Growth and Photosynthetic Characteristics of Toxic and Non-Toxic Strains of the Cyanobacteria Microcystis aeruginosa and Anabaena circinalis in Relation to Light

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Cell Morphology and Trichome Length
2.3. Pigment Analysis
2.4. Photosynthetic Parameters
2.5. Statistical Analysis
3. Results
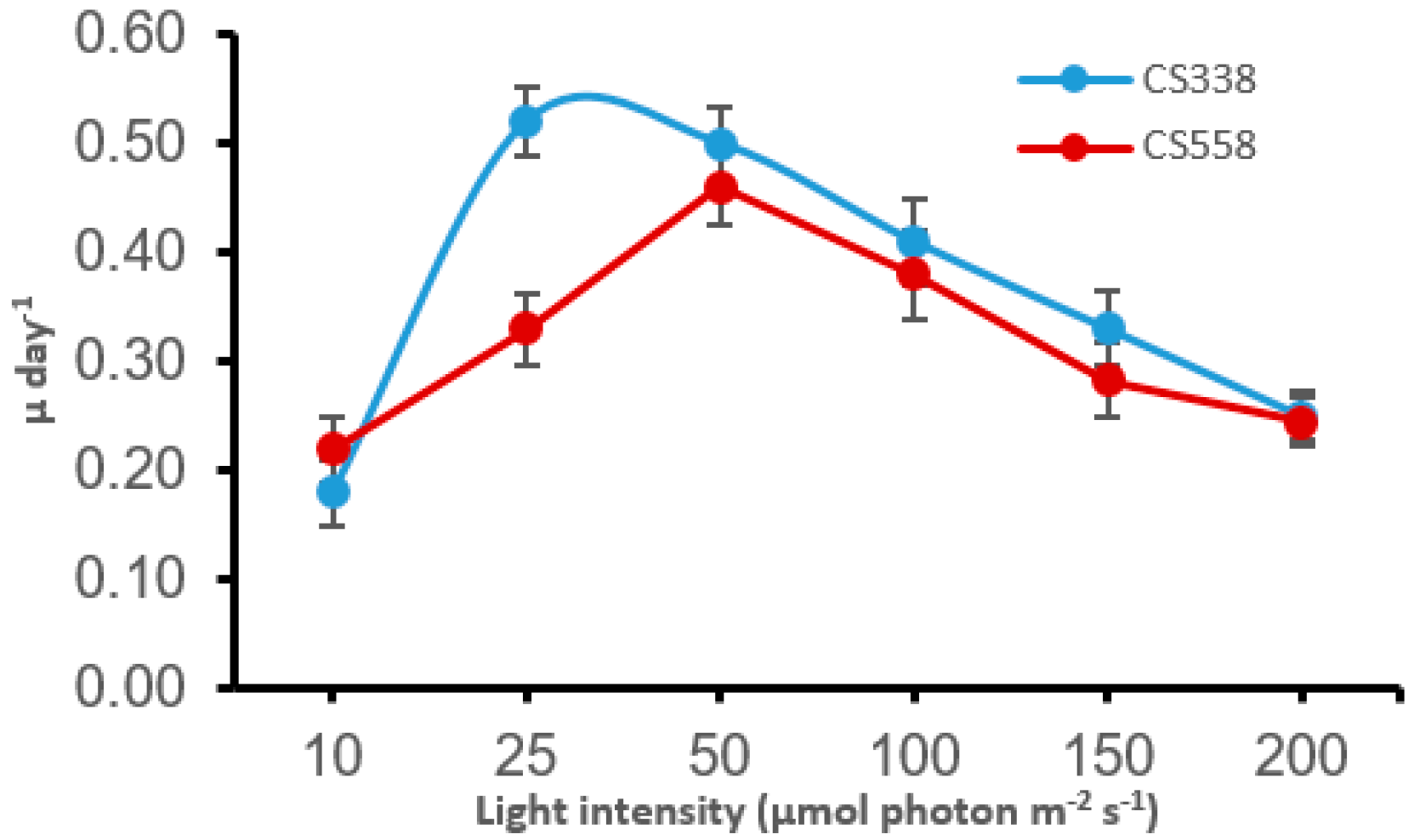
3.1. Growth and Acclimation of Cyanobacteria at Different Light Levels
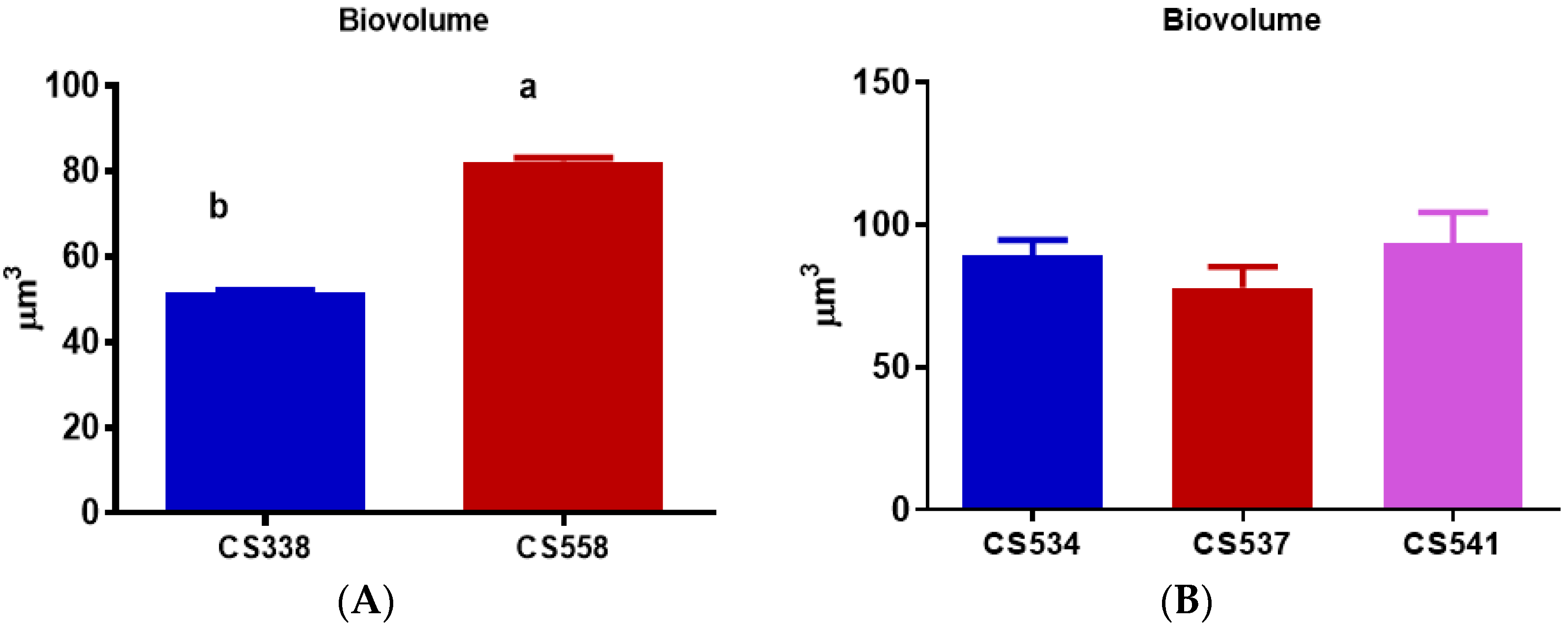
3.2. Cell Biovolume
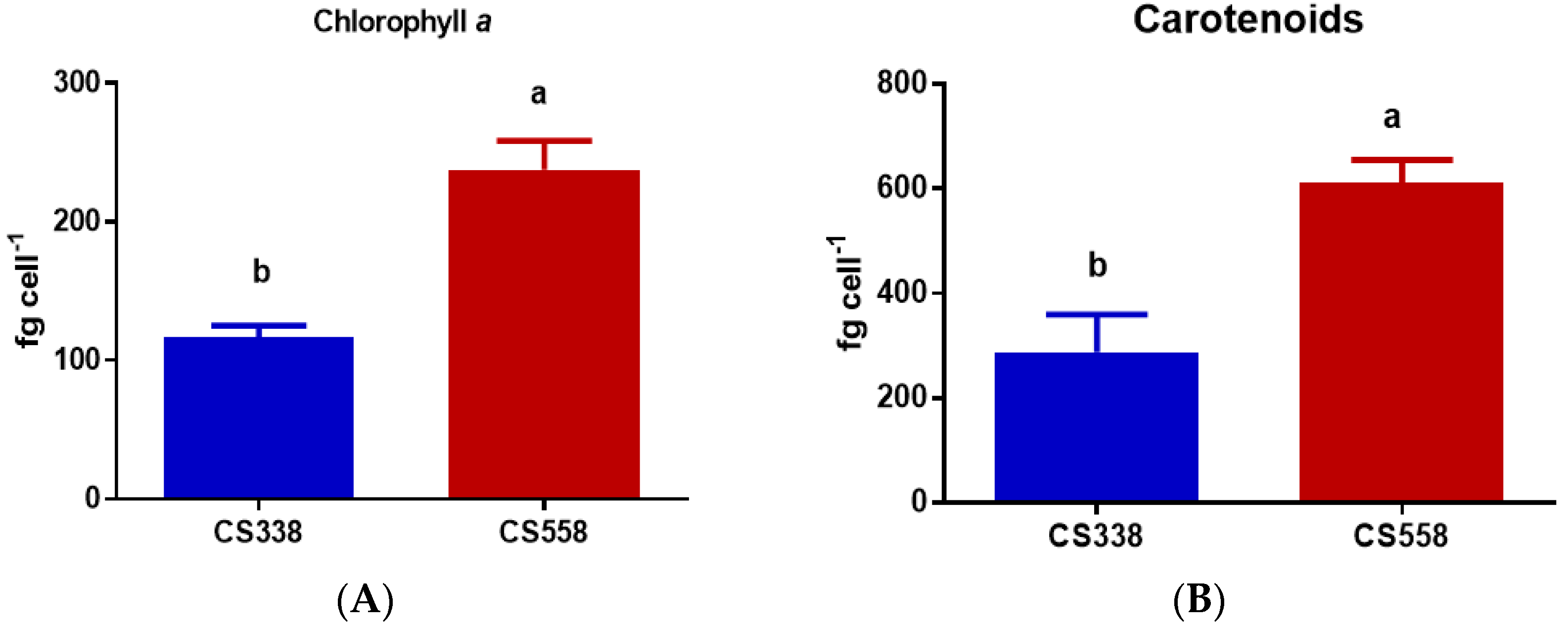
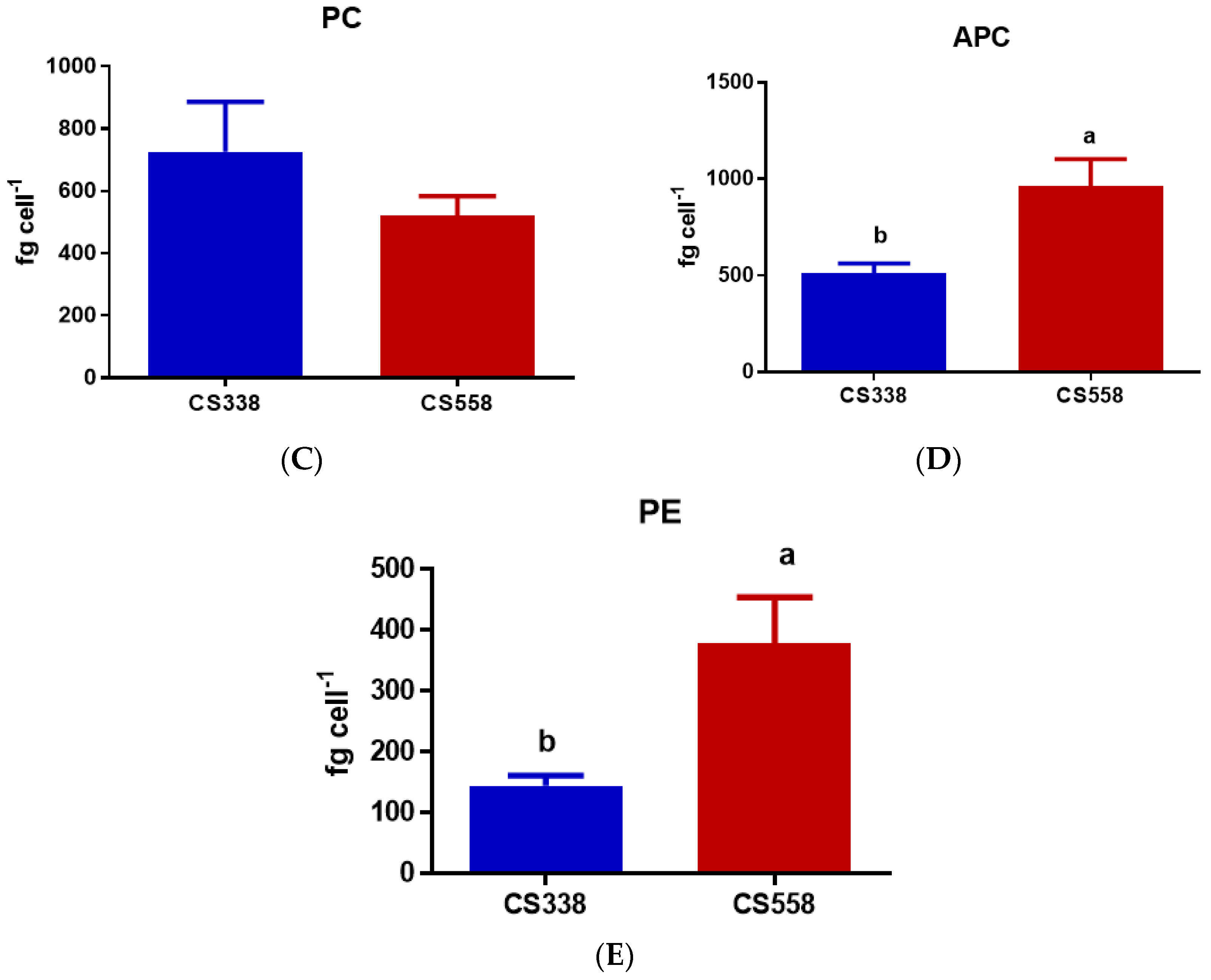
3.3. Pigments
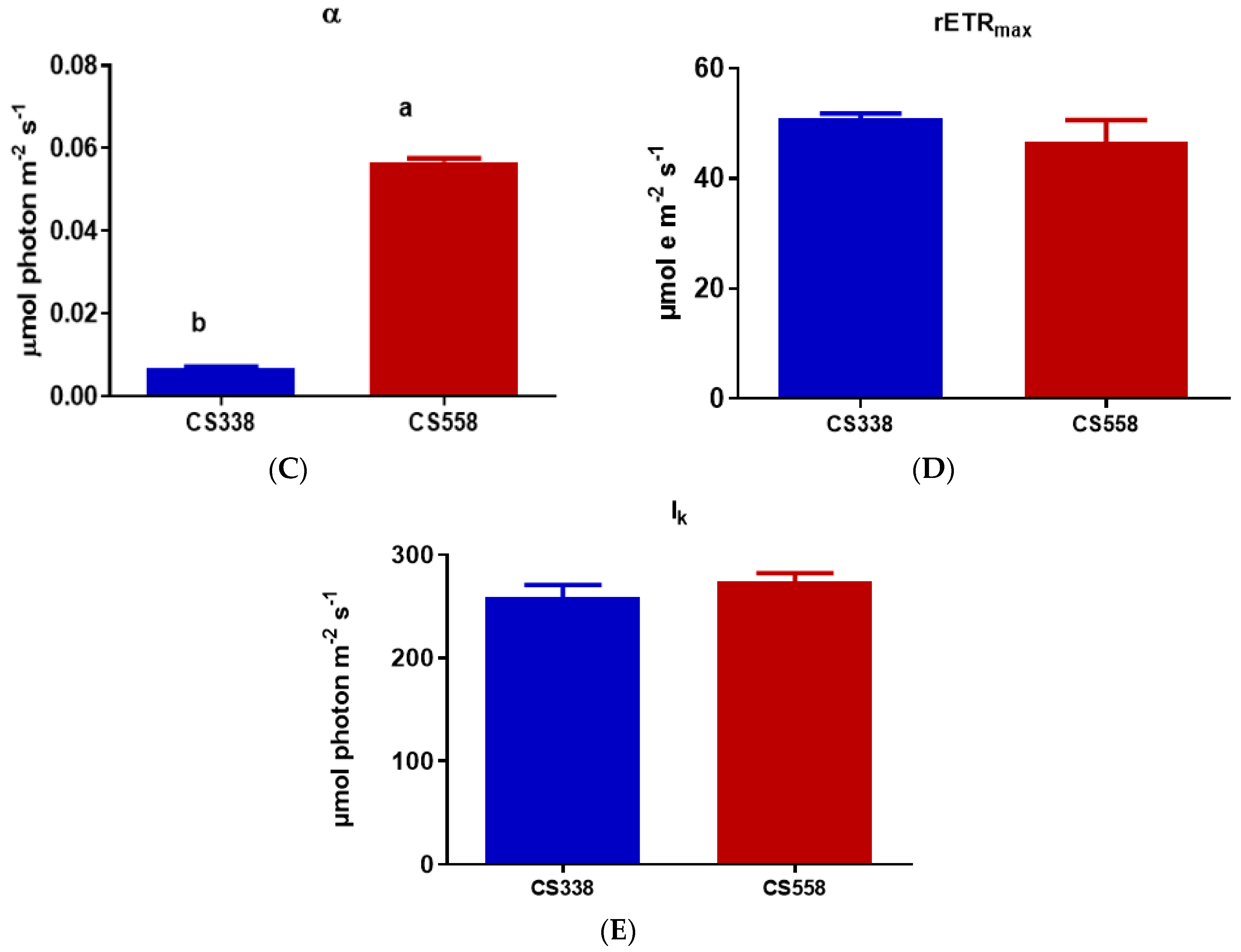
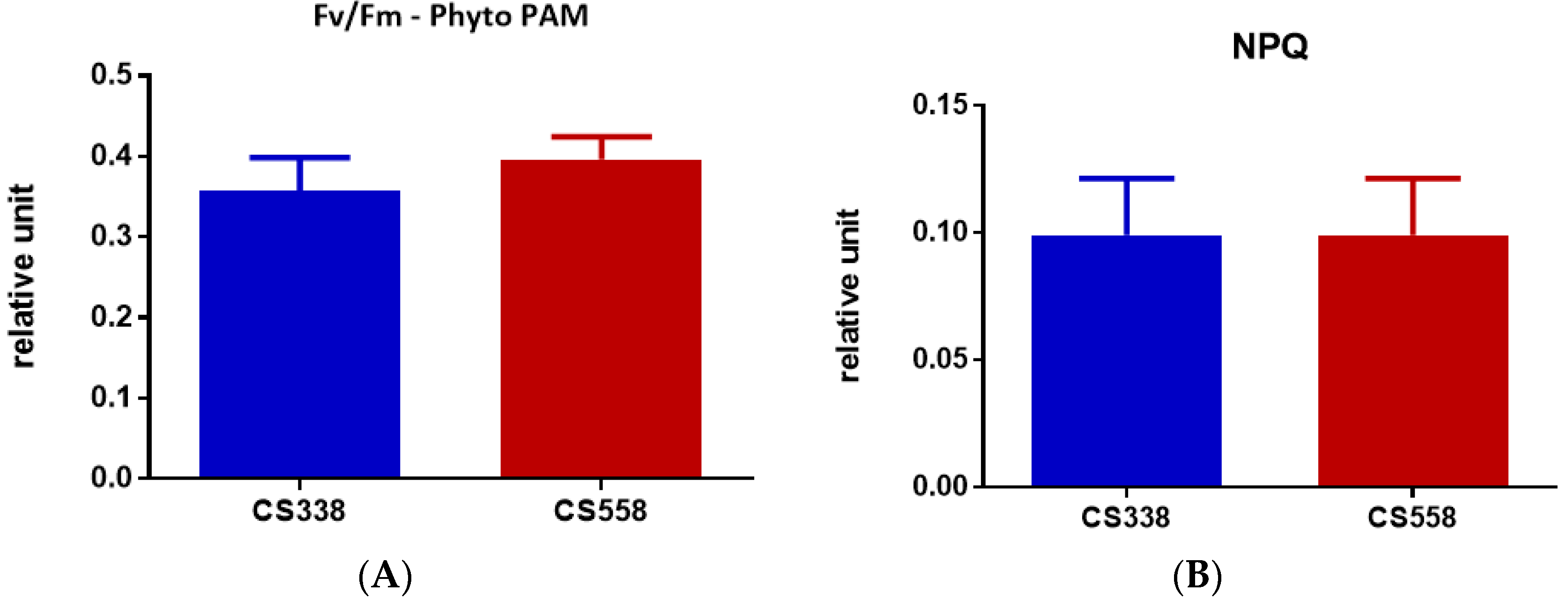
3.4. Dark Respiration (Rd) and Photosynthesis
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hamilton, D.P.; Wood, S.A.; Dietrich, D.R.; Puddick, J. Costs of harmful blooms of freshwater cyanobacteria. In Cyanobacteria: An Economic Perspective; Sharma, N.K., Rai, A.K., Stal, L.J., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2014; pp. 245–256. [Google Scholar]
- Connolly, B.; Hylands, P.; Brain, P. Economic Impact of the 2008 Blue Green Algal Bloom on the Gippsland Tourism Industry Report-April 2009; Nexus Consulting Australia: Bairnsdale, Australia, 2009. [Google Scholar]
- Wagner, C.; Adrian, R. Cyanobacteria dominance: Quantifying the effects of climate change. Limnol. Oceanogr. 2009, 54, 2460–2468. [Google Scholar] [CrossRef]
- Carey, C.C.; Ibelings, B.W.; Hoffman, E.P. Eco-physiological adaptations that favour freshwater cyanobacteria in a changing climate. Water Res. 2012, 46, 1394–1407. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Peperzak, L. Climate change and harmful algal blooms in the North Sea. Acta Oecol. 2003, 24 (Suppl. 1), 139–144. [Google Scholar] [CrossRef]
- Paul, V.J. Global warming and cyanobacterial harmful algal blooms. In Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Hudnell, H.K., Ed.; Springer Science: New York, NY, USA, 2008; Volume 619, pp. 239–257. [Google Scholar]
- Watson, S.B.; McCauley, E.; Downing, J.A. Patterns in phytoplankton taxonomic composition across temperate lakes of differing nutrient status. Limnol. Oceanogr. 1997, 42, 487–495. [Google Scholar] [CrossRef]
- Davis, T.W.; Harke, M.J.; Marcoval, M.A.; Goleski, J.; Orano-Dawson, C.; Berry, D.L.; Gobler, C.J. Effects of nitrogenous compounds and phosphorus on the growth of toxic and non-toxic strains of Microcystis during bloom events. Aquat. Microb. Ecol. 2010, 61, 149–162. [Google Scholar] [CrossRef]
- Gobler, C.J.; Davis, T.W.; Coyne, K.J.; Boyer, G.L. The interactive influences of utrient loading and zooplankton grazing on the growth and toxicity of cyanobacteria blooms in a eutrophic lake. Harmful Algae 2007, 6, 119–131. [Google Scholar] [CrossRef]
- Quiblier, C.; Wood, S.; Echenique-Subiabre, I.; Heath, M.; Villeneuve, A.; Humbert, J.F. A review of current knowledge on toxic benthic freshwater cyanobacteria-ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar]
- Kardinaal, W.E.A.; Tonk, L.; Janse, I.; Hol, S.; Slot, P.; Huisman, J.; Visser, P.M. Competition for light between toxic and nontoxic strains of the harmful cyanobacterium microcystis. Appl. Environ. Microbiol. 2007, 73, 2939–2946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapala, J.; Sivonen, K.; Luukkainen, R.; Niemela, S.I. Anatoxin-a concentration in Anabaena and Aphanizomenon under different environmental-conditions and comparison of growth by toxic and nontoxic Anabaena strains—A laboratory study. J. Appl. Phycol. 1993, 5, 581–591. [Google Scholar] [CrossRef]
- Watanabe, M.F.; Oishi, S. Effects of environmental-factors on toxicity of a cyanobacterium (Microcystis aeruginosa) under culture conditions. Appl. Environ. Microbiol. 1985, 49, 1342–1344. [Google Scholar] [PubMed]
- Dyble, J.; Tester, P.A.; Litaker, R.W. Effects of light intensity on cylindrospermopsin production in the cyanobacterial hab species Cylindrospermopsis raciborskii. Afr. J. Mar. Sci. 2006, 28, 309–312. [Google Scholar] [CrossRef]
- Sivonen, K.; Carmichael, W.W.; Namikoshi, M.; Rinehart, K.L.; Dahlem, A.M.; Niemela, S.I. Isolation and characterization of hepatotoxic microcystin homologs from the filamentous freshwater cyanobacterium Nostoc sp. strain-152. Appl. Environ. Microbiol. 1990, 56, 2650–2657. [Google Scholar] [PubMed]
- Padisak, J. Cylindrospermopsis raciborskii (Woloszynska) Seenayya et Subba Raju, an expanding, highly adaptive cyanobacterium: Worldwide distribution and review of its ecology. Arch. Hydrobiol. Suppl. 1997, 107, 563–593. [Google Scholar]
- Briand, J.F.; Leboulanger, C.; Humbert, J.F.; Bernard, C.; Dufour, P. Cylindrospermopsis raciborskii (Cyanobacteria) invasion at mid-latitudes: Selection, wide tolerance or global warming? J. Phycol. 2004, 40, 231–238. [Google Scholar] [CrossRef]
- Vidal, L.; Kruck, C. Cylindrospermopsis raciborskii (Cyanobacteria) extends its distribution to latitude 34°53′s: Taxonomical and ecological features in uruguayan eutrophic lakes. Pan-Am. J. Aquat. Sci. 2008, 3, 142–151. [Google Scholar]
- Piccini, C.; Aubriot, L.; Fabre, A.; Amaral, V.; Gonzalez-Piana, M.; Giani, A.; Figueredo, C.C.; Vidal, L.; Kruk, C.; Bonilla, S. Genetic and eco-physiological differences of south american Cylindrospermopsis raciborskii isolates support the hypothesis of multiple ecotypes. Harmful Algae 2011, 10, 644–653. [Google Scholar] [CrossRef]
- Mehnert, G.; Rucker, J.; Nicklisch, A.; Leunert, F.; Wiedner, C. Effects of thermal acclimation and photoacclimation on lipophilic pigments in an invasive and a native cyanobacterium of temperate regions. Eur. J. Phycol. 2012, 47, 182–192. [Google Scholar] [CrossRef]
- Bolch, C.J.S.; Blackburn, S.I. Isolation and purification of Australian isolates of the toxic cyanobacterium Microcystis aeruginosa Kutz. J. Appl. Phycol. 1996, 8, 5–13. [Google Scholar] [CrossRef]
- Pierangelini, M.; Stojkovic, S.; Orr, P.T.; Beardall, J. Photosynthetic characteristics of two Cylindrospermopsis raciborskii stains differing in their toxicity. J. Phycol. 2014, 50, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Hillebrand, H.; Durselen, C.D.; Kirschtel, D.; Pollingher, U.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 1999, 35, 403–424. [Google Scholar] [CrossRef]
- Ritchie, R.J. Universal chlorophyll equations for estimating chlorophylls a, b, c, and d and total chlorophylls in natural assemblages of photosynthetic organisms using acetone, methanol, or ethanol solvents. Photosynthetica 2008, 46, 115–126. [Google Scholar] [CrossRef]
- Jenssen, A. Chlorophyll and carotenoids. In Handbook of Phycological Methods, Physiological and Biochemical Methods; Hellebust, J.A., Craigie, J.S., Eds.; Cambridge University Press: London, UK, 1978; Volume 2, pp. 59–70. [Google Scholar]
- Bennett, A.; Bogorad, L. Complementary chromatic adaptation in a filamentous blue-green alga. J. Cell Biol. 1973, 58, 419–435. [Google Scholar] [CrossRef] [PubMed]
- Antenucci, J.P.; Ghadouani, A.; Burford, M.A.; Romero, J.R. The long-term effect of artificial destratification on phytoplankton species composition in a subtropical reservoir. Freshw. Biol. 2005, 50, 1081–1093. [Google Scholar] [CrossRef]
- Raven, J.A.; Finke, Z.V.; Irwin, A.J. Picophytoplankton: Bottom-up and top-down controls on ecology and evolution. Vie Milieu 2005, 55, 209–215. [Google Scholar]
- Henley, W.J. On the measurement and interpretation of photosynthetic light-response curves in algae in the context of photoinhibition and diel changes. J. Phycol. 1993, 29, 729–739. [Google Scholar] [CrossRef]
- Richardson, K.; Beardall, J.; Raven, J.A. Adaptation of unicellular algae to irradiance: An analysis of strategies. New Phytol. 1983, 93, 157–191. [Google Scholar] [CrossRef]
- MacIntyre, H.L.; Kana, T.M.; Anning, T.; Geider, R.J. Photoacclimation of photosynthesis irradiance response curves and photosynthetic pigments in microalgae and cyanobacteria. J. Phycol. 2002, 38, 17–38. [Google Scholar] [CrossRef]
- Gao, K.; Yu, H.; Brown, M.T. Solar PAR and UV radiation affects the physiology and morphology of the cyanobacterium Anabaena sp. Pcc 7120. J. Photochem. Photobiol. B Biol. 2007, 89, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Kirilovsky, D. Photoprotection in cyanobacteria: The orange carotenoid protein (ocp)-related non-photochemicalquenching mechanism. Photosynth. Res. 2007, 93, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S.; Grossman, A. Photoprotection in cyanobacteria: Regulation of light harvesting. Photochem. Photobiol. 2008, 84, 1410–1420. [Google Scholar] [CrossRef] [PubMed]
- Campbell, D.; Hurry, V.; Clarke, A.K.; Gustafsson, P.; Gunnar, O. Chlorophyll fluorescence analysis of cyanobacterial photosynthesis and acclimation. Microbiol. Mol. Biol. Rev. 1998, 62, 667–683. [Google Scholar] [PubMed]
- Campbell, D.; Oquist, G. Predicting light acclimation in cyanobacteria from non-photochemical quenching of psii fluorescence, which reflects state transitions in these organisms. Plant Physiol. 1996, 111, 1293–1298. [Google Scholar] [CrossRef] [PubMed]












© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.A.; Beardall, J. Growth and Photosynthetic Characteristics of Toxic and Non-Toxic Strains of the Cyanobacteria Microcystis aeruginosa and Anabaena circinalis in Relation to Light. Microorganisms 2017, 5, 45. https://doi.org/10.3390/microorganisms5030045
Islam MA, Beardall J. Growth and Photosynthetic Characteristics of Toxic and Non-Toxic Strains of the Cyanobacteria Microcystis aeruginosa and Anabaena circinalis in Relation to Light. Microorganisms. 2017; 5(3):45. https://doi.org/10.3390/microorganisms5030045
Chicago/Turabian StyleIslam, M. Ashraful, and John Beardall. 2017. "Growth and Photosynthetic Characteristics of Toxic and Non-Toxic Strains of the Cyanobacteria Microcystis aeruginosa and Anabaena circinalis in Relation to Light" Microorganisms 5, no. 3: 45. https://doi.org/10.3390/microorganisms5030045
APA StyleIslam, M. A., & Beardall, J. (2017). Growth and Photosynthetic Characteristics of Toxic and Non-Toxic Strains of the Cyanobacteria Microcystis aeruginosa and Anabaena circinalis in Relation to Light. Microorganisms, 5(3), 45. https://doi.org/10.3390/microorganisms5030045