
Immune Response to Coccidioidomycosis and the Development of a Vaccine


{kind=link}
Abstract
:1. Coccidioidomycosis and Coccidioides
2. Public Health Impact of Coccidioidomycosis
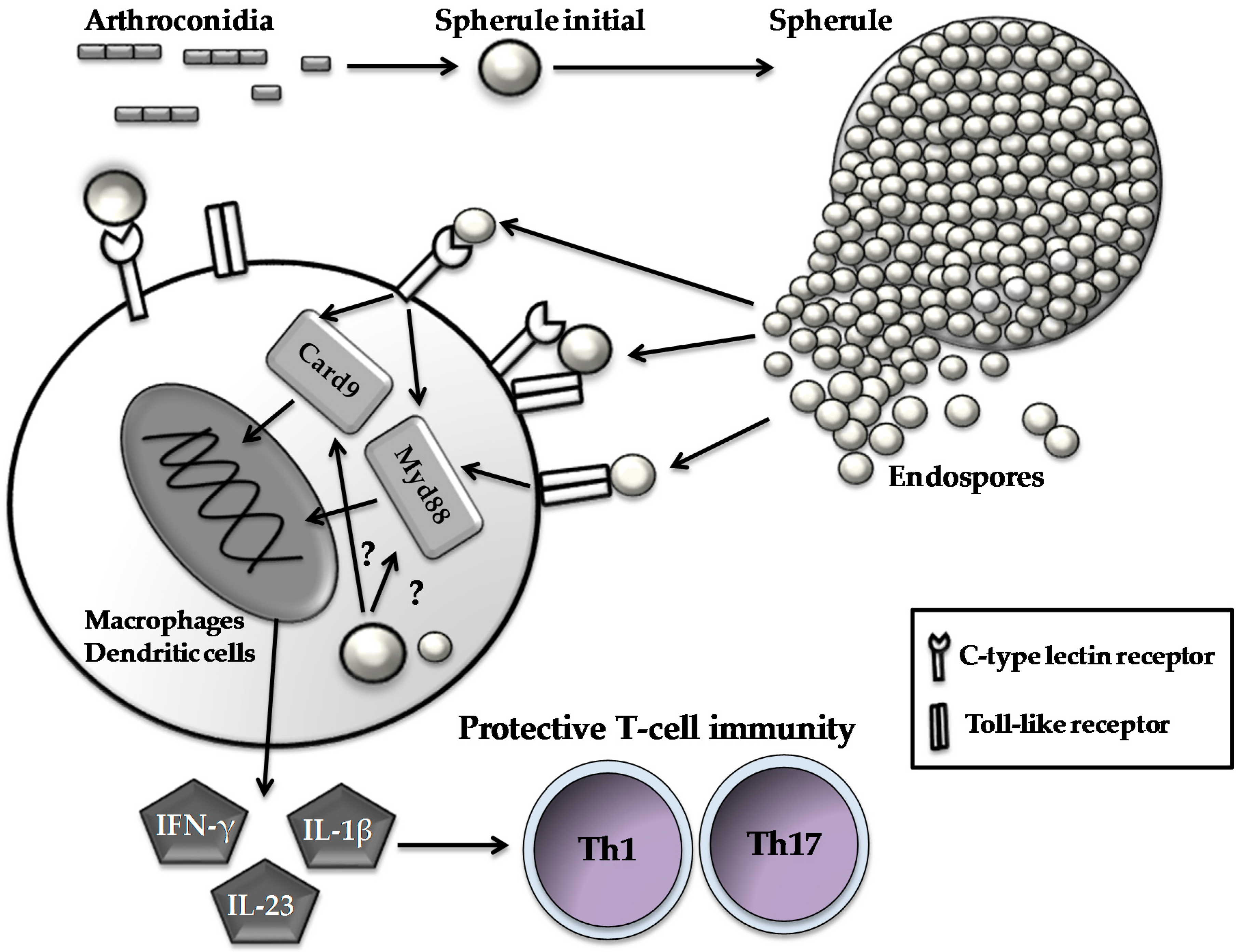
3. Responses of Innate Immune Cells to Coccidioides Infection
4. Recognition of Coccidioides by Pattern Recognition Receptors (PRRs)
5. Adaptive T-Cell Responses to Coccidioides Infection
6. Development of a Multivalent Vaccine against Coccidioides Infection
7. Murine Models for Evaluation of Immune Responses to Coccidioides Infection
8. Conclusions
Acknowledgments
Conflicts of Interest
Abbreviations
| Amn1 | α-1,2-mannosidase |
| Ag2/Pra | proline-rich coccidioidal antigen |
| Card9 | caspase recruitment domain-containing protein 9 |
| CLRs | C-type lectin receptors |
| CTLDs | C-type lectin-like domains |
| CpG | 5′-cytosine-phosphate-guanine-3′ oligonucleotide |
| Csa | Coccidioides specific antigen |
| DCs | dendritic cells |
| FKS | formalin killed spherule |
| GP | β-glucan particles |
| HLA-DR4 | human leukocyte antigen-DR Related |
| IFA | incomplete Freud’s adjuvant |
| iNOS | inducible nitric oxide synthase |
| ITAM | immune receptor tyrosine-based activation motif |
| MHC | major histocompatibility complex |
| Mincle | macrophage-inducible C-type lectin |
| MyD88 | myeloid differentiation primary response protein 88 |
| NODs | Nod-like receptors |
| NO | Nitric oxide |
| NOX2 | nicotinamide adenine dinucleotide phosphate oxidase subunit gp91 (phox) |
| PAMPs | pathogen-associated molecular patterns |
| Pep1 | aspartyl protease 1 |
| Plb | phospholipase B |
| PBMCs | peripheral blood monocytic cells |
| PRRs | pattern recognition receptors |
| RNS | reactive nitrogen species |
| ROS | reactive oxygen species |
| SOWgp | spherule outer wall glycoprotein |
| STAT1 | signal transducer and activator of transcription 1 |
| Tc | cytotoxic T-cells |
| Th | T helper cells |
| TLRs | Toll-like receptors |
References
- Engelthaler, D.M.; Roe, C.C.; Hepp, C.M.; Teixeira, M.; Driebe, E.M.; Schupp, J.M.; Gade, L.; Waddell, V.; Komatsu, K.; Arathoon, E.; et al. Local population structure and patterns of western hemisphere dispersal for Coccidioides spp., the fungal cause of Valley fever. MBio 2016, 7, e00550-16. [Google Scholar] [CrossRef] [PubMed]
- Litvintseva, A.P.; Marsden-Haug, N.; Hurst, S.; Hill, H.; Gade, L.; Driebe, E.M.; Ralston, C.; Roe, C.; Barker, B.M.; Goldoft, M.; et al. Valley fever: Finding new places for an old disease: Coccidioides immitis found in Washington State soil associated with recent human infection. Clin. Infect. Dis. 2015, 60, e1–e3. [Google Scholar] [CrossRef] [PubMed]
- Cole, G.T.; Sun, S.H. Arthroconidium-spherule-endospore transformation in Coccidioides immitis. In Fungal Dimorphism: With Emphasis on Fungi Pathogenic for Humans; Szaniszlo, P.J., Harris, J.L., Eds.; Springer: Boston, MA, USA, 1985; pp. 281–333. [Google Scholar]
- Fisher, M.C.; Koenig, G.L.; White, T.J.; Taylor, J.W. Molecular and phenotypic description of Coccidioides posadasii sp. nov., previously recognized as the non-California population of Coccidioides immitis. Mycologia 2002, 94, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Levine, H.B. Purification of the spherule-endospore phase of Coccidioides immitis. Sabouraudia 1961, 1, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Muhammed, M.; Feldmesser, M.; Shubitz, L.F.; Lionakis, M.S.; Sil, A.; Wang, Y.; Glavis-Bloom, J.; Lewis, R.E.; Galgiani, J.N.; Casadevall, A.; et al. Mouse models for the study of fungal pneumonia: A collection of detailed experimental protocols for the study of Coccidioides, Cryptococcus, Fusarium, Histoplasma and combined infection due to Aspergillus-Rhizopus. Virulence 2012, 3, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.R.; Bowers, J.R.; Barker, B.M. Dust devil: The life and times of the fungus that causes Valley fever. PLoS Pathog. 2015, 11, e1004762. [Google Scholar] [CrossRef] [PubMed]
- Galgiani, J.N.; Ampel, N.M.; Blair, J.E.; Catanzaro, A.; Geertsma, F.; Hoover, S.E.; Johnson, R.H.; Kusne, S.; Lisse, J.; MacDonald, J.D.; et al. Infectious Diseases Society of America (IDSA) clinical practice guideline for the treatment of coccidioidomycosis. Clin. Infect. Dis. 2016, 63, e112–e146. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.; Barker, B.M.; Hoover, S.; Nix, D.E.; Ampel, N.M.; Frelinger, J.A.; Orbach, M.J.; Galgiani, J.N. Recent advances in our understanding of the environmental, epidemiological, immunological, and clinical dimensions of coccidioidomycosis. Clin. Microbiol. Rev. 2013, 26, 505–525. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.; Benedict, K.; Park, B.J.; Thompson, G.R., 3rd. Coccidioidomycosis: Epidemiology. Clin. Epidemiol. 2013, 5, 185–197. [Google Scholar] [PubMed]
- Wheeler, C.; Lucas, K.D.; Mohle-Boetani, J.C. Rates and risk factors for coccidioidomycosis among prison inmates, California, USA, 2011. Emerg. Infect. Dis. 2015, 21, 70–75. [Google Scholar] [CrossRef] [PubMed]
- MMWR. Increase in reported coccidioidomycosis—United States, 1998–2011. MMWR Morb. Mortal. Wkly. Rep. 2013, 62, 217–221. [Google Scholar]
- Corzo-Leon, D.E.; Armstrong-James, D.; Denning, D.W. Burden of serious fungal infections in Mexico. Mycoses 2015, 58, 34–44. [Google Scholar] [CrossRef]
- Giacomazzi, J.; Baethgen, L.; Carneiro, L.C.; Millington, M.A.; Denning, D.W.; Colombo, A.L.; Pasqualotto, A.C. The burden of serious human fungal infections in Brazil. Mycoses 2016, 59, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Thompson, G.R., 3rd. Pulmonary coccidioidomycosis. Semin. Respir. Crit. Care Med. 2011, 32, 754–763. [Google Scholar] [CrossRef] [PubMed]
- Sondermeyer, G.; Lee, L.; Gilliss, D.; Tabnak, F.; Vugia, D. Coccidioidomycosis-associated hospitalizations, California, USA, 2000–2011. Emerg. Infect. Dis. 2013, 19, 1590–1597. [Google Scholar] [CrossRef] [PubMed]
- Sondermeyer, G.L.; Lee, L.A.; Gilliss, D.; Vugia, D.J. Coccidioidomycosis-Associated Deaths in California, 2000–2013. Public Health Rep. 2016, 131, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Drutz, D.J.; Huppert, M. Coccidioidomycosis: Factors affecting the host-parasite interaction. J. Infect. Dis. 1983, 147, 372–390. [Google Scholar] [CrossRef] [PubMed]
- Erwig, L.P.; Gow, N.A. Interactions of fungal pathogens with phagocytes. Nat. Rev. Microbiol. 2016, 14, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Thompson, G.R., 3rd; Hastey, C.J.; Hodge, G.C.; Lunetta, J.M.; Pappagianis, D.; Heinrich, V. Coccidioides endospores and spherules draw strong chemotactic, adhesive, and phagocytic responses by individual human neutrophils. PLoS ONE 2015, 10, e0129522. [Google Scholar]
- Hung, C.Y.; Castro-Lopez, N.; Cole, G.T. Card9- and MyD88-mediated gamma interferon and nitric oxide production is essential for resistance to subcutaneous Coccidioides posadasii infection. Infect. Immun. 2016, 84, 1166–1175. [Google Scholar] [CrossRef] [PubMed]
- Galgiani, J.N. Differences in oxidant release by human polymorphonuclear leukocytes produced by stimulation with different phases of Coccidioides immitis. J. Infect. Dis. 1995, 172, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.Y.; del Pilar Jiménez-Alzate, M.; Gonzalez, A.; Wuthrich, M.; Klein, B.S.; Cole, G.T. Interleukin-1 receptor but not Toll-like receptor 2 is essential for MyD88-dependent Th17 immunity to Coccidioides infection. Infect. Immun. 2014, 82, 2106–2114. [Google Scholar] [CrossRef] [PubMed]
- Parkos, C.A. Neutrophil-epithelial interactions: A double-edged sword. Am. J. Pathol. 2016, 186, 1404–1416. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Viriyakosol, S.; Fierer, J.; Brown, G.D.; Kirkland, T.N. Innate immunity to the pathogenic fungus Coccidioides posadasii is dependent on Toll-like receptor 2 and Dectin-1. Infect. Immun. 2005, 73, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- Viriyakosol, S.; Jimenez Mdel, P.; Gurney, M.A.; Ashbaugh, M.E.; Fierer, J. Dectin-1 is required for resistance to coccidioidomycosis in mice. MBio 2013, 4, e00597-12. [Google Scholar] [CrossRef] [PubMed]
- Beaman, L.; Benjamini, E.; Pappagianis, D. Activation of macrophages by lymphokines: Enhancement of phagosome-lysosome fusion and killing of Coccidioides immitis. Infect. Immun. 1983, 39, 1201–1207. [Google Scholar] [PubMed]
- Beaman, L. Effects of recombinant gamma interferon and tumor necrosis factor on in vitro interactions of human mononuclear phagocytes with Coccidioides immitis. Infect. Immun. 1991, 59, 4227–4229. [Google Scholar] [PubMed]
- Hung, C.Y.; Gonzalez, A.; Wuthrich, M.; Klein, B.S.; Cole, G.T. Vaccine immunity to coccidioidomycosis occurs by early activation of three signal pathways of T helper cell response (Th1, Th2, and Th17). Infect. Immun. 2011, 79, 4511–4522. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Hung, C.Y.; Cole, G.T. Nitric oxide synthase activity has limited influence on the control of Coccidioides infection in mice. Microb. Pathog. 2011, 51, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Hung, C.Y.; Cole, G.T. Absence of phagocyte NADPH oxidase 2 leads to severe inflammatory response in lungs of mice infected with Coccidioides. Microb. Pathog. 2011, 51, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Margolis, D.A.; Viriyakosol, S.; Fierer, J.; Kirkland, T.N. The role of reactive oxygen intermediates in experimental coccidioidomycois in mice. BMC Microbiol. 2011, 11, 71. [Google Scholar] [CrossRef] [PubMed]
- Cole, G.T.; Kirkland, T.N.; Franco, M.; Zhu, S.; Yuan, L.; Sun, S.H.; Hearn, V.M. Immunoreactivity of a surface wall fraction produced by spherules of Coccidioides immitis. Infect. Immun. 1988, 56, 2695–2701. [Google Scholar] [PubMed]
- Hung, C.Y.; Yu, J.J.; Seshan, K.R.; Reichard, U.; Cole, G.T. A parasitic phase-specific adhesin of Coccidioides immitis contributes to the virulence of this respiratory Fungal pathogen. Infect. Immun. 2002, 70, 3443–3456. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.Y.; Seshan, K.R.; Yu, J.J.; Schaller, R.; Xue, J.; Basrur, V.; Gardner, M.J.; Cole, G.T. A metalloproteinase of Coccidioides posadasii contributes to evasion of host detection. Infect. Immun. 2005, 73, 6689–6703. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Hung, C.Y.; Cole, G.T. Coccidioides releases a soluble factor that suppresses nitric oxide production by murine primary macrophages. Microb. Pathog. 2011, 50, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.M.; Klein, B.S. Dendritic cells in antifungal immunity and vaccine design. Cell Host Microbe 2012, 11, 436–446. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.O.; Ampel, N.M.; Galgiani, J.N.; Lake, D.F. Dendritic cells pulsed with Coccidioides immitis lysate induce antigen-specific naive T cell activation. J. Infect. Dis. 2001, 184, 1220–1224. [Google Scholar] [CrossRef] [PubMed]
- Dionne, S.O.; Podany, A.B.; Ruiz, Y.W.; Ampel, N.M.; Galgiani, J.N.; Lake, D.F. Spherules derived from Coccidioides posadasii promote human dendritic cell maturation and activation. Infect. Immun. 2006, 74, 2415–2422. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.O.; Ampel, N.M.; Lake, D.F. Reversal of coccidioidal anergy in vitro by dendritic cells from patients with disseminated coccidioidomycosis. J. Immunol. 2002, 169, 2020–2025. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, S.; Awasthi, V.; Magee, D.M.; Coalson, J.J. Efficacy of antigen 2/proline-rich antigen cDNA-transfected dendritic cells in immunization of mice against Coccidioides posadasii. J. Immunol. 2005, 175, 3900–3906. [Google Scholar] [CrossRef] [PubMed]
- Vilekar, P.; Awasthi, V.; Lagisetty, P.; King, C.; Shankar, N.; Awasthi, S. In vivo trafficking and immunostimulatory potential of an intranasally-administered primary dendritic cell-based vaccine. BMC Immunol. 2010, 11, 60. [Google Scholar] [CrossRef] [PubMed]
- Plato, A.; Hardison, S.E.; Brown, G.D. Pattern recognition receptors in antifungal immunity. Semin. Immunopathol. 2015, 37, 97–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; LeBert, V.; Hung, C.Y.; Galles, K.; Saijo, S.; Lin, X.; Cole, G.T.; Klein, B.S.; Wuthrich, M. C-type lectin receptors differentially induce Th17 cells and vaccine immunity to the endemic mycosis of North America. J. Immunol. 2014, 192, 1107–1119. [Google Scholar] [CrossRef] [PubMed]
- Del Pilar Jimenez, A.M.; Viriyakosol, S.; Walls, L.; Datta, S.K.; Kirkland, T.; Heinsbroek, S.E.; Brown, G.; Fierer, J. Susceptibility to Coccidioides species in C57BL/6 mice is associated with expression of a truncated splice variant of Dectin-1 (Clec7a). Genes Immun. 2008, 9, 338–348. [Google Scholar] [CrossRef] [PubMed]
- Viriyakosol, S.; del Pilar Jiménez, M.; Saijo, S.; Fierer, J. Neither dectin-2 nor the mannose receptor is required for resistance to Coccidioides immitis in mice. Infect. Immun. 2014, 82, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, S. Susceptibility of TLR4-defective C3H/HeJ mice to Coccidioides posadasii infection. Med. Mycol. 2010, 48, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Dowling, J.K.; Mansell, A. Toll-like receptors: The swiss army knife of immunity and vaccine development. Clin. Transl. Immunol. 2016, 5, e85. [Google Scholar] [CrossRef] [PubMed]
- Fierer, J.; Waters, C.; Walls, L. Both CD4+ and CD8+ T cells can mediate vaccine-induced protection against Coccidioides immitis infection in mice. J. Infect. Dis. 2006, 193, 1323–1331. [Google Scholar] [CrossRef] [PubMed]
- Wuthrich, M.; Filutowicz, H.I.; Warner, T.; Deepe, G.S., Jr.; Klein, B.S. Vaccine immunity to pathogenic fungi overcomes the requirement for CD4 help in exogenous antigen presentation to CD8+ T cells: Implications for vaccine development in immune-deficient hosts. J. Exp. Med. 2003, 197, 1405–1416. [Google Scholar] [CrossRef] [PubMed]
- Nanjappa, S.G.; Heninger, E.; Wuthrich, M.; Gasper, D.J.; Klein, B.S. Tc17 cells mediate vaccine immunity against lethal fungal pneumonia in immune deficient hosts lacking CD4+ T cells. PLoS Pathog. 2012, 8, e1002771. [Google Scholar] [CrossRef] [PubMed]
- Nanjappa, S.G.; Heninger, E.; Wuthrich, M.; Sullivan, T.; Klein, B. Protective antifungal memory CD8+ T cells are maintained in the absence of CD4+ T cell help and cognate antigen in mice. J. Clin. Investig. 2012, 122, 987–999. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, J.J.; Paul, W.E. Mechanisms underlying lineage commitment and plasticity of helper CD4+ T cells. Science 2010, 327, 1098–1102. [Google Scholar] [CrossRef] [PubMed]
- Caza, T.; Landas, S. Functional and phenotypic plasticity of CD4+ T cell subsets. Biomed. Res. Int. 2015, 2015, 521957. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.A.; Magee, D.M. Coccidioidomycosis: Host response and vaccine development. Clin. Microbiol. Rev. 2004, 17, 804–839. [Google Scholar] [CrossRef] [PubMed]
- Cole, G.T.; Xue, J.M.; Okeke, C.N.; Tarcha, E.J.; Basrur, V.; Schaller, R.A.; Herr, R.A.; Yu, J.J.; Hung, C.Y. A vaccine against coccidioidomycosis is justified and attainable. Med. Mycol. 2004, 42, 189–216. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Hung, C.Y.; Yu, J.J.; Cole, G.T. Immune response of vaccinated and non-vaccinated mice to Coccidioides posadasii infection. Vaccine 2005, 23, 3535–3544. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Yu, J.J.; Hung, C.Y.; Lehmann, P.F.; Cole, G.T. Recombinant urease and urease DNA of Coccidioides immitis elicit an immunoprotective response against coccidioidomycosis in mice. Infect. Immun. 2001, 69, 2878–2887. [Google Scholar] [CrossRef] [PubMed]
- Shubitz, L.F.; Dial, S.M.; Perrill, R.; Casement, R.; Galgiani, J.N. Vaccine-induced cellular immune responses differ from innate responses in susceptible and resistant strains of mice infected with Coccidioides posadasii. Infect. Immun. 2008, 76, 5553–5564. [Google Scholar] [CrossRef] [PubMed]
- Allendorfer, R.; Brunner, G.D.; Deepe, G.S., Jr. Complex requirements for nascent and memory immunity in pulmonary histoplasmosis. J. Immunol. 1999, 162, 7389–7396. [Google Scholar] [PubMed]
- Wuthrich, M.; Warner, T.; Klein, B.S. IL-12 is required for induction but not maintenance of protective, memory responses to Blastomyces dermatitidis: Implications for vaccine development in immune-deficient hosts. J. Immunol. 2005, 175, 5288–5297. [Google Scholar] [CrossRef] [PubMed]
- Wuthrich, M.; Filutowicz, H.I.; Warner, T.; Klein, B.S. Requisite elements in vaccine immunity to Blastomyces dermatitidis: Plasticity uncovers vaccine potential in immune-deficient hosts. J. Immunol. 2002, 169, 6969–6976. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.A.; Magee, D.M. Protective immunity in coccidioidomycosis: The life cycle and biology of Coccidioides immitis. Res. Immunol. 1998, 149, 417–428. [Google Scholar] [CrossRef]
- Xue, J.; Chen, X.; Selby, D.; Hung, C.Y.; Yu, J.J.; Cole, G.T. A genetically engineered live attenuated vaccine of Coccidioides posadasii protects BALB/c mice against coccidioidomycosis. Infect. Immun. 2009, 77, 3196–3208. [Google Scholar] [CrossRef] [PubMed]
- Wuthrich, M.; Gern, B.; Hung, C.Y.; Ersland, K.; Rocco, N.; Pick-Jacobs, J.; Galles, K.; Filutowicz, H.; Warner, T.; Evans, M.; et al. Vaccine-induced protection against 3 systemic mycoses endemic to North America requires Th17 cells in mice. J. Clin. Investig. 2011, 121, 554–568. [Google Scholar] [CrossRef] [PubMed]
- Wuthrich, M.; Hung, C.Y.; Gern, B.H.; Pick-Jacobs, J.C.; Galles, K.J.; Filutowicz, H.I.; Cole, G.T.; Klein, B.S. A TCR transgenic mouse reactive with multiple systemic dimorphic fungi. J. Immunol. 2011, 187, 1421–1431. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.Y.; Hurtgen, B.J.; Bellecourt, M.; Sanderson, S.D.; Morgan, E.L.; Cole, G.T. An agonist of human complement fragment C5a enhances vaccine immunity against Coccidioides infection. Vaccine 2012, 30, 4681–4690. [Google Scholar] [CrossRef] [PubMed]
- Galgiani, J.N.; Ampel, N.M. Coccidioides immitis in patients with human immunodeficiency virus infections. Semin. Respir. Infect. 1990, 5, 151–154. [Google Scholar] [PubMed]
- Ampel, N.M.; Nesbit, L.A.; Nguyen, C.T.; Chavez, S.; Knox, K.S.; Johnson, S.M.; Pappagianis, D. Cytokine profiles from antigen-stimulated whole-blood samples among patients with pulmonary or nonmeningeal disseminated coccidioidomycosis. Clin. Vaccine Immunol. 2015, 22, 917–922. [Google Scholar] [CrossRef] [PubMed]
- Vinh, D.C.; Schwartz, B.; Hsu, A.P.; Miranda, D.J.; Valdez, P.A.; Fink, D.; Lau, K.P.; Long-Priel, D.; Kuhns, D.B.; Uzel, G.; et al. Interleukin-12 receptor beta1 deficiency predisposing to disseminated coccidioidomycosis. Clin. Infect. Dis. 2011, 52, e99–e102. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, E.P.; Hsu, A.P.; Pechacek, J.; Bax, H.I.; Dias, D.L.; Paulson, M.L.; Chandrasekaran, P.; Rosen, L.B.; Carvalho, D.S.; Ding, L.; et al. Signal transducer and activator of transcription 1 (STAT1) gain-of-function mutations and disseminated coccidioidomycosis and histoplasmosis. J. Allergy Clin. Immunol. 2013, 131, 1624–1634. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Okada, S.; Kong, X.F.; Kreins, A.Y.; Cypowyj, S.; Abhyankar, A.; Toubiana, J.; Itan, Y.; Audry, M.; Nitschke, P.; et al. Gain-of-function human STAT1 mutations impair IL-17 immunity and underlie chronic mucocutaneous candidiasis. J. Exp. Med. 2011, 208, 1635–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spinello, I.M.; Munoz, A.; Johnson, R.H. Pulmonary coccidioidomycosis. Semin. Respir. Crit. Care Med. 2008, 29, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Spring, M.; Murphy, J.; Nielsen, R.; Dowler, M.; Bennett, J.W.; Zarling, S.; Williams, J.; de la Vega, P.; Ware, L.; Komisar, J.; et al. First-in-human evaluation of genetically attenuated Plasmodium falciparum sporozoites administered by bite of Anopheles mosquitoes to adult volunteers. Vaccine 2013, 31, 4975–4983. [Google Scholar] [CrossRef] [PubMed]
- Cole, G.T.; Hurtgen, B.J.; Hung, C.Y. Progress toward a human vaccine against coccidioidomycosis. Curr. Fungal Infect. Rep. 2012, 6, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Cole, G.T.; Hung, C.Y.; Sanderson, S.D.; Hurtgen, B.J.; Wuthrich, M.; Klein, B.S.; Deepe, G.S.; Ostroff, G.R.; Levitz, S.M. Novel strategies to enhance vaccine immunity against coccidioidomycosis. PLoS Pathog. 2013, 9, e1003768. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Magee, D.M.; Quitugua, T.N.; Cox, R.A. Genetic vaccination against Coccidioides immitis: Comparison of vaccine efficacy of recombinant antigen 2 and antigen 2 cDNA. Infect. Immun. 1999, 67, 630–635. [Google Scholar] [PubMed]
- Shubitz, L.; Peng, T.; Perrill, R.; Simons, J.; Orsborn, K.; Galgiani, J.N. Protection of mice against Coccidioides immitis intranasal infection by vaccination with recombinant antigen 2/PRA. Infect. Immun. 2002, 70, 3287–3289. [Google Scholar] [CrossRef] [PubMed]
- Abuodeh, R.O.; Shubitz, L.F.; Siegel, E.; Snyder, S.; Peng, T.; Orsborn, K.I.; Brummer, E.; Stevens, D.A.; Galgiani, J.N. Resistance to Coccidioides immitis in mice after immunization with recombinant protein or a DNA vaccine of a proline-rich antigen. Infect. Immun. 1999, 67, 2935–2940. [Google Scholar] [PubMed]
- Pan, S.; Cole, G.T. Molecular and biochemical characterization of a Coccidioides immitis-specific antigen. Infect. Immun. 1995, 63, 3994–4002. [Google Scholar] [PubMed]
- Shubitz, L.F.; Yu, J.J.; Hung, C.Y.; Kirkland, T.N.; Peng, T.; Perrill, R.; Simons, J.; Xue, J.; Herr, R.A.; Cole, G.T.; et al. Improved protection of mice against lethal respiratory infection with Coccidioides posadasii using two recombinant antigens expressed as a single protein. Vaccine 2006, 24, 5904–5911. [Google Scholar] [CrossRef] [PubMed]
- Orsborn, K.I.; Shubitz, L.F.; Peng, T.; Kellner, E.M.; Orbach, M.J.; Haynes, P.A.; Galgiani, J.N. Protein expression profiling of Coccidioides posadasii by two-dimensional differential in-gel electrophoresis and evaluation of a newly recognized peroxisomal matrix protein as a recombinant vaccine candidate. Infect. Immun. 2006, 74, 1865–1872. [Google Scholar] [CrossRef] [PubMed]
- Galgiani, J.N.; Sun, S.H.; Dugger, K.O.; Ampel, N.M.; Grace, G.G.; Harrison, J.; Wieden, M.A. An arthroconidial-spherule antigen of Coccidioides immitis: Differential expression during in vitro fungal development and evidence for humoral response in humans after infection or vaccination. Infect. Immun. 1992, 60, 2627–2635. [Google Scholar] [PubMed]
- Tarcha, E.J.; Basrur, V.; Hung, C.Y.; Gardner, M.J.; Cole, G.T. Multivalent recombinant protein vaccine against coccidioidomycosis. Infect. Immun. 2006, 74, 5802–5813. [Google Scholar] [CrossRef] [PubMed]
- Tarcha, E.J.; Basrur, V.; Hung, C.Y.; Gardner, M.J.; Cole, G.T. A recombinant aspartyl protease of Coccidioides posadasii induces protection against pulmonary coccidioidomycosis in mice. Infect. Immun. 2006, 74, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Herr, R.A.; Hung, C.Y.; Cole, G.T. Evaluation of two homologous proline-rich proteins of Coccidioides posadasii as candidate vaccines against coccidioidomycosis. Infect. Immun. 2007, 75, 5777–5787. [Google Scholar] [CrossRef] [PubMed]
- Azmi, F.; Ahmad Fuaad, A.A.; Skwarczynski, M.; Toth, I. Recent progress in adjuvant discovery for peptide-based subunit vaccines. Hum. Vaccine Immunother. 2014, 10, 778–796. [Google Scholar] [CrossRef]
- Levitz, S.M.; Golenbock, D.T. Beyond empiricism: Informing vaccine development through innate immunity research. Cell 2012, 148, 1284–1292. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; McAleer, J.P.; Lin, Y.; Paterson, D.L.; Zheng, M.; Alcorn, J.F.; Weaver, C.T.; Kolls, J.K. Th17 cells mediate clade-specific, serotype-independent mucosal immunity. Immunity 2011, 35, 997–1009. [Google Scholar] [CrossRef] [PubMed]
- Hurtgen, B.J.; Hung, C.Y.; Ostroff, G.R.; Levitz, S.M.; Cole, G.T. Construction and evaluation of a novel recombinant T cell epitope-based vaccine against coccidioidomycosis. Infect. Immun. 2012, 80, 3960–3974. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Ostroff, G.R.; Lee, C.K.; Agarwal, S.; Ram, S.; Rice, P.A.; Specht, C.A.; Levitz, S.M. Relative contributions of Dectin-1 and complement to immune responses to particulate b-glucans. J. Immunol. 2012, 189, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Ostroff, G.R.; Lee, C.K.; Specht, C.A.; Levitz, S.M. Characterization and optimization of the glucan particle-based vaccine platform. Clin. Vaccine Immunol. 2013, 20, 1585–1591. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, T.N.; Fierer, J. Inbred mouse strains differ in resistance to lethal Coccidioides immitis infection. Infect. Immun. 1983, 40, 912–916. [Google Scholar] [PubMed]
- Magee, D.M.; Cox, R.A. Roles of gamma interferon and interleukin-4 in genetically determined resistance to Coccidioides immitis. Infect. Immun. 1995, 63, 3514–3519. [Google Scholar] [PubMed]
- Fierer, J. IL-10 and susceptibility to Coccidioides immitis infection. Trends Microbiol. 2006, 14, 426–427. [Google Scholar] [CrossRef] [PubMed]
- Del Pilar Jiménez, M.; Walls, L.; Fierer, J. High levels of interleukin-10 impair resistance to pulmonary coccidioidomycosis in mice in part through control of nitric oxide synthase 2 expression. Infect. Immun. 2006, 74, 3387–3395. [Google Scholar] [CrossRef] [PubMed]
- Hurtgen, B.J.; Castro-Lopez, N.; Jimenez-Alzate, M.D.; Cole, G.T.; Hung, C.Y. Preclinical identification of vaccine induced protective correlates in human leukocyte antigen expressing transgenic mice infected with Coccidioides posadasii. Vaccine 2016, 34, 5336–5343. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Galarza, F.F.; Takeshita, L.Y.; Santos, E.J.; Kempson, F.; Maia, M.H.; da Silva, A.L.; Teles e Silva, A.L.; Ghattaoraya, G.S.; Alfirevic, A.; Jones, A.R.; et al. Allele frequency net 2015 update: New features for HLA epitopes, KIR and disease and HLA adverse drug reaction associations. Nucleic Acids Res. 2015, 43, D784–D788. [Google Scholar] [CrossRef] [PubMed]
- Ru, Z.; Xiao, W.; Pajot, A.; Kou, Z.; Sun, S.; Maillere, B.; Zhao, G.; Ojcius, D.M.; Lone, Y.C.; Zhou, Y. Development of a humanized HLA-A2.1/DP4 transgenic mouse model and the use of this model to map HLA-DP4-restricted epitopes of HBV envelope protein. PLoS ONE 2012, 7, e32247. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro-Lopez, N.; Hung, C.-Y. Immune Response to Coccidioidomycosis and the Development of a Vaccine. Microorganisms 2017, 5, 13. https://doi.org/10.3390/microorganisms5010013
Castro-Lopez N, Hung C-Y. Immune Response to Coccidioidomycosis and the Development of a Vaccine. Microorganisms. 2017; 5(1):13. https://doi.org/10.3390/microorganisms5010013
Chicago/Turabian StyleCastro-Lopez, Natalia, and Chiung-Yu Hung. 2017. "Immune Response to Coccidioidomycosis and the Development of a Vaccine" Microorganisms 5, no. 1: 13. https://doi.org/10.3390/microorganisms5010013
APA StyleCastro-Lopez, N., & Hung, C. -Y. (2017). Immune Response to Coccidioidomycosis and the Development of a Vaccine. Microorganisms, 5(1), 13. https://doi.org/10.3390/microorganisms5010013