google Scholar] [CrossRef]
- Li, H.; Mishra, M.; Ding, S.; Miyamoto, M.M. Diversity and Dynamics of “Candidatus Endobugula” and Other Symbiotic Bacteria in Chinese Populations of the Bryozoan, Bugula neritina. Microb. Ecol. 2019, 77, 243–256. [Google Scholar] [CrossRef]
- Lim, G.E.; Haygood, M.G. “Candidatus Endobugula Glebosa”, a Specific Bacterial Symbiont of the Marine Bryozoan Bugula simplex. Appl. Environ. Microbiol. 2004, 70, 4921–4929. [Google Scholar] [CrossRef]
- Fu, Z.; Han, F.; Huang, K.; Zhang, J.; Qin, J.G.; Chen, L.; Li, E. Combined Toxic Effects of Thiamethoxam on Intestinal Flora, Transcriptome and Physiology of Pacific White Shrimp Litopenaeus vannamei. Sci. Total Environ. 2022, 830, 154799. [Google Scholar] [CrossRef]
- Quintino-Rivera, J.G.; Elizondo-González, R.; Gamboa-Delgado, J.; Guzmán-Villanueva, L.T.; Peña-Rodriguez, A. Metabolic Turnover Rate, Digestive Enzyme Activities, and Bacterial Communities in the White Shrimp Litopenaeus vannamei under Compensatory Growth. PeerJ 2023, 11, e14747. [Google Scholar] [CrossRef]
- Barreto-Curiel, F.; Ramirez-Puebla, S.T.; Ringø, E.; Escobar-Zepeda, A.; Godoy-Lozano, E.; Vazquez-Duhalt, R.; Sanchez-Flores, A.; Viana, M.T. Effects of Extruded Aquafeed on Growth Performance and Gut Microbiome of Juvenile Totoaba macdonaldi. Anim. Feed Sci. Technol. 2018, 245, 91–103. [Google Scholar] [CrossRef]
- Zheng, Y.; Yu, M.; Liu, Y.; Su, Y.; Xu, T.; Yu, M.; Zhang, X.H. Comparison of Cultivable Bacterial Communities Associated with Pacific White Shrimp (Litopenaeus vannamei) Larvae at Different Health Statuses and Growth Stages. Aquaculture 2016, 451, 163–169. [Google Scholar] [CrossRef]
- Restrepo, L.; Domínguez-Borbor, C.; Bajaña, L.; Betancourt, I.; Rodríguez, J.; Bayot, B.; Reyes, A. Microbial Community Characterization of Shrimp Survivors to AHPND Challenge Test Treated with an Effective Shrimp Probiotic (Vibrio diabolicus). Microbiome 2021, 9, 88. [Google Scholar] [CrossRef] [PubMed]
- Donachie, S.P.; Hou, S.; Gregory, T.S.; Malahoff, A.; Alam, M. Idiomarina loihiensis sp. nov., a Halophilic γ-Proteobacterium from the Lō’ihi Submarine Volcano, Hawai’i. Int. J. Syst. Evol. Microbiol. 2003, 53, 1873–1879. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, C.; Papale, M.; Lo Giudice, A. Idiomarina sp. Isolates from Cold and Temperate Environments as Biosurfactant Producers. J. Mar. Sci. Eng. 2022, 10, 1135. [Google Scholar] [CrossRef]
- Rizzo, C.; Michaud, L.; Hörmann, B.; Gerçe, B.; Syldatk, C.; Hausmann, R.; De Domenico, E.; Lo Giudice, A. Bacteria Associated with Sabellids (Polychaeta: Annelida) as a Novel Source of Surface Active Compounds. Mar. Pollut. Bull. 2013, 70, 125–133. [Google Scholar] [CrossRef]
- Bayat, Z.; Hassanshahian, M.; Hesni, M.A. Study the Symbiotic Crude Oil-Degrading Bacteria in the Mussel Mactra stultorum Collected from the Persian Gulf. Mar. Pollut. Bull. 2016, 105, 120–124. [Google Scholar] [CrossRef]
- Vidal-Verdú, À.; Molina-Menor, E.; Satari, L.; Pascual, J.; Peretó, J.; Porcar, M. Maritalea mediterranea sp. nov., Isolated from Marine Plastic Residues. Int. J. Syst. Evol. Microbiol. 2023, 73, 005677. [Google Scholar] [CrossRef]
- Fukui, Y.; Abe, M.; Kobayashi, M.; Ishihara, K.; Oikawa, H.; Yano, Y.; Satomi, M. Maritalea porphyrae sp. nov., Isolated from a Red Alga (Porphyra yezoensis), and Transfer of Zhangella mobilis to Maritalea mobilis comb. nov. Int. J. Syst. Evol. Microbiol. 2011, 62, 43–48. [Google Scholar] [CrossRef]
- Hwang, C.Y.; Cho, K.D.; Yih, W.; Cho, B.C. Maritalea myrionectae gen. nov., sp. nov., Isolated from a Culture of the Marine Ciliate Myrionecta rubra. Int. J. Syst. Evol. Microbiol. 2009, 59, 609–614. [Google Scholar] [CrossRef]
- Zhukova, N.V.; Eliseikina, M.G.; Balakirev, E.S.; Ayala, F.J. Multiple Bacterial Partners in Symbiosis with the Nudibranch Mollusk Rostanga alisae. Sci. Rep. 2022, 12, 169. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, Y.; Jiao, N.; Stepanauskas, R.; Luo, H. Ecological Genomics of the Uncultivated Marine Roseobacter Lineage CHAB-I-5. Appl. Environ. Microbiol. 2016, 82, 2100–2111. [Google Scholar] [CrossRef] [PubMed]
- Coulthurst, S.J. The Type VI Secretion System—A Widespread and Versatile Cell Targeting System. Res. Microbiol. 2013, 164, 640–654. [Google Scholar] [CrossRef]
- Griffith, R.S.; Black, H.R. Erythromycin. Med. Clin. N. Am. 1970, 54, 1199–1215. [Google Scholar] [CrossRef]
- Bakke, I.; Coward, E.; Andersen, T.; Vadstein, O. Selection in the Host Structures the Microbiota Associated with Developing Cod Larvae (Gadus Morhua). Environ. Microbiol. 2015, 17, 3914–3924. [Google Scholar] [CrossRef]
- Cicala, F.; Lago-Lestón, A.; Gomez-Gil, B.; Gollas-Galván, T.; Chong-Robles, J.; Cortés-Jacinto, E.; Martínez-Porchas, M. Gut Microbiota Shifts in the Giant Tiger Shrimp, Penaeus monodon, during the Postlarvae, Juvenile, and Adult Stages. Aquac. Int. 2020, 28, 1421–1433. [Google Scholar] [CrossRef]
- Angthong, P.; Uengwetwanit, T.; Arayamethakorn, S.; Chaitongsakul, P.; Karoonuthaisiri, N.; Rungrassamee, W. Bacterial Analysis in the Early Developmental Stages of the Black Tiger Shrimp (Penaeus monodon). Sci. Rep. 2020, 10, 4896. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compared Groups | Observed | ACE | |
|---|---|---|---|
| Egg | Nauplii | 0.631 | 0.829 |
| Egg | Zoea | 0.009 | 0.026 |
| Egg | Mysis | 0.015 | 0.007 |
| Nauplii | Zoea | 0.002 | 0.004 |
| Nauplii | Mysis | 0.007 | 0.001 |
| Zoea | Mysis | 0.882 | 0.297 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Callac, N.; Giraud, C.; Pham, D.; Ansquer, D.; Wabete, N.; Boulo, V. Active Microbiota of Penaeus stylirostris Larvae: Partially Shaped via Vertical and Horizontal Transmissions and Larval Ontogeny. Microorganisms 2024, 12, 608. https://doi.org/10.3390/microorganisms12030608
Callac N, Giraud C, Pham D, Ansquer D, Wabete N, Boulo V. Active Microbiota of Penaeus stylirostris Larvae: Partially Shaped via Vertical and Horizontal Transmissions and Larval Ontogeny. Microorganisms. 2024; 12(3):608. https://doi.org/10.3390/microorganisms12030608
Chicago/Turabian StyleCallac, Nolwenn, Carolane Giraud, Dominique Pham, Dominique Ansquer, Nelly Wabete, and Viviane Boulo. 2024. "Active Microbiota of Penaeus stylirostris Larvae: Partially Shaped via Vertical and Horizontal Transmissions and Larval Ontogeny" Microorganisms 12, no. 3: 608. https://doi.org/10.3390/microorganisms12030608
APA StyleCallac, N., Giraud, C., Pham, D., Ansquer, D., Wabete, N., & Boulo, V. (2024). Active Microbiota of Penaeus stylirostris Larvae: Partially Shaped via Vertical and Horizontal Transmissions and Larval Ontogeny. Microorganisms, 12(3), 608. https://doi.org/10.3390/microorganisms12030608