
Association of Acidotolerant Cyanobacteria to Microbial Mats below pH 1 in Acidic Mineral Precipitates in Río Tinto River in Spain

 ,
,  ,
,  , ,
, , 
 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. pH Measurement
2.3. Irradiance Records
2.4. X-ray Diffraction (XRD) Analysis
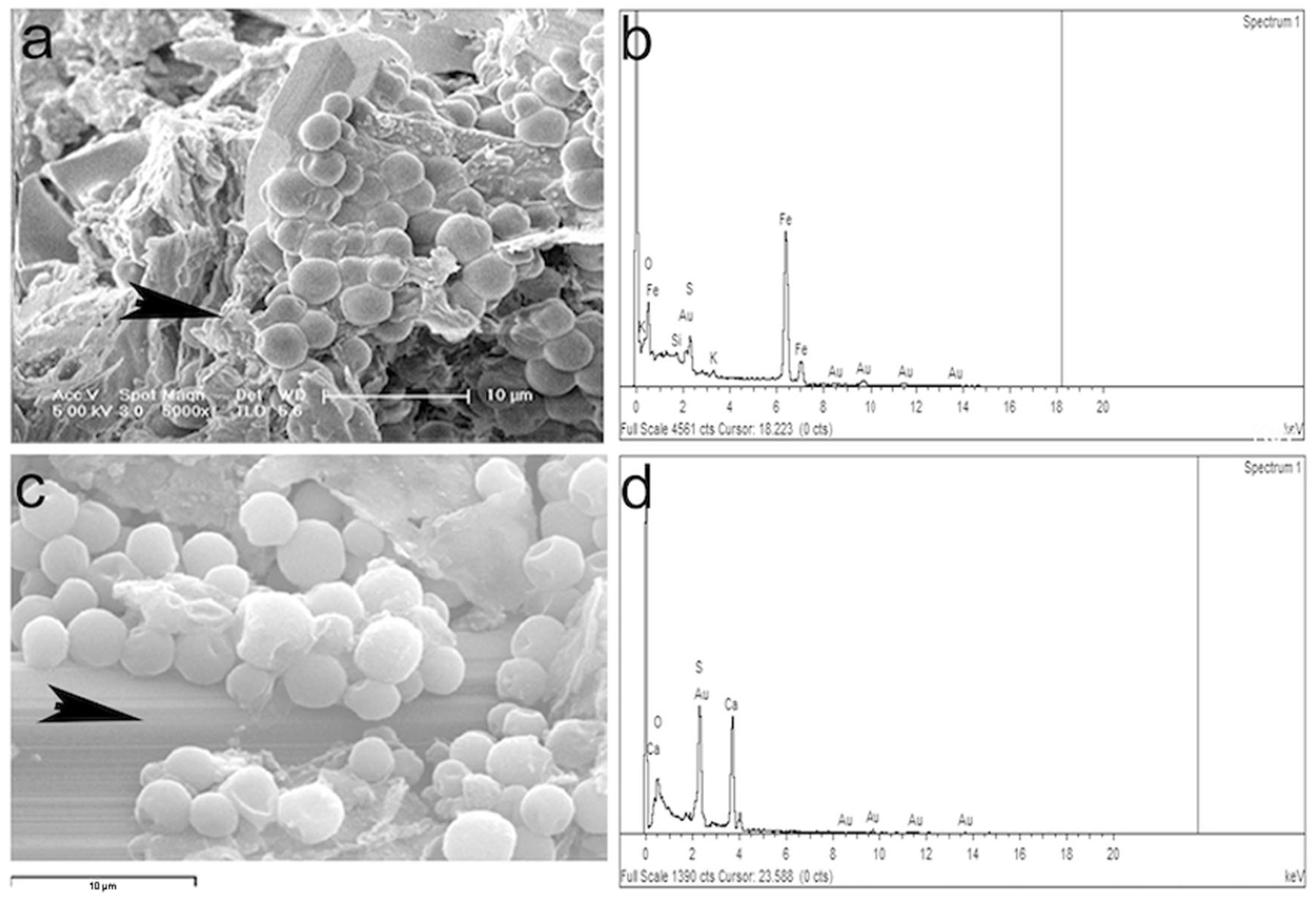
2.5. Scanning Electron Microscopy (SEM)
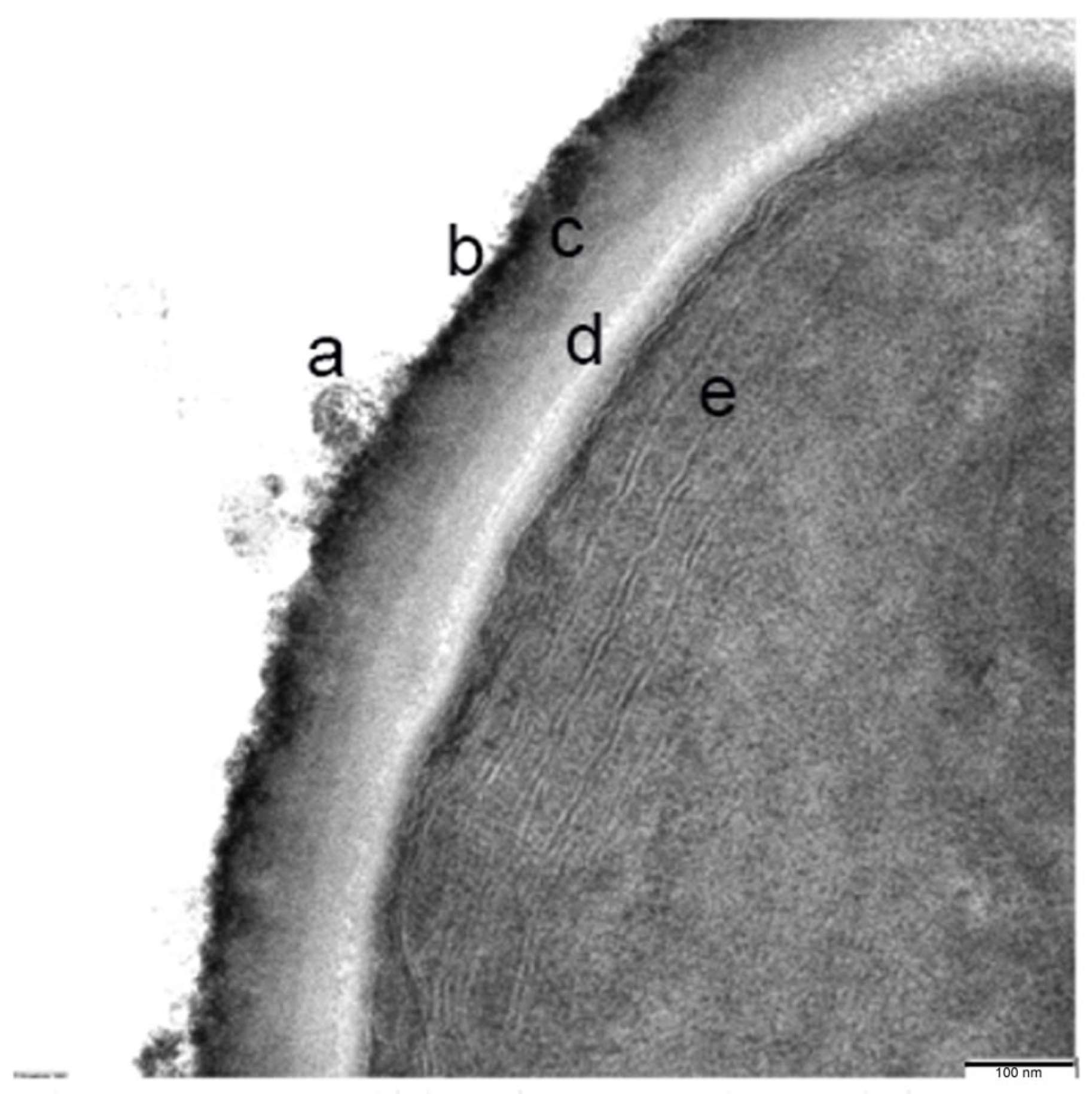
2.6. Transmission Electron Microscopy (TEM) and Immunocytochemistry
2.7. Fluorescence “In Situ” Hybridization Combined with Catalyzed Reporter Deposition (CARD-FISH)
2.8. Metagenomics Shotgun Sequencing and Analysis
2.9. Spectroscopy Measurements Setup
3. Results
3.1. Physicochemical Characterization
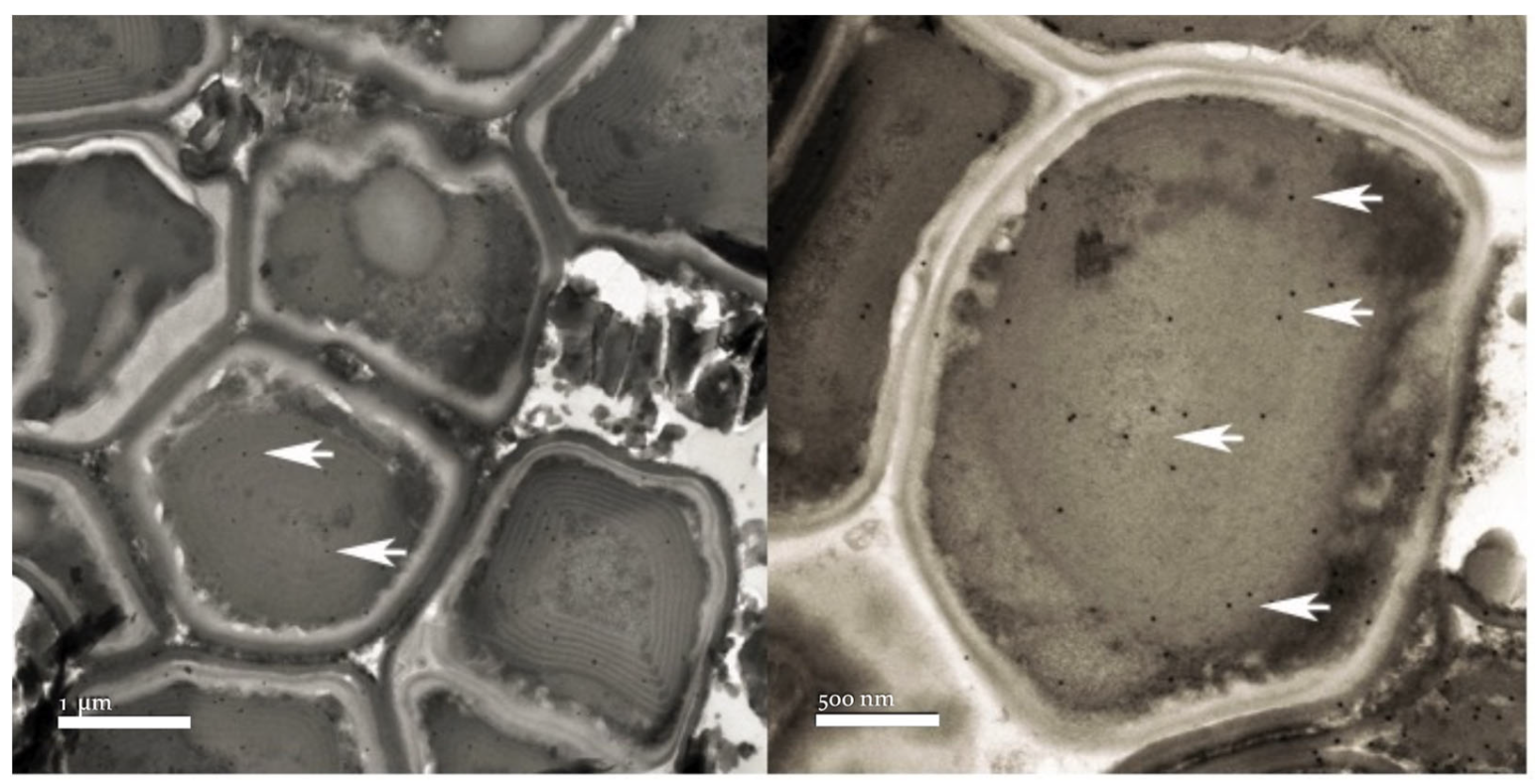
3.2. Microscopy Observations
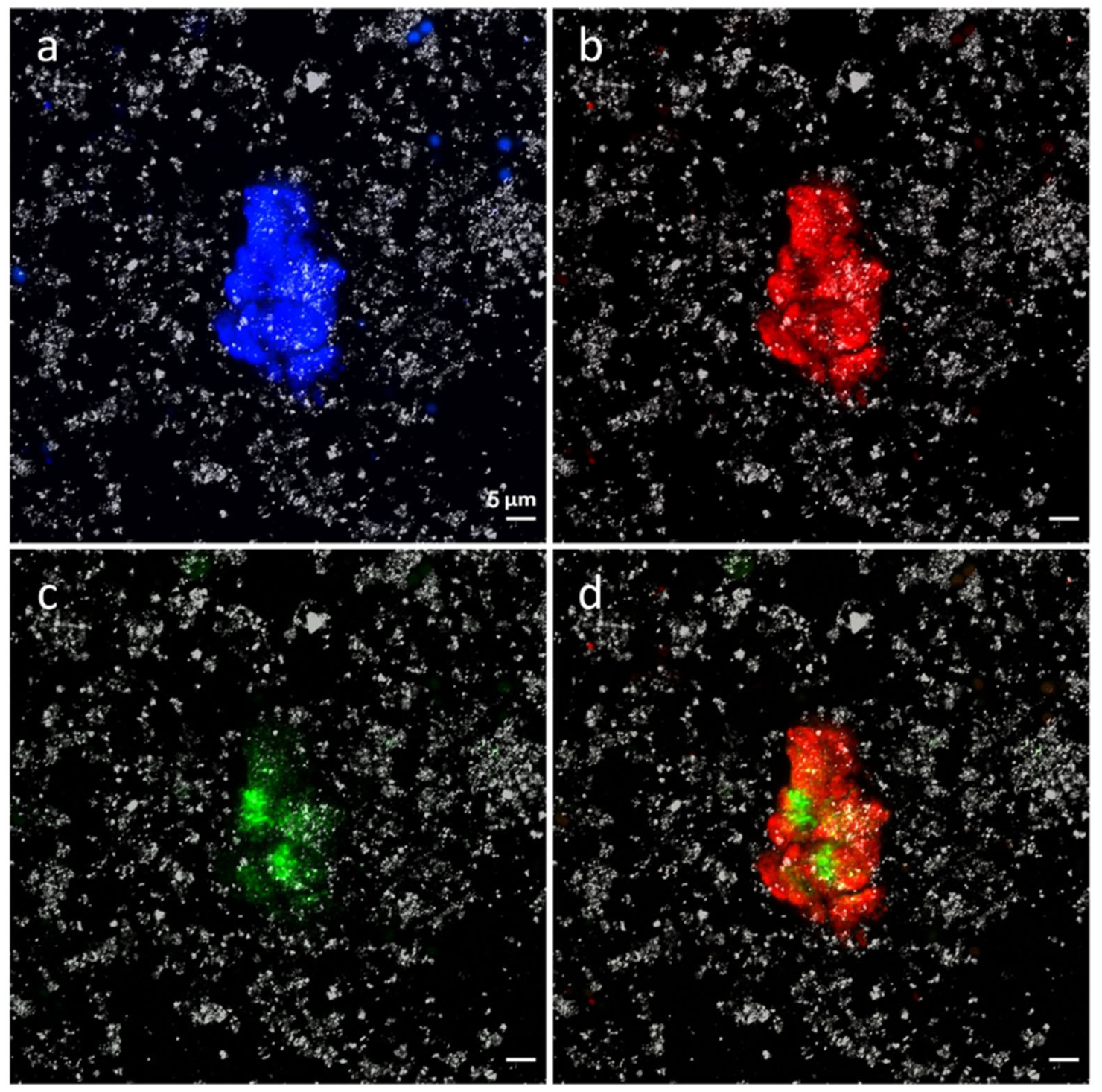
3.3. Immunological and Fluorescence In Situ Hybridization Analysis
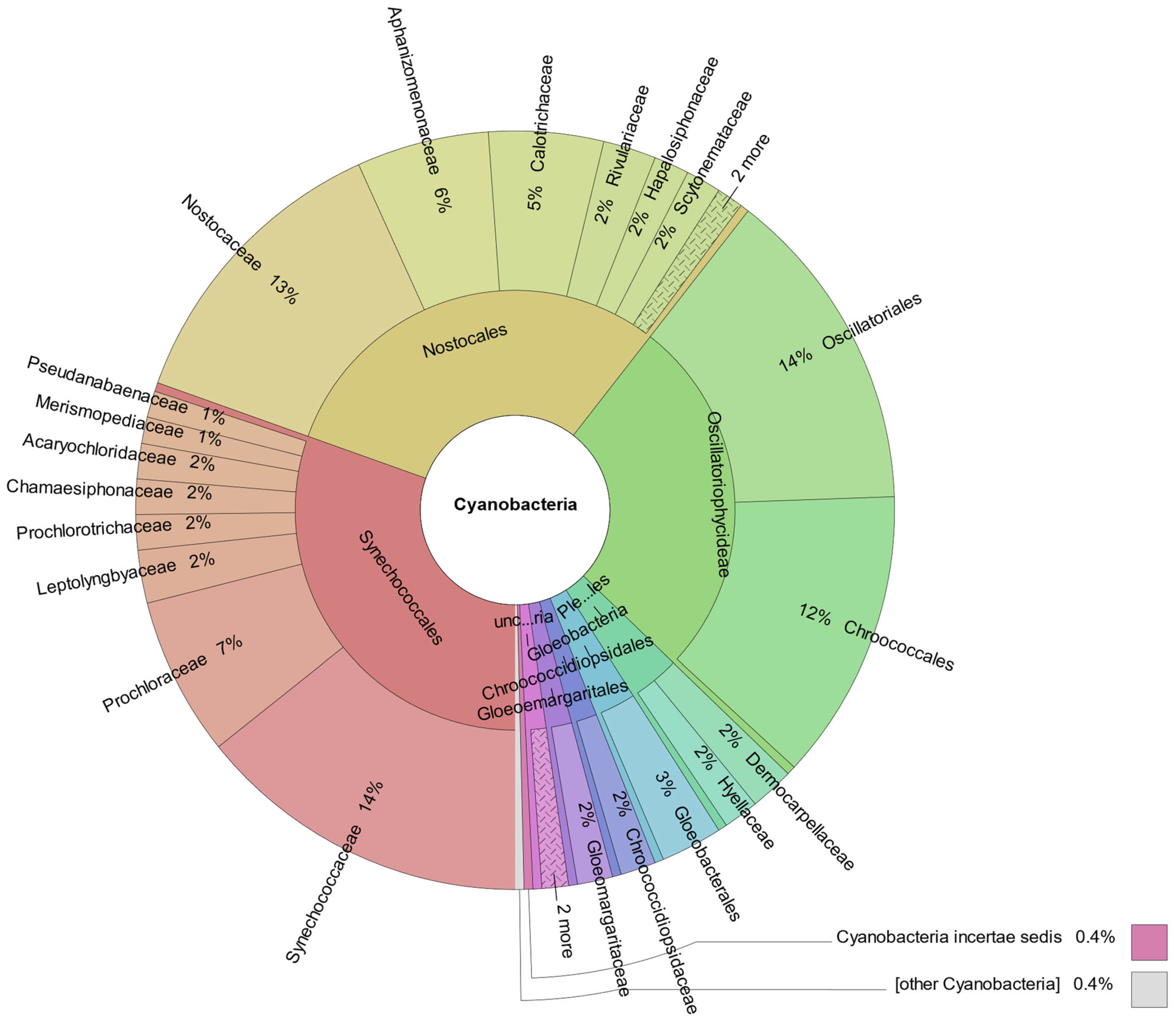
3.4. Metagenomic Analysis
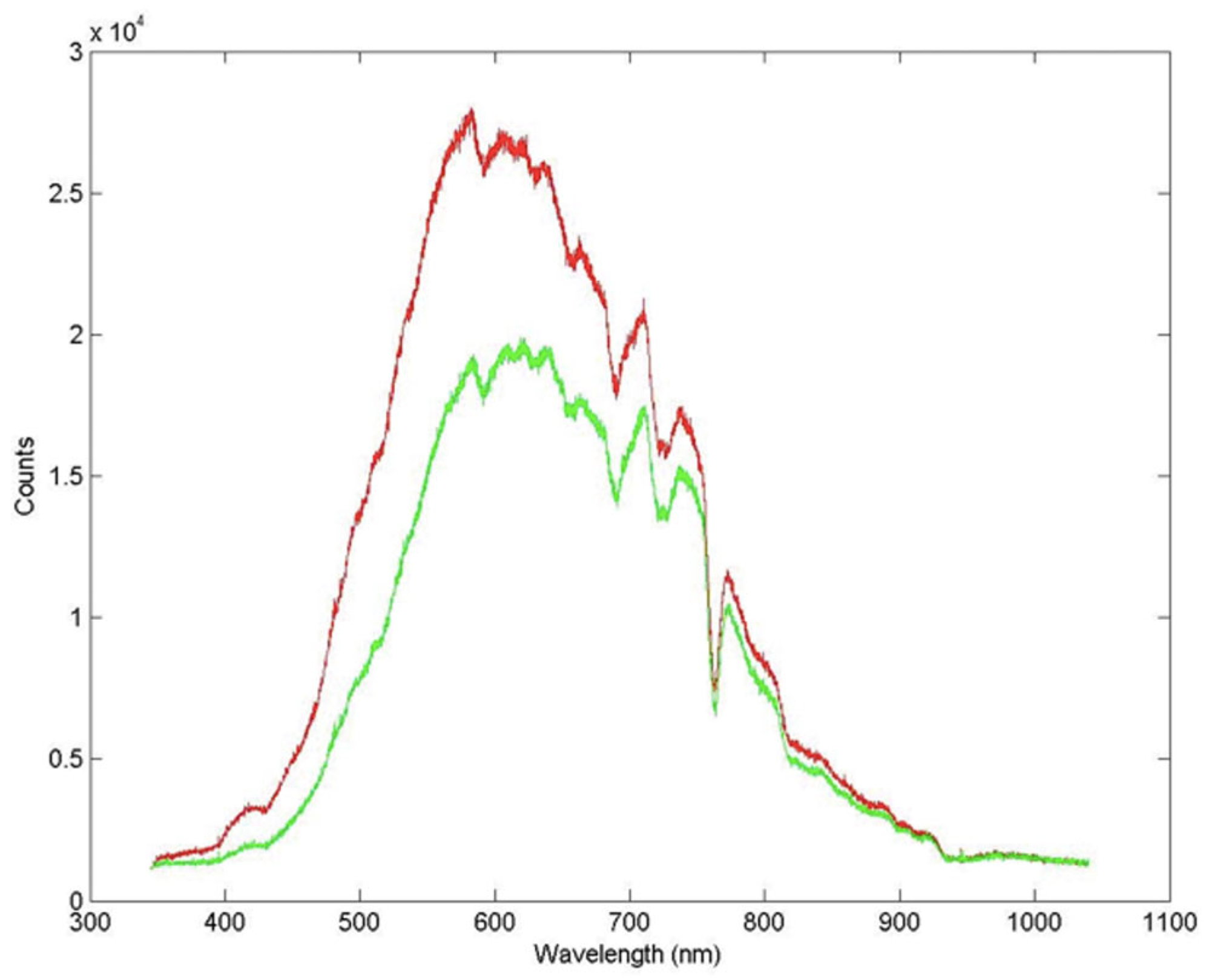
3.5. VIS-NIR and Micro-Raman spectroscopy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oren, A. A proposal for further integration of the cyanobacteria under the Bacteriological Code. Int. J. Syst. Evol. Microbiol. 2004, 54 Pt 5, 1895–1902. [Google Scholar] [CrossRef] [PubMed]
- De los Ríos, A.; Grube, M.; Sancho, L.G.; Ascaso, C. Ultrastructural and genetic characteristics of endolithic cyanobacterial biofilms colonizing Antarctic granite rocks. FEMS Microbiol. Ecol. 2007, 59, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Whitton, B.A.; Potts, M. (Eds.) Ecology of Cyanobacteria II: Their Diversity in Space and Time; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Keshari, N.; Zhao, Y.; Das, S.K.; Zhu, T.; Lu, X. Cyanobacterial Community Structure and Isolates from Representative Hot Springs Yunnan Province, China Using an Integrative Approach. Front. Microbiol. 2022, 13, 872598. [Google Scholar] [CrossRef] [PubMed]
- Alcorta, J.; Alarcón-Schumacher, T.; Salgado, O.; Díez, B. Taxonomic Novelty and Distinctive Genomic Features of Hot Spring Cyanobacteria. Front. Genet. 2020, 11, 568223. [Google Scholar] [CrossRef] [PubMed]
- Kaštovský, J.; Johansen, J.R.; Hauerová, R.; Akagha, M.U. Hot Is Rich—An Enormous Diversity of Simple Trichal Cyanobacteria from Yellowstone Hot Springs. Diversity 2023, 15, 975. [Google Scholar] [CrossRef]
- Dominic, T.K.; Madhusoodanan, P.V. Cyanobacteria from extreme acidic environments. Curr. Sci. 1999, 77, 1021–1023. [Google Scholar]
- Mesa, V.; Gallego, J.L.; González-Gil, R.; Lauga, B.; Sánchez, J.; Méndez-García, C.; Peláez, A.I. Bacterial, Archaeal, and Eukaryotic Diversity across Distinct Microhabitats in an Acid Mine Drainage. Front. Microbiol. 2017, 8, 1756. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, C.E.W.; Schäfer, H.; Beisker, W. Do acid-tolerant Cyanobacteria exist? Clean Soil Air Water 1998, 26, 13–19. [Google Scholar] [CrossRef]
- Lessmann, D.; Fyson, A.; Nixdorf, B. Phytoplankton of the extremely acidic mining lakes of Lusatia (Germany) with pH ≤ 3. Hydrobiologia 2000, 433, 123–128. [Google Scholar] [CrossRef]
- Korzhenkov, A.A.; Toshchakov, S.V.; Bargiela, R.; Gibbard, H.; Ferrer, M.; Teplyuk, A.V.; Jones, D.L.; Kublanov, I.V.; Golyshin, P.N.; Golyshina, O.V. Archaea dominate the microbial community in an ecosystem with low-to-moderate temperature and extreme acidity. Microbiome 2019, 7, 11. [Google Scholar] [CrossRef]
- Hirooka, S.; Hirose, Y.; Kanesaki, Y.; Higuchi, S.; Fujiwara, T.; Onuma, R.; Era, A.; Ohbayashi, R.; Uzuka, A.; Nozaki, H.; et al. Acidophilic green algal genome provides insights into adaptation to an acidic environment. Proc. Natl. Acad. Sci. USA 2017, 114, E8304–E8313. [Google Scholar] [CrossRef] [PubMed]
- Kallas, T.; Castenholz, R.W. Internal pH and ATP-ADP pools in the Cyanobacterium Synechococcus sp. during exposure to growth inhibiting los pH. J. Bacteriol. 1982, 149, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Poza-Carrión, C.; Fernández-Valiente, E.; Fernández-Piñas, F.; Leganés, F. Acclimation of photosynthetic pigments and photosynthesis of cyanobacterium Nostoc sp. strain UAM206 to combined fluctuations of irradiance, pH, and inorganic carbón availability. J. Plant Physiol. 2001, 158, 1455–1461. [Google Scholar] [CrossRef]
- Kallas, T.; Castenholz, R.W. Rapid transient growth at low pH in the cyanobacterium Synechococcus sp. J. Bacteriol. 1982, 149, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Mangan, N.M.; Flamholz, A.F.; Hood, R.D.; Milo, R.; Savage, D.F. pH determines the energetic efficiency of the cyanobacterial CO2 concentrating mechanism. Proc. Natl. Acad. Sci. USA 2016, 113, E5354–E5362. [Google Scholar] [CrossRef] [PubMed]
- Amils, R.; González-Toril, E.; Fernández-Remolar, D.; Gómez, F.; Rodríguez, N.; Durán, C. Interaction of the sulfur and iron cycles in the Tinto River ecosystem. Rev. Environ. Sci. Biotechnol. 2002, 1, 299–309. [Google Scholar] [CrossRef]
- Davis, R., Jr.; Welty, A.; Borrego, J.; Morales, J.; Pendon, J.; Ryan, J.G. Rio Tinto estuary (Spain): 5000 years of pollution. Environ. Geol. 2000, 39, 1107–1116. [Google Scholar] [CrossRef]
- Nieto, J.M.; Sarmiento, A.M.; Olías, M.; Canovas, C.R.; Riba, I.; Kalman, J.; Delvalls, T.A. Acid mine drainage pollution in the Tinto and Odiel rivers (Iberian Pyrite Belt, SW Spain) and bioavailability of the transported metals to the Huelva Estuary. Environ. Int. 2007, 33, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, O.; Nieto, J.M. Background conditions and mining pollution throughout history in the Río Tinto. (SW Spain). Environmernts 2015, 2, 295–316. [Google Scholar]
- Fernández-Remolar, D.; Rodríguez, N.; Gómez, F.; Amils, R. Geological record of an acidic environment driven by iron hydrochemistry: The Tinto River system. J. Geophys. Res. 2003, 108, 5080–5095. [Google Scholar] [CrossRef]
- Gómez-Ortiz, D.; Fernández-Remolar, D.C.; Granda, Á.; Quesada, C.; Granda, T.; Prieto-Ballesteros, O.; Molina, A.; Amils, R. Identification of the subsurface sulfide bodies responsible for acidity in Rio Tinto source water, Spain. Earth Planet. Sci. Lett. 2014, 391, 36–41. [Google Scholar] [CrossRef]
- Amils, R.; Fernández-Remolar, D.C.; The IPBSL Team. Río Tinto: A Geochemical and Mineralogical Terrestrial Analogue of Mars. Life 2014, 4, 511–534. [Google Scholar] [CrossRef] [PubMed]
- Amils, R.; Escudero, C.; Oggerin, M.; Puente Sanchez, F.; Arce Rodríguez, A.; Fernandez Remolar, D.; Rodríguez, N.; García Villadangos, M.; Sanz, J.L.; Briones, C.; et al. Coupled C, H, N, S and Fe biogeochemical cycles operating in the continental deep subsurface of the Iberian Pyrite Belt. Environ. Microbiol. 2023, 25, 428–453. [Google Scholar] [CrossRef] [PubMed]
- López-Archilla, A.I.; Marín, I.; Amils, R. Microbial community composition and ecology of an acidic aquatic environment: The Tinto River, Spain. Microb. Ecol. 2001, 41, 20–35. [Google Scholar] [CrossRef] [PubMed]
- Amaral-Zettler, L.A.; Gómez, F.; Zettler, E.; Keenan, B.G.; Amils, R. Microbiology: Eukaryotic diversity in Spain’s River of Fire. Nature 2002, 417, 137. [Google Scholar] [CrossRef] [PubMed]
- Jansson, C.; Northen, T. Calcifying cyanobacteria—The potential of biomineralization for carbon capture and storage. Curr. Opin. Biotechnol. 2010, 21, 365–371. [Google Scholar] [CrossRef]
- Reynolds, E.S. The use of lead citrate at high pH as an electron opaque stain in electron microscopy. J. Cell Biol. 1963, 17, 208. [Google Scholar] [CrossRef] [PubMed]
- Escudero, C.; Vera, M.; Oggerin, M.; Amils, R. Active microbial biofilms in deep poor porous continental subsurface rocks. Sci. Rep. 2018, 8, 1538. [Google Scholar] [CrossRef]
- Amann, R.I.; Binder, B.J.; Olson, R.J.; Chisholm, S.W.; Devereux, R.; Stahl, D.A. Combination of 16S rRNA-targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial populations. App. Environ. Microbiol. 1990, 56, 1919–1925. [Google Scholar] [CrossRef]
- Schönhuber, W.; Zarda, B.; Eix, S.; Rippka, R.; Herdman, M.; Ludwig, W.; Amann, R. In situ identification of cyanobacteria with horseradish peroxidase-labeled, rRNA-targeted oligonucleotide probes. Appl. Environ. Microbiol. 1999, 65, 1259–1267. [Google Scholar] [CrossRef]
- Daims, H.; Brühl, A.; Amann, R.; Schleifer, K.-H.; Wagner, M. The Domain-specific Probe EUB338 is Insufficient for the Detection of all Bacteria: Development and Evaluation of a more Comprehensive Probe Set. Syst. Appl. Microbiol. 1999, 22, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Meth. 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Uritskiy, G.V.; DiRuggiero, J.; Taylor, J. MetaWRAP—A flexible pipeline for genome-resolved metagenomic data analysis. Microbiome 2018, 6, 158. [Google Scholar] [CrossRef] [PubMed]
- Nurk, S.; Meleshko, D.; Korobeynikov, A.; Pevzner, P.A. metaSPAdes: A new versatile metagenomic assembler. Genome Res. 2017, 27, 824–834. [Google Scholar] [CrossRef] [PubMed]
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef] [PubMed]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive metagenomic visualization in a Web browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef] [PubMed]
- Hyatt, D.; Chen, G.L.; LoCascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and Sensitive Protein Alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Gantt, E.; Conti, S.F. Ultrastructure of blue-green algae. J. Bacteriol. 1969, 97, 1486–1493. [Google Scholar] [CrossRef]
- Beversdorf, L.J.; Miller, T.R.; McMahon, K.D. The Role of Nitrogen Fixation in Cyanobacterial Bloom Toxicity in a Temperate, Eutrophic Lake. PLoS ONE 2013, 8, e56103. [Google Scholar] [CrossRef]
- Kim, J.; Rees, D.C. Nitrogenase and biological nitrogen fixation. Biochemistry 1994, 33, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Frías, J.E.; Mérida, A.; Herrero, A.; Martín-Nieto, J.; Flores, E. General distribution of the Nitrogen control gene ntcA in Cyanobacteria. J. Bacteriol. 1993, 175, 5710–5713. [Google Scholar] [CrossRef]
- López-Maury, L.; Heredia-Martínez, L.G.; Florencio, F.J. Characterization of TrxC, an Atypical Thioredoxin Exclusively Present in Cyanobacteria. Antioxidants 2018, 7, 164. [Google Scholar] [CrossRef] [PubMed]
- Hannemann, L.; Suppanz, I.; Ba, Q.; MacInnes, K.; Drepper, F.; Warscheid, B.; Koch, H.G. Redox Activation of the Universally Conserved ATPase YchF by Thioredoxin 1. Antioxid. Redox Signal. 2016, 24, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Holmgren, A. The thioredoxin antioxidant system. Free Radic. Biol. Med. 2014, 66, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Holian, O.; Kumar, R.; Attar, R. Apoprotein A-l is a cofactor independent substrate of protein kinase c. Biochem. Biophys. Res. Commun. 1991, 179, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Petit, M.A.; Ehrlich, D. Essential bacterial helicases that counteract the toxicity of recombination proteins. EMBO J. 2002, 21, 3137–3147. [Google Scholar] [CrossRef] [PubMed]
- Yeom, J.; Lee, Y.; Park, W. ATP-dependent RecG Helicase is required for the transcriptional regulator OxyR Function in Pseudomonas species. J. Biol. Chem. 2012, 287, 24492–24504. [Google Scholar] [CrossRef] [PubMed]
- Cloutis, E.A.; Hawthorne, F.C.; Mertzman, S.A.; Krenn, K.; Craig, M.A.; Marcino, D.; Methot, M.; Strong, J.; Mustard, J.F.; Blaney, D.L.; et al. Detection and discrimination of sulfate minerals using reflectance spectroscopy. Icarus 2006, 184, 121–157. [Google Scholar] [CrossRef]
- Kutser, T. Quantitative detection of chlorophyll in cyanobacterial blooms by satellite remote sensing. Limnol. Oceanogr. 2004, 49, 2179–2189. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Schalles, J.F.; Rundquist, D.C.; Schiebe, F.R.; Yacobi, Y.Z. Comparative reflectance properties of algal cultures with manipulated densities. J. Appl. Phycol. 1999, 11, 345–354. [Google Scholar] [CrossRef]
- Němečková, K.; Culka, A.; Jehlička, J. Detecting pigments from gypsum endoliths using Raman spectroscopy: From field prospection to laboratory studies. J. Raman Spectrosc. 2021, 53, 630–644. [Google Scholar] [CrossRef]
- Jehlička, J.; Edwards, H.G.; Osterrothová, K.; Novotná, J.; Nedbalová, L.; Kopecký, J.; Němec, I.; Oren, A. Potential and limits of Raman spectroscopy for carotenoid detection in microorganisms: Implications for astrobiology. Philos. Trans. R. Soc. A 2014, 372, 20140199. [Google Scholar] [CrossRef]
- Margulis, L.; Mazur, P.; Banghoom, E.S.; Halvorson, H.O.; Jukes, T.H.S.; Kaplan, I.R. The Viking Mission implications for life in the Vallis Marineris area. Science 1979, 305, 78–81. [Google Scholar]
- Elwood Madden, M.; Bodnar, R.; Rimstidt, J. Jarosite as an indicator of water-limited chemical weathering on Mars. Nature 2004, 431, 321–823. [Google Scholar] [CrossRef]
- Fairén, A.G.; Schulze-Makuch, D.; Rodríguez, A.P.; Fink, W.; Davila, A.F.; Uceda, E.R.; Furfaro, R.; Amils, R.; McKay, C.P. Evidence for Amazonian acidic liquid water on Mars—A reinterpretation of MER mission results. Planet. Space Sci. 2009, 57, 276–287. [Google Scholar] [CrossRef]
- Andrews-Hanna, J.C.; Zuber, M.T.; Arvidson, R.E.; Wiseman, S.M. Early Mars hydrology: Meridiani playa depositsand the sedimentary record of Arabia Terra. J. Geophys. Res. 2010, 115, E06002. [Google Scholar] [CrossRef]
- Bižić, M.; Klintzsch, T.; Ionescu, D.; Hindiyeh, M.Y.; Günthel, M.; Muro-Pastor, A.M.; Eckert, W.; Urich, T.; Keppler, F.; Grossart, H.P. Aquatic and terrestrial cyanobacteria produce methane. Sci. Adv. 2020, 6, eaax5343. [Google Scholar] [CrossRef]
- Hanson, R.S.; Hanson, T.E. Methanotrophic bacteria. Microbiol. Rev. 1996, 60, 439–471. [Google Scholar] [CrossRef]
- Nangle, S.N.; Wolfson, M.Y.; Hartsough, L.; Ma, N.J.; Mason, C.E.; Merighi, M.; Nathan, V.; Silver, P.A.; Simon, M.; Swett, J.; et al. The case of biotech on Mars. Nat. Biotech. 2020, 38, 401–407. [Google Scholar] [CrossRef]
- Fernández-Remolar, D.; Morris, R.V.; Gruener, J.E.; Amils, R.; Knoll, A.H. The Rio Tinto Basin, Spain: Mineralogy, Sedimentary Geobiology, and implications for interpretation of outcrop rocks at Meridiani Planum, Mars. Earth Planet. Sci. Lett. 2005, 240, 149–167. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez, F.; Rodríguez, N.; Rodríguez-Manfredi, J.A.; Escudero, C.; Carrasco-Ropero, I.; Martínez, J.M.; Ferrari, M.; De Angelis, S.; Frigeri, A.; Fernández-Sampedro, M.; et al. Association of Acidotolerant Cyanobacteria to Microbial Mats below pH 1 in Acidic Mineral Precipitates in Río Tinto River in Spain. Microorganisms 2024, 12, 829. https://doi.org/10.3390/microorganisms12040829
Gómez F, Rodríguez N, Rodríguez-Manfredi JA, Escudero C, Carrasco-Ropero I, Martínez JM, Ferrari M, De Angelis S, Frigeri A, Fernández-Sampedro M, et al. Association of Acidotolerant Cyanobacteria to Microbial Mats below pH 1 in Acidic Mineral Precipitates in Río Tinto River in Spain. Microorganisms. 2024; 12(4):829. https://doi.org/10.3390/microorganisms12040829
Chicago/Turabian StyleGómez, Felipe, Nuria Rodríguez, José Antonio Rodríguez-Manfredi, Cristina Escudero, Ignacio Carrasco-Ropero, José M. Martínez, Marco Ferrari, Simone De Angelis, Alessandro Frigeri, Maite Fernández-Sampedro, and et al. 2024. "Association of Acidotolerant Cyanobacteria to Microbial Mats below pH 1 in Acidic Mineral Precipitates in Río Tinto River in Spain" Microorganisms 12, no. 4: 829. https://doi.org/10.3390/microorganisms12040829
APA StyleGómez, F., Rodríguez, N., Rodríguez-Manfredi, J. A., Escudero, C., Carrasco-Ropero, I., Martínez, J. M., Ferrari, M., De Angelis, S., Frigeri, A., Fernández-Sampedro, M., & Amils, R. (2024). Association of Acidotolerant Cyanobacteria to Microbial Mats below pH 1 in Acidic Mineral Precipitates in Río Tinto River in Spain. Microorganisms, 12(4), 829. https://doi.org/10.3390/microorganisms12040829