
Pathogenicity of Aeromonas veronii Isolated from Diseased Macrobrachium rosenbergii and Host Immune-Related Gene Expression Profiles

Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Signs and Bacterial Isolation
2.2. Experimental Infection
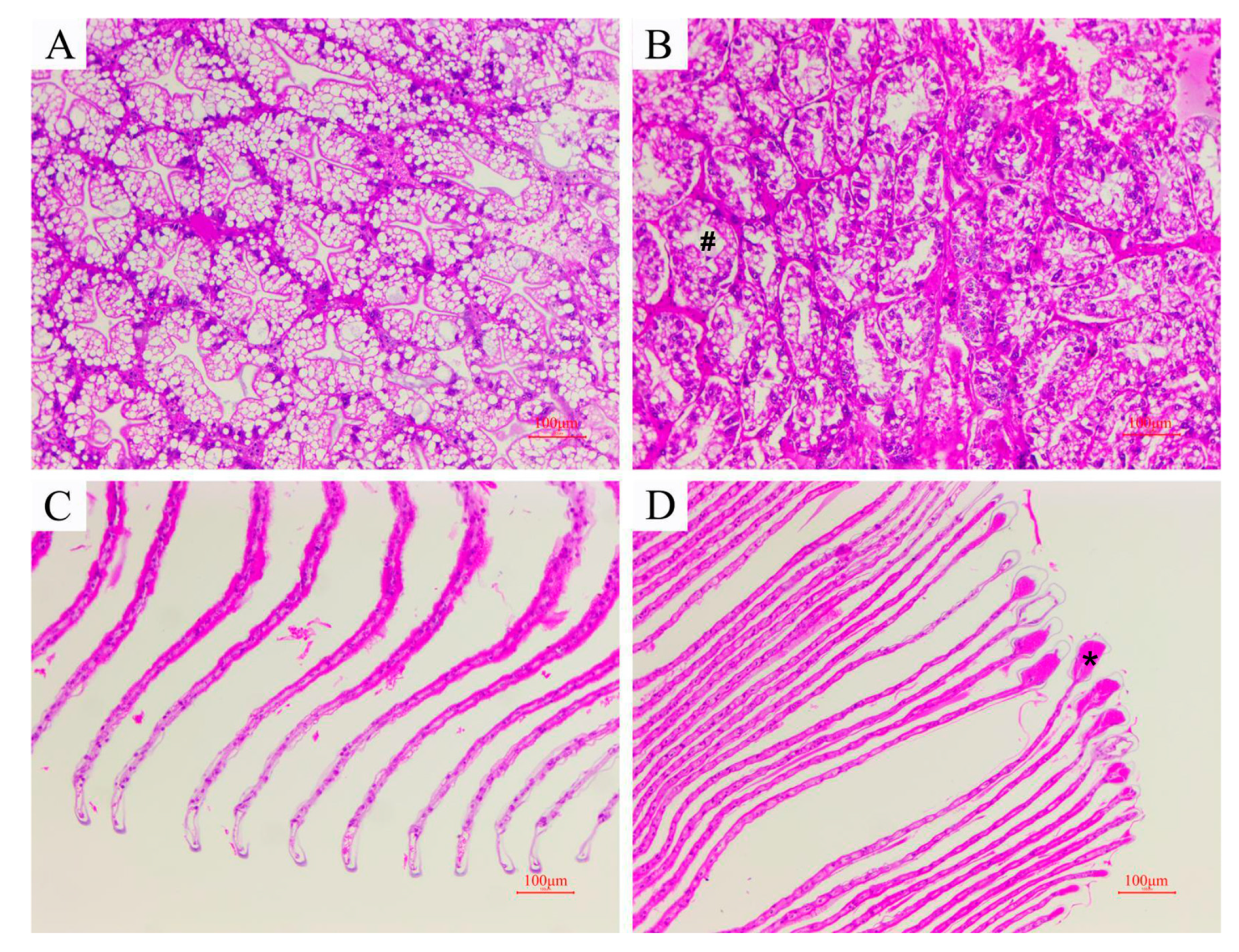
2.3. Histological Analysis
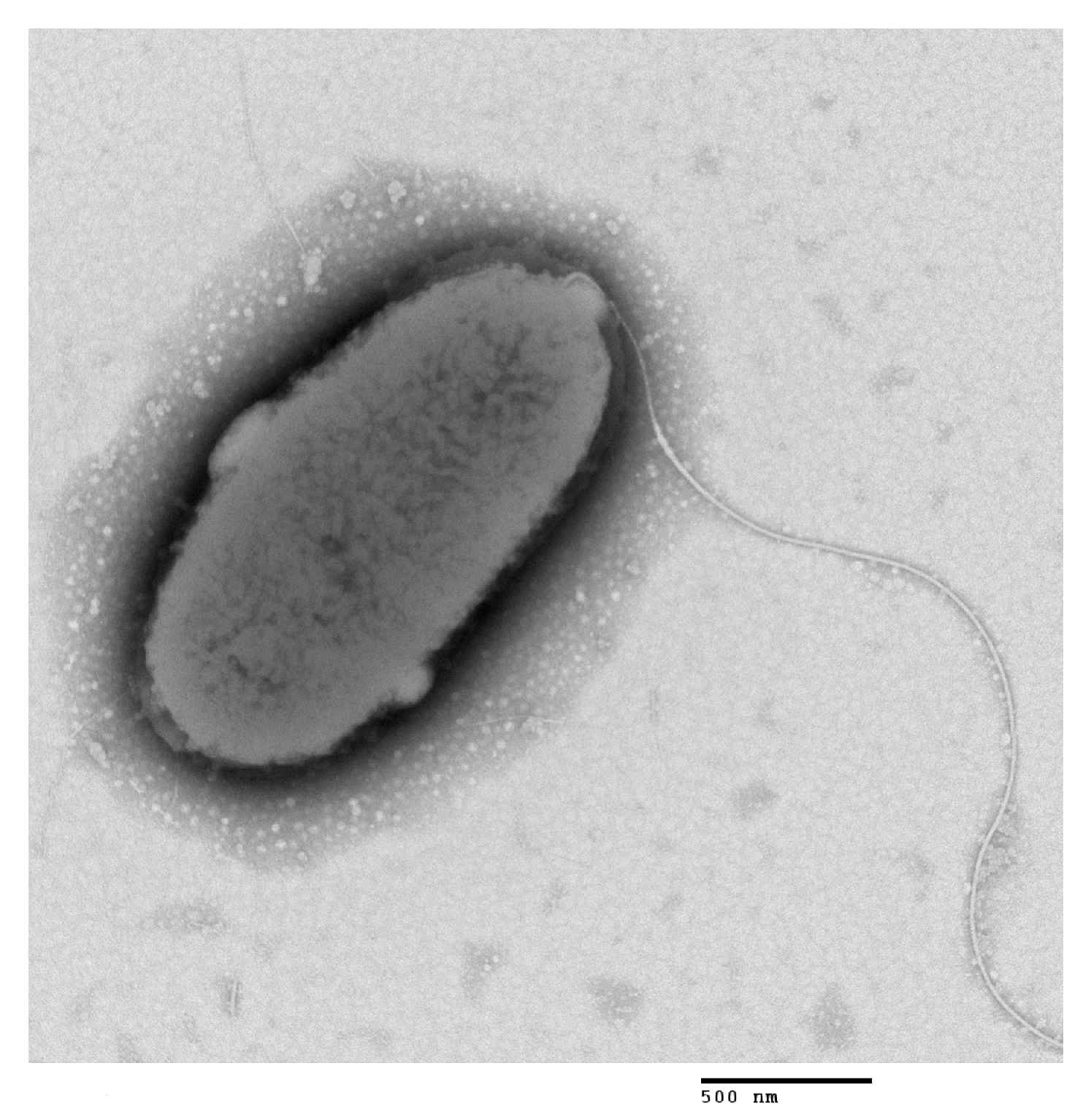
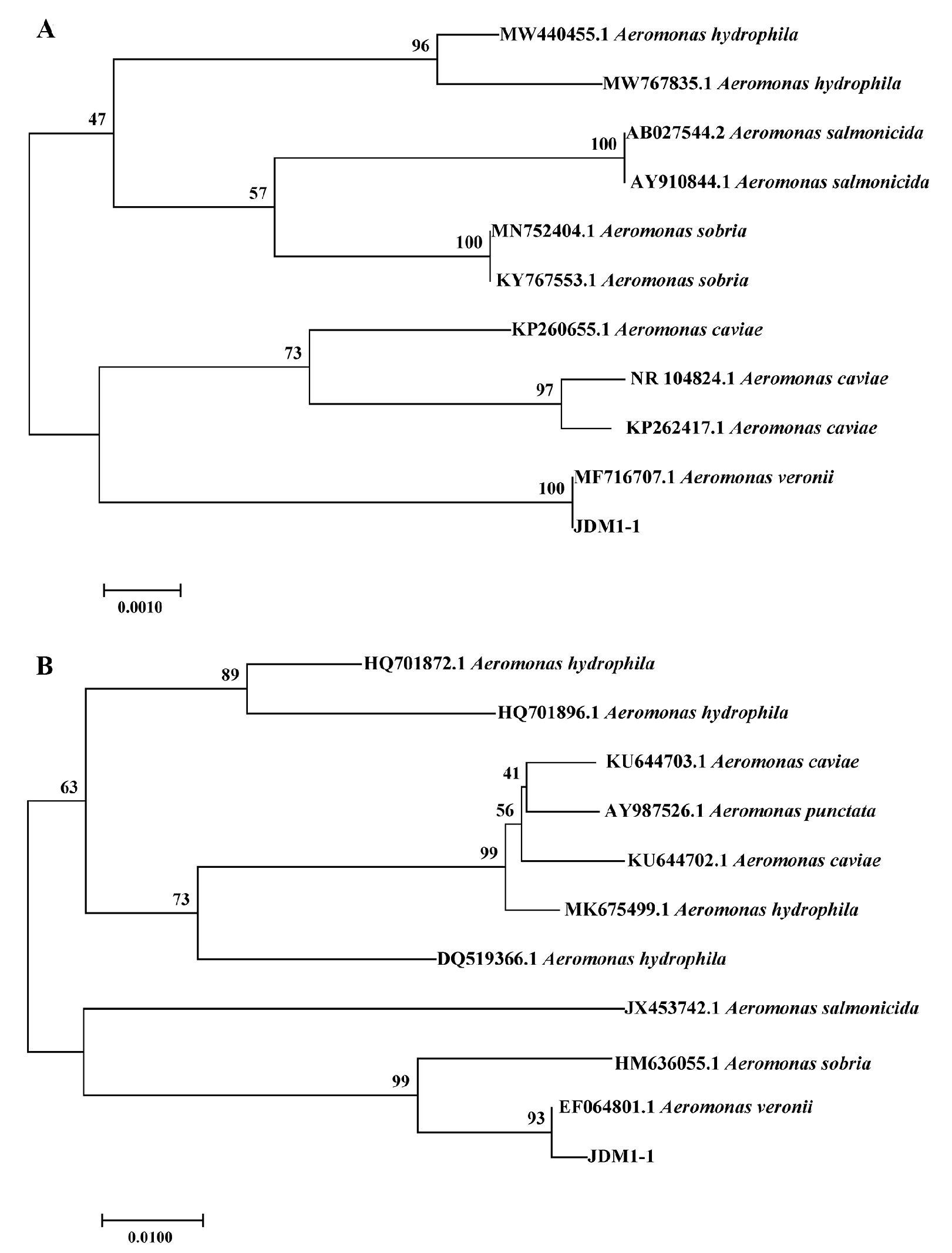
2.4. Bacterial Identification
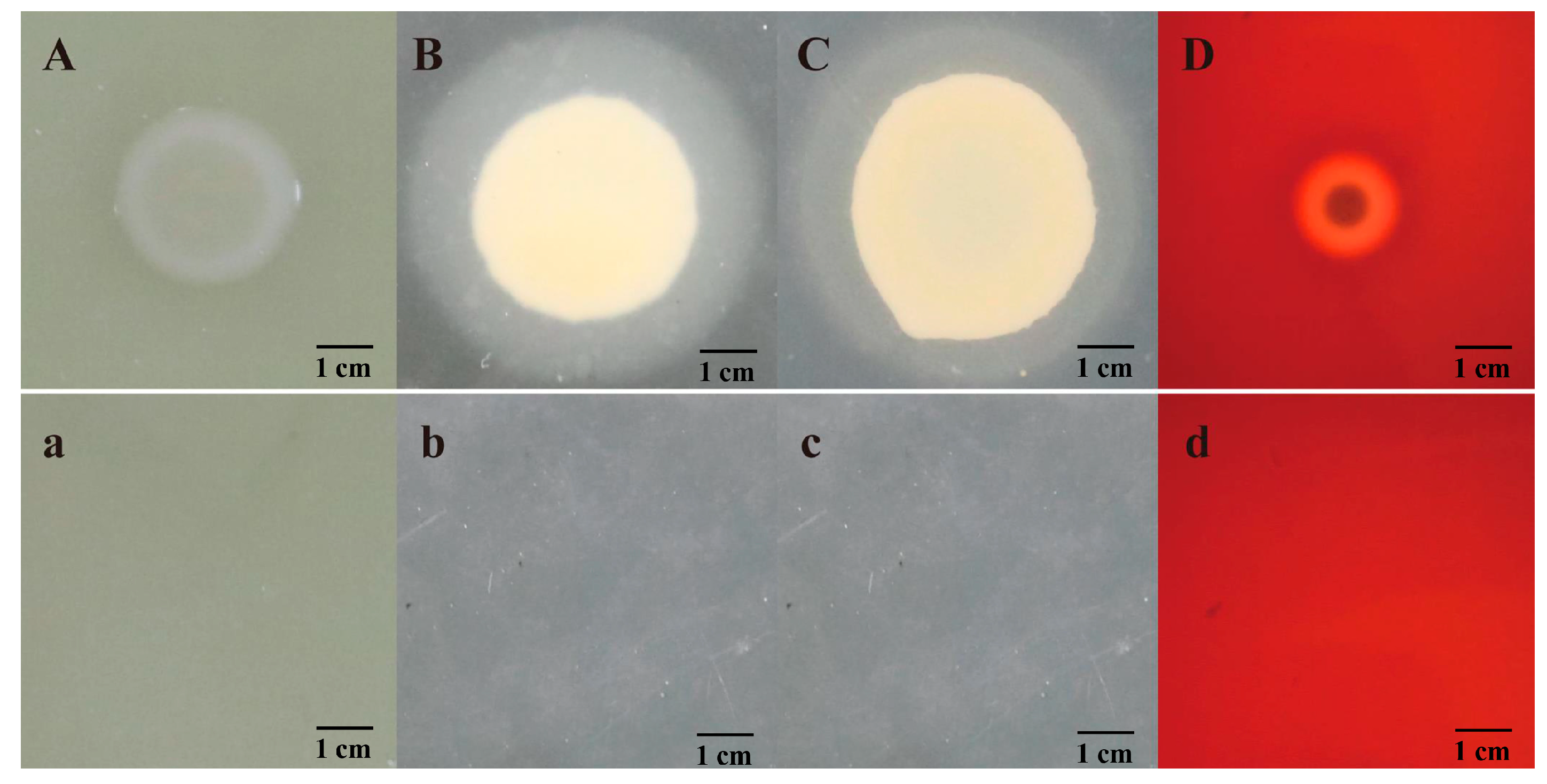
2.5. Determination of Extracellular Enzymes and Hemolysin
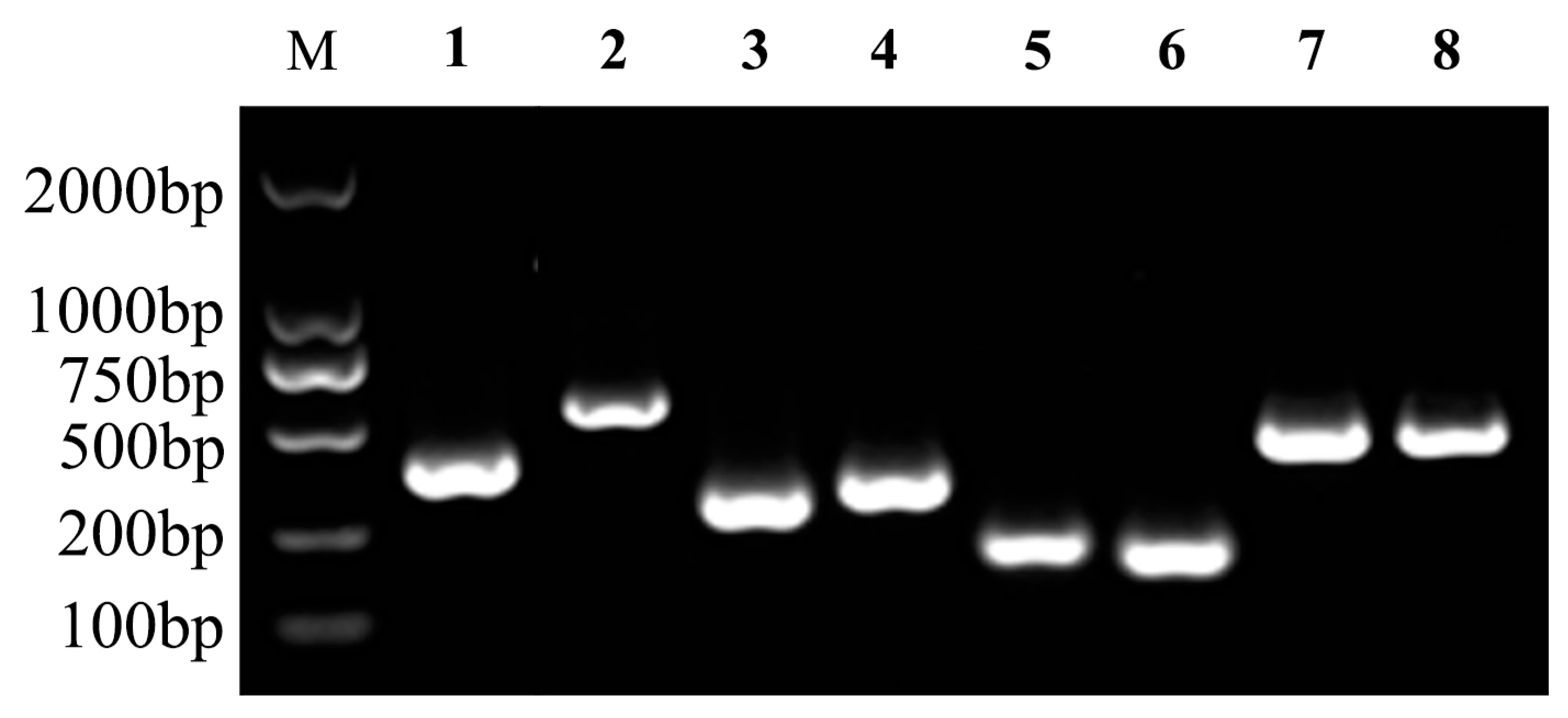
2.6. Virulence-Related Gene Detection
2.7. Detection of Immune-Related Gene Expression after Infection Using qRT-PCR
3. Results
3.1. Pathogenicity of Isolate JDM1-1
3.2. Histopathological Changes
3.3. Identification and Characterization of Isolate JDM1-1
3.4. Virulence Genes and Extracellular Products of Isolate JDM1-1
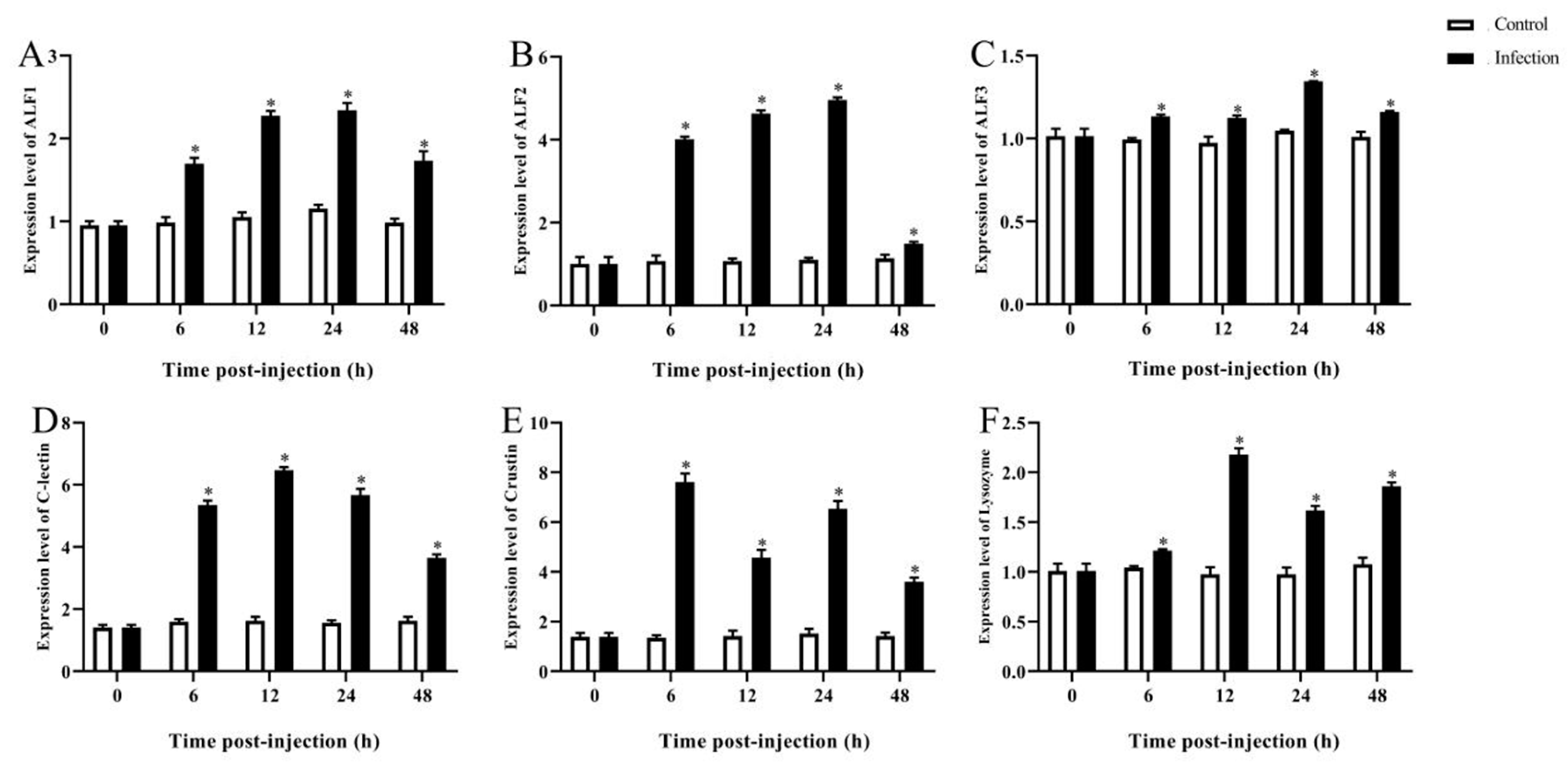
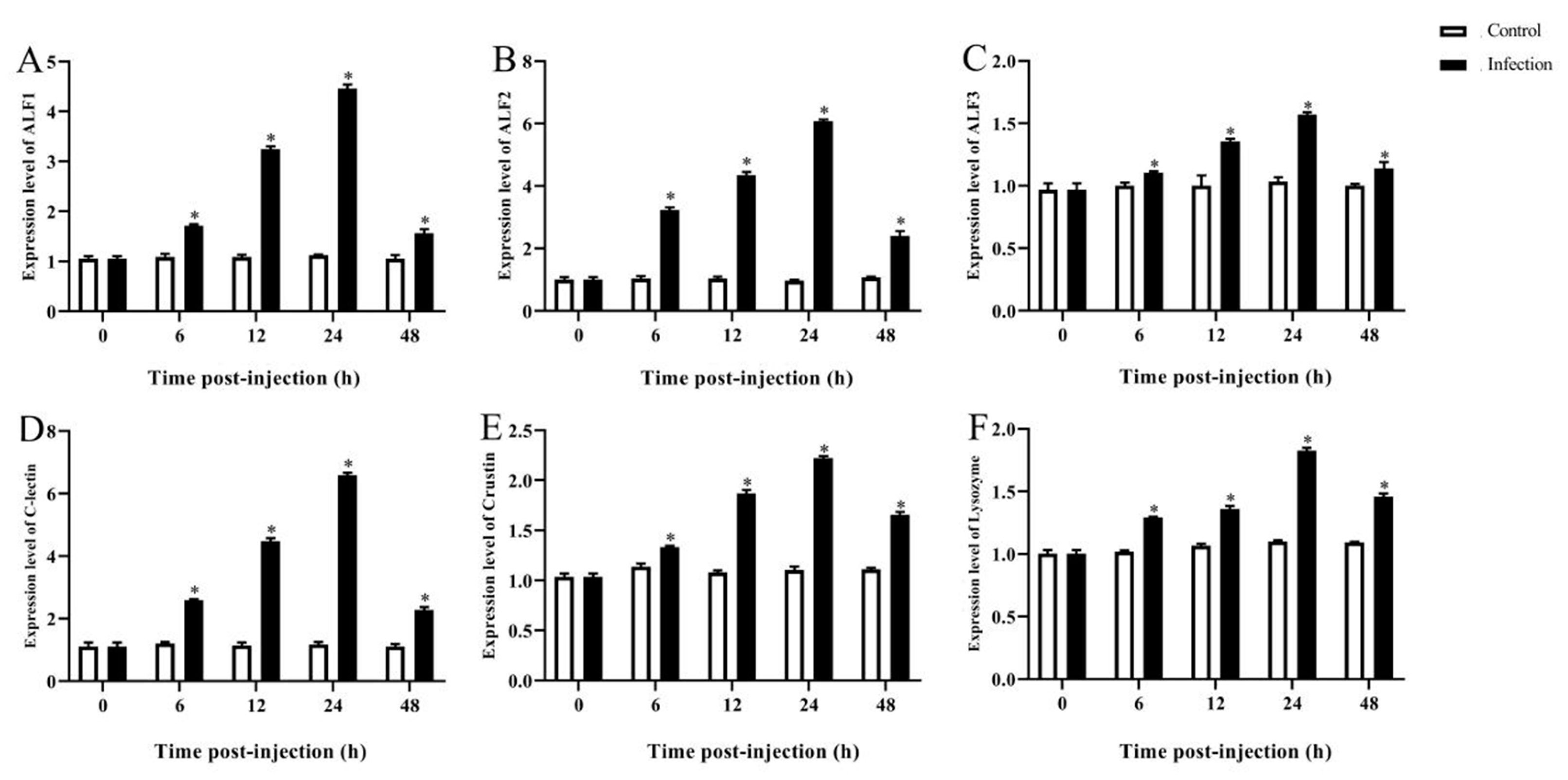
3.5. Expression Analysis of Immune-Related Genes of M. rosenbergii at Different Hours Post Infection
3.5.1. Immune-Related Gene Expression in the Hepatopancreas after A. veronii Infection
3.5.2. Immune-Related Gene Expression in the Intestines after A. veronii Infection
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fisheries Bureau, Department of Agriculture of China. 2022 China Fishery Statistical Yearbook; China Agriculture Press: Beijing, China, 2023; p. 24. [Google Scholar]
- Hooper, C.; Debnath, P.P.; Stentiford, G.D.; Bateman, K.S.; Salin, K.R.; Bass, D. Diseases of the giant river prawn Macrobrachium rosenbergii: A review for a growing industry. Rev. Aquac. 2023, 15, 738–758. [Google Scholar] [CrossRef]
- Low, C.; Yusoff, M.M.; Kuppusamy, G.; Nadzri, N.A. Molecular biology of Macrobrachium rosenbergii nodavirus infection in giant freshwater prawn. J. Fish Dis. 2018, 41, 1771–1781. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Chen, X.; Zhao, R.H.; Li, C.; Gao, W.; Zhang, Q.L.; Huang, J. Description of a natural infection with Decapod Iridescent Virus 1 in farmed giant freshwater prawn, Macrobrachium rosenbergii. Viruses 2019, 11, 354. [Google Scholar] [CrossRef]
- Ravi, M.; Nazeer Basha, A.; Taju, G.; Ram Kumar, R.; Sahul Hameed, A.S. Clearance of Macrobrachium rosenbergii nodavirus (MrNV) and extra small virus (XSV) and immunological changes in experimentally injected Macrobrachium rosenbergii. Fish Shellfish Immun. 2010, 28, 428–433. [Google Scholar] [CrossRef]
- Gangnonngiw, W.; Kiatpathomchai, W.; Sriurairatana, S.; Laisutisan, K.; Flegel, T.W. Parvo-like virus in the hepatopancreas of freshwater prawns Macrobrachium rosenbergii cultivated in Thailand. Dis. Aquat. Org. 2009, 85, 167–173. [Google Scholar] [CrossRef]
- Kiran, R.B.P.; Rajendran, K.V.; Jung, S.J.; Oh, M.J. Experimental susceptibility of different life-stages of the giant freshwater prawn, Macrobrachium rosenbergii (de Man), to white spot syndrome virus (WSSV). J. Fish Dis. 2002, 25, 201–207. [Google Scholar] [CrossRef]
- Hsieh, C.Y.; Chuang, P.C.; Chen, L.C.; Chien, T.; Chien, M.S.; Huang, K.C.; Kao, H.F.; Tung, M.C.; Tsai, S.S. Infectious hypodermal and haematopoietic necrosis virus (IHHNV) infections in giant freshwater prawn, Macrobrachium rosenbergii. Aquaculture 2006, 258, 73–79. [Google Scholar] [CrossRef]
- Khuntia, C.P.; Das, B.K.; Samantaray, B.R.; Samantaray, B.R.; Samal, S.K.; Mishra, B.K. Characterization and pathogenicity studies of Vibrio parahaemolyticus isolated from diseased freshwater prawn, Macrobrachium rosenbergii (de Man). Aquac. Res. 2008, 39, 301–310. [Google Scholar] [CrossRef]
- Jayaprakash, N.S.; Rejish Kumar, V.J.; Philip, R.; Bright Singh, I.S. Vibrios associated with Macrobrachium rosenbergii (De Man, 1879) larvae from three hatcheries on the Indian southwest coast. Aquac. Res. 2006, 37, 351–358. [Google Scholar] [CrossRef]
- Peng, X.; Zhong, Z.; Zhong, H.; Gong, J.; Du, T.; Ding, L.; Lan, X.; Tu, H.; Tang, Q.; Xia, Z.; et al. Histopathological observation and comparative transcriptome analysis reveal immune response mechanisms to Aeromonas dhakensis infection in Macrobrachium rosenbergii. Fish Shellfish Immunol. 2023, 142, 109151. [Google Scholar] [CrossRef]
- Sung, H.H.; Hwang, S.F.; Tasi, F.M. Responses of Giant Freshwater Prawn (Macrobrachium rosenbergii) to Challenge by Two Strains of Aeromonas spp. J. Invertebr. Pathol. 2000, 76, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wang, C.H. The susceptibility of the giant freshwater prawn Macrobrachium rosenbergii to Lactococcus garvieae and its resistance under copper sulfate stress. Dis. Aquat. Org. 2001, 47, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Janda, J.M.; Abbott, S.L. The genus Aeromonas: Taxonomy, Pathogenicity, and Infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef] [PubMed]
- Hoai, T.D.; Trang, T.T.; Van, T.N.; Giang, N.T.H.; Van, K.V. Aeromonas veronii caused disease and mortality in channel catfish in Vietnam. Aquaculture 2019, 513, 734425. [Google Scholar] [CrossRef]
- Mallik, S.K.; Joshi, N.; Shahi, N.; Kala, K.; Singh, S.; Giri, A.K.; Pant, K.; Chandra, S. Characterization and pathogenicity of Aeromonas veronii associated with mortality in cage farmed grass carp, Ctenopharyngodon idella (Valenciennes, 1844) from the Central Himalayan region of India. Antonie Leeuwenhoek 2020, 113, 2063–2076. [Google Scholar] [CrossRef] [PubMed]
- Raj, N.S.; Swaminathan, T.R.; Dharmaratnam, A.; Raja, S.A.; Ramraj, D.; Lal, K.K. Aeromonas veronii caused bilateral exophthalmia and mass mortality in cultured Nile tilapia, Oreochromis niloticus (L.) in India. Aquaculture 2019, 512, 734278. [Google Scholar] [CrossRef]
- Min, F.H.; Qian, M.Q.; Hao, S.Z.; Han, Z.Z.; Ran, D.Z.; Quan, T.Q.; Mu, X.X.; Ming, W.L.; Lin, Z.R.; He, B.C. Study on the cause and control of Carapace ulcer of Litopenaeus vannamei in factory freshwater aquaculture. J. Biol. 2019, 36, 65–69. [Google Scholar]
- Behreans, A.L.; Karber, L. Determination of LD50. In Screening in Pharmacology; Academic Press: New York, NY, USA, 1953; p. 60. [Google Scholar]
- Brenner, D.J.; Krieg, N.R.; Staley, J.T. Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Part B; Springer: East Lansing, MI, USA, 2021; Volume 2, p. 561. [Google Scholar]
- Zhang, X.J.; Bai, X.S.; Yan, B.L.; Bi, K.R.; Qin, L. Vibrio harveyi as a causative agent of mass mortalities of megalopa in the seed production of swimming crab Portunus trituberculatus. Aquac. Int. 2014, 22, 661–672. [Google Scholar] [CrossRef]
- Chen, Q.Y.; Zhang, Z.R.; Tang, H.Y.; Zhou, L.Y.; Ao, S.Q.; Zhou, Y.F.; Zhu, X.H.; Gao, X.J.; Jiang, Q.; Tu, C.D.; et al. Aeromonas hydrophila associated with red spot disease in Macrobrachium nipponense and host immune-related gene expression profiles. J. Invertebr. Pathol. 2021, 182, 107584. [Google Scholar] [CrossRef]
- Stentiford, G.D. Diseases of commercially exploited crustaceans: Cross-cutting issues for global fisheries and aquaculture. J. Invertebr. Pathol. 2011, 106, 3–5. [Google Scholar] [CrossRef]
- Pan, X.Y.; Shen, J.Y.; Li, J.Y.; He, B.X.; Hao, G.J. Identification and biological characteristics of the pathogen causing Macrobrachium nipponense soft-shell syndrome. J. Appl. Microbiol. 2009, 36, 1571–1576. [Google Scholar]
- Chen, F.; Sun, J.; Han, Z.; Yang, X.; Xian, J.A.; Lv, A.; Hu, X.; Shi, H. Isolation, Identification and Characteristics of Aeromonas veronii From Diseased Crucian Carp (Carassius auratus gibelio). Front. Microbiol. 2019, 10, 2742. [Google Scholar] [CrossRef] [PubMed]
- Chandrarathna, H.P.S.U.; Chamilani, N.; Dananjaya, S.H.S.; Wijerathne, C.U.B.; Wimalasena, S.H.M.P.; Kwun, H.J.; Heo, G.J.; Jehee, L.; Mahanama, D.Z. Outcome of co-infection with opportunistic and multidrug resistant Aeromonas hydrophila and A. veronii in zebrafish: Identification, characterization, pathogenicity and immune responses. Fish Shellfish Immunol. 2018, 80, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Bücker, R.; Krug, S.M.; Rosenthal, R.; Günzel, D.; Fromm, A.; Zeitz, M.; Chakraborty, T.; Fromm, M.; Epple, H.-J.; Schulzke, J.-D. Aerolysin from Aeromonas hydrophila perturbs tight junction integrity and cell lesion repair in intestinal epithelial HT-29/B6 cells. J. Infect. Dis. 2011, 204, 1283–1292. [Google Scholar] [CrossRef]
- Namba, A.; Mano, N.; Takano, H.; Beppu, T.; Ueda, K.; Hirose, H. OmpA is an adhesion factor of Aeromonas veronii, an optimistic pathogen that habituates in carp intestinal tract. J. Appl. Microbiol. 2008, 105, 1441–1451. [Google Scholar] [CrossRef] [PubMed]
- Lau, T.T.V.; Puah, S.M.; Tan, J.A.M.A.; Puthucheary, S.D.; Chua, K.H. Characterization of the relationship between polar and lateral flagellar genes in clinical Aeromonas dhakensis: Phenotypic, genetic and biochemical analyses. Braz. J. Microbiol. 2021, 52, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Sen, K.; Rodgers, M. Distribution of six virulence factors in Aeromonas species isolated from US drinking water utilities: A PCR identification. J. Appl. Microbiol. 2010, 97, 1077–1086. [Google Scholar] [CrossRef] [PubMed]
- Zhan, F.; Li, Y.; Shi, F.; Lu, Z.; Yang, M.; Li, Q.; Lin, L.; Qin, Z. Transcriptome analysis of Macrobrachium rosenbergii hemocytes reveals in-depth insights into the immune response to Vibrio parahaemolyticus infection. Fish Shellfish Immunol. 2023, 133, 108533. [Google Scholar] [CrossRef] [PubMed]
- Heuzenroeder, M.W.; Wong, C.Y.F.; Flower, R.L.P. Distribution of two hemolytic toxin genes in clinical and environmental isolates of Aeromonas spp.: Correlation with virulence in a suckling mouse model. FEMS Microbiol. Lett. 2010, 174, 131–136. [Google Scholar] [CrossRef]
- Arockiaraj, J.; Gnanam, A.J.; Muthukrishnan, D.; Gudimella, R.; Milton, J.; Singh, A.; Muthupandian, S.; Kasi, M.; Bhassu, S. Crustin, a WAP domain containing antimicrobial peptide from freshwater prawn Macrobrachium rosenbergii: Immune characterization. Fish Shellfish Immunol. 2013, 34, 109–118. [Google Scholar] [CrossRef]
- Kim, M.S.; Jeon, J.M.; Oh, C.W.; Kim, H.W. Molecular characterization of three crustin genes in the morotoge shrimp, Pandalopsis japonica. Comp. Biochem. Phys. B 2012, 163, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Ren, Q. A Kruppel-like factor from Macrobrachium rosenbergii (MrKLF) involved in innate immunity against pathogen infection. Fish Shellfish Immunol. 2019, 95, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Somboonwiwat, K.; Marcos, M.; Tassanakajon, A.; Klinbunga, S.; Aumelas, A.; Romestand, B.; Gueguen, Y.; Boze, H.; Moulin, G.; Bachère, E. Recombinant expression and anti-microbial activity of anti-lipopolysaccharide factor (ALF) from the black tiger shrimp Penaeus monodon. Dev. Comp. Immunol. 2005, 29, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Jiravanichpaisal, P.; Soderhall, I.; Cerenius, L.; Soderhall, K. Antilipopolysaccharide Factor Interferes with White Spot Syndrome Virus Replication In Vitro and In Vivo in the Crayfish Pacifastacus leniusculus. J. Virol. 2006, 80, 10365–10371. [Google Scholar] [CrossRef] [PubMed]
- Li, X.L.; Zhang, Y.F.; Shen, P.J.; Xu, Y.; Gao, Q.; Cheng, H.H.; Wang, W.D.; Gu, Z.M.; Chen, X.F. Baicalin protects giant freshwater prawn, Macrobrachium rosenbergii against Vibrio parahaemolyticus via modulation of the Toll signaling pathway. J. World Aquac. Soc. 2021, 52, 987–1000. [Google Scholar] [CrossRef]
- Zheng, J.B.; Mao, Y.; Su, Y.Q.; Wang, J. Identification and functional characterization of a novel C-type lectin from the kuruma shrimp, Marsupenaeus japonicus. Biochem. Biophys. Res. Commun. 2020, 530, 547–553. [Google Scholar] [CrossRef]
- Li, Y.B.; Pan, L.Q.; Yu, J.H. The injection of one recombinant C-type lectin (LvLec) induced the immune response of hemocytes in Litopenaeus vannamei. Fish Shellfish Immun. 2022, 124, 324–331. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequence (5′–3′) | Product Size (bp) | Reference |
|---|---|---|---|
| aer | CCTATGGCCTGAGCGAGAAG | 417 | MH332385.1 |
| CCAGTTCCAGTCCCACCACT | |||
| ompA | GCGGTTTATCGCTTTGGT | 397 | CP032839.1 |
| CACGCTTGGACTTGCTGA | |||
| flgA | GGGACCTGCTGAGTGAAA | 326 | CP032839.1 |
| GACCGATACGGCACCTAC | |||
| flgM | GCTACTGTCAAGCTGGACTC | 194 | CP032839.1 |
| AGATTGGCCTCGAAACTG | |||
| flgN | AGTTGCTTGCTGCGATAGA | 181 | CP032839.1 |
| AGACGACGGTTTGAGACG | |||
| lip | GTGCCGTCTGCCTTGGTGA | 598 | CP032839.1 |
| CCCGTCTATTGCGGGTTCGA | |||
| hlyA | CGGACGATTATCAGGATGG | 289 | KY624579.1 |
| CAAGAACGAGTTTCAGTGGC | |||
| tapA | ATGACCTCTAGCCCCAATA | 550 | CP050851.1 |
| ACCCGATTGATTTCTGCC |
| Target Gene | Primer Sequence (5′–3′) | Product Size (bp) |
|---|---|---|
| ALF1 | GCCGATGGTGTCCTGGATG | 157 |
| TCCATGCGTCGTCCTCCG | ||
| ALF2 | GGCACCAAACTCACTGGA | 163 |
| CTTAGCACATGCGACCCTG | ||
| ALF3 | GAACTGCTGTCCAACCCTG | 232 |
| CCGGATGCTCCTCCGTTATC | ||
| Crustin | TGAAACTAACCTGTTCCAACG | 165 |
| GAATGCCCTGCGATCCGAAGAA | ||
| C-lectin | TGAAATTGCCTGTTGTTATG | 180 |
| GGAGGGTGAGATGTAGCC | ||
| Lysozyme | GACCTTGCGTCATGCCAGAT | 182 |
| CCATGGGTTTATGTGCGTCTTC | ||
| 18S rRNA | TATACGCTAGTGGAGCTGGAA | 182 |
| GGGGAGGTAGTGACGAAAAAT |
| Group | Infected Amount | Density (CFU/mL) | Dead Prawns after Challenge | Mortality | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 12 h | 1 d | 2 d | 3 d | 4 d | 5 d | 6 d | 7 d | ||||
| Test | 30 | 1.8 × 108 | 18 | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 100.00% |
| 30 | 1.8 × 107 | 7 | 13 | 19 | 25 | 27 | 28 | 29 | 29 | 96.67% | |
| 30 | 1.8 × 106 | 0 | 3 | 6 | 10 | 13 | 15 | 16 | 17 | 56.67% | |
| 30 | 1.8 × 105 | 0 | 0 | 2 | 3 | 5 | 5 | 5 | 6 | 16.67% | |
| 30 | 1.8 × 104 | 0 | 0 | 0 | 2 | 3 | 4 | 4 | 4 | 13.33% | |
| Control | 30 | PBS | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Biochemical Test | JDM1-1 | A. veronii a | A. sobria a | A. hydrophila a |
|---|---|---|---|---|
| Oxidase | + | + | + | + |
| Motility | + | + | + | + |
| V-P test | + | + | d | + |
| O-F test | F | F | F | F |
| Lactose | - | - | - | d |
| Maltose | + | + | + | + |
| Mannitol | + | + | + | + |
| Mannose | + | + | + | [+] |
| Sorbitol | - | - | d | - |
| Inositol | - | - | - | - |
| Salicin | + | + | - | + |
| H2S production | - | - | - | + |
| α-methyl-D-glucoside | + | [+] | d | d |
| Tartrate utilization | + | [+] | d | - |
| Nitrate reduction | + | + | + | + |
| Citrate utilization | + | + | - | - |
| Mucate utilization | − | − | - | - |
| Malonate utilization | − | − | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.; Chen, Z.; Zhang, Z.; Qian, Q.; Chen, A.; Qin, L.; Tang, X.; Jiang, Q.; Zhang, X. Pathogenicity of Aeromonas veronii Isolated from Diseased Macrobrachium rosenbergii and Host Immune-Related Gene Expression Profiles. Microorganisms 2024, 12, 694. https://doi.org/10.3390/microorganisms12040694
Gao X, Chen Z, Zhang Z, Qian Q, Chen A, Qin L, Tang X, Jiang Q, Zhang X. Pathogenicity of Aeromonas veronii Isolated from Diseased Macrobrachium rosenbergii and Host Immune-Related Gene Expression Profiles. Microorganisms. 2024; 12(4):694. https://doi.org/10.3390/microorganisms12040694
Chicago/Turabian StyleGao, Xiaojian, Zhen Chen, Zirui Zhang, Qieqi Qian, Anting Chen, Lijie Qin, Xinzhe Tang, Qun Jiang, and Xiaojun Zhang. 2024. "Pathogenicity of Aeromonas veronii Isolated from Diseased Macrobrachium rosenbergii and Host Immune-Related Gene Expression Profiles" Microorganisms 12, no. 4: 694. https://doi.org/10.3390/microorganisms12040694
APA StyleGao, X., Chen, Z., Zhang, Z., Qian, Q., Chen, A., Qin, L., Tang, X., Jiang, Q., & Zhang, X. (2024). Pathogenicity of Aeromonas veronii Isolated from Diseased Macrobrachium rosenbergii and Host Immune-Related Gene Expression Profiles. Microorganisms, 12(4), 694. https://doi.org/10.3390/microorganisms12040694