Multiple Antibiotic-Resistant, Extended Spectrum-β-Lactamase (ESBL)-Producing Enterobacteria in Fresh Seafood

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Isolation, and Identification of Enterobacteria from Seafood
2.2. Antibiotic Susceptibility Tests
2.3. Detection of ESBL Phenotypes
2.4. Detection of Resistance Genes by PCR
3. Results
3.1. Species Diversity of ESBL+ Enterobacteria Isolated from Seafood
3.2. Distribution of ESBL+ Enterobacteria in Seafood
3.3. Antibiotic Resistance of Isolates
3.4. PCR Detection of β-LactamaseGenes
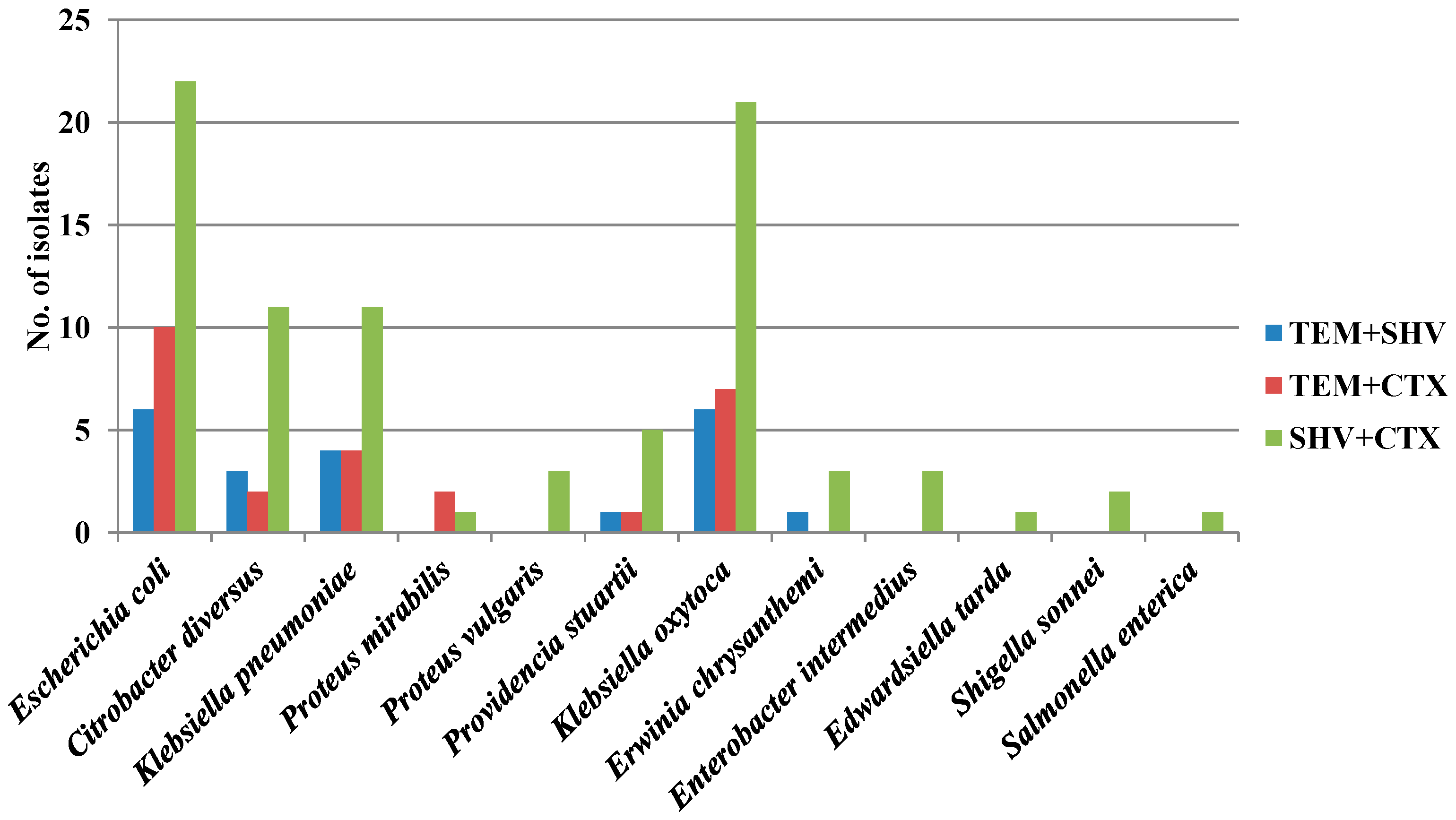
3.5. Co-Occurrence of ESBL Genes in Enterobacteria
3.6. blaNDM-Harboring Enterobacteria in Seafood
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Arias, C.A.; Murray, B.E. Antibiotic-resistant bugs in the 21st century—A clinical super-challenge. N. Engl. J. Med. 2009, 360, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, U.; Aggarwal, R. Extended spectrum-lactamases (ESBL)—An emerging threat to clinical therapeutics. Indian J. Med. Microbiol. 2004, 22, 75–80. [Google Scholar] [PubMed]
- Pitout, J.D.D.; Laupland, K.B. Extended-spectrum beta-lactamase-producing Enterobacteriaceae: An emerging public-health concern. Lancet Infect. Dis. 2008, 8, 159–166. [Google Scholar] [CrossRef]
- Jacoby, G.; Bush, K. β-lactam resistance in the 21st century. In Frontiers in Antimicrobial Resistance; American Society for Microbiology: Washington, DC, USA, 2005. [Google Scholar]
- Paterson, D.L.; Bonomo, R.A. Extended-spectrum beta-lactamases: A clinical update. Clin. Microbiol. Rev. 2005, 18, 657–686. [Google Scholar] [CrossRef] [PubMed]
- Chong, Y.; Ito, Y.; Kamimura, T. Genetic evolution and clinical impact in extended-spectrum β-lactamase-producing Escherichia coli and Klebsiella pneumoniae. Infect. Genet. Evol. 2011, 11, 1499–1504. [Google Scholar] [CrossRef] [PubMed]
- Philippon, A.; Labia, R.; Jacoby, G. Extended-spectrum beta-lactamases. Antimicrob. Agents Chemother. 1989, 33, 1131–1136. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, P.; Naas, T.; Poirel, L. Global spread of Carbapenemase-producing Enterobacteriaceae. Emerg. Infect. Dis. 2011, 17, 1791–1798. [Google Scholar] [CrossRef] [PubMed]
- Queenan, A.M.; Bush, K. Carbapenemases: The versatile beta-lactamases. Clin. Microbiol. Rev. 2007, 20, 440–458. [Google Scholar] [CrossRef] [PubMed]
- Yong, D.; Toleman, M.A.; Giske, C.G.; Cho, H.S.; Sundman, K.; Lee, K.; Walsh, T.R. Characterization of a new metallo-beta-lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob. Agents Chemother. 2009, 53, 5046–5054. [Google Scholar] [CrossRef] [PubMed]
- Bonomo, R.A. New Delhi metallo-β-lactamase and multidrug resistance: A global SOS? Clin. Infect. Dis. 2011, 52, 485–487. [Google Scholar] [CrossRef] [PubMed]
- Moellering, R.C. NDM-1—A cause for worldwide concern. N. Engl. J. Med. 2010, 363, 2377–2379. [Google Scholar] [CrossRef] [PubMed]
- Isozumi, R.; Yoshimatsu, K.; Yamashiro, T.; Hasebe, F.; Nguyen, B.M.; Ngo, T.C.; Yasuda, S.P.; Koma, T.; Shimizu, K.; Arikawa, J. blaNDM-1-positive Klebsiella pneumoniae from environment, Vietnam. Emerg. Infect. Dis. 2012, 18, 1383–1385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Qiu, S.; Wang, Y.; Qi, L.; Hao, R.; Liu, X.; Shi, Y.; Hu, X.; An, D.; Li, Z.; et al. Higher isolation of NDM-1 producing Acinetobacter baumannii from the sewage of the hospitals in Beijing. PLoS ONE 2014, 8, e64857. [Google Scholar] [CrossRef] [PubMed]
- Ponce, E.; Khan, A.A.; Cheng, C.-M.; Summage-West, C.; Cerniglia, C.E. Prevalence and characterization of Salmonella enterica serovar Weltevreden from imported seafood. Food Microbiol. 2008, 25, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Sanath Kumar, H.; Otta, S.K.; Karunasagar, I.; Karunasagar, I. Detection of Shiga-toxigenic Escherichia coli (STEC) in fresh seafood and meat marketed in Mangalore, India by PCR. Lett. Appl. Microbiol. 2001, 33, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Sanath Kumar, H.; Sunil, R.; Venugopal, M.N.; Karunasagar, I.; Karunasagar, I. Detection of Salmonella spp. in tropical seafood by polymerase chain reaction. Int. J. Food Microbiol. 2003, 88, 91–95. [Google Scholar] [CrossRef]
- Shabarinath, S.; Sanath Kumar, H.; Khushiramani, R.; Karunasagar, I.; Karunasagar, I. Detection and characterization of Salmonella associated with tropical seafood. Int. J. Food Microbiol. 2007, 114, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Adesiji, Y.O.; Deekshit, V.K.; Karunasagar, I. Antimicrobial-resistant genes associated with Salmonella spp. isolated from human, poultry, and seafood sources. Food Sci. Nutr. 2014, 2, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Boss, R.; Overesch, G.; Baumgartner, A. Antimicrobial Resistance of Escherichia coli, Enterococci, Pseudomonas aeruginosa, and Staphylococcus aureus from Raw Fish and Seafood Imported into Switzerland. J. Food Prot. 2016, 79, 1240–1246. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhou, H.; Qin, L.; Pang, Z.; Qin, T.; Ren, H.; Pan, Z.; Zhou, J. Frequency, Antimicrobial Resistance and Genetic Diversity of Klebsiella pneumoniae in Food Samples. PLoS ONE 2016, 11, e0153561. [Google Scholar] [CrossRef] [PubMed]
- Le, Q.P.; Ueda, S.; Nguyen, T.N.H.; Dao, T.V.K.; Van Hoang, T.A.; Tran, T.T.N.; Hirai, I.; Nakayama, T.; Kawahara, R.; Do, T.H.; et al. Characteristics of Extended-Spectrum β-Lactamase-Producing Escherichia coli in Retail Meats and Shrimp at a Local Market in Vietnam. Foodborne Pathog. Dis. 2015, 12, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.S.; Parvathi, A.; Karunasagar, I.; Karunasagar, I. Prevalence and antibiotic resistance of Escherichia coli in tropical seafood. World J. Microbiol. Biotechnol. 2005, 21, 619–623. [Google Scholar] [CrossRef]
- Bushnell, G.; Mitrani-Gold, F.; Mundy, L.M. Emergence of New Delhi metallo-β-lactamase type 1-producing enterobacteriaceae and non-enterobacteriaceae: Global case detection and bacterial surveillance. Int. J. Infect. Dis. 2013, 17, e325–e333. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, R.; Panda, S.; Singh, D.V. Emergence and dissemination of antibiotic resistance: A global problem. Indian J. Med. Microbiol. 2012, 30, 384–390. [Google Scholar] [PubMed]
- Singh, A.S.; Lekshmi, M.; Nayak, B.B.; Kumar, S.H. Isolation of Escherichia coli harboring blaNDM-5 from fresh fish in India. J. Microbiol. Immunol. Infect. 2014. [Google Scholar] [CrossRef] [PubMed]
- MacFaddin, J.F. Biochemical Tests for Identification of Medical Bacteria, 2nd ed.; Williams and Wilkins: Baltimore, MD, USA, 1980. [Google Scholar]
- Pandit, L.; Kumar, S.; Karunasagar, I.; Karunasagar, I. Diagnosis of partially treated culture-negative bacterial meningitis using 16S rRNA universal primers and restriction endonuclease digestion. J. Med. Microbiol. 2005, 54, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. CLSI Performance Standards for Antimicrobial Susceptibility Testing; nineteenth informational supplement M100-S19, Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2009. [Google Scholar]
- Edelstein, M.; Pimkin, M.; Palagin, I.; Edelstein, I.; Stratchounski, L. Prevalence and molecular epidemiology of CTX-M extended-spectrum beta-lactamase-producing Escherichia coli and Klebsiella pneumoniae in Russian hospitals. Antimicrob. Agents Chemother. 2003, 47, 3724–3732. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, J.K.; Jay, C.; Metchock, B.; Berkowitz, F.; Weigel, L.; Crellin, J.; Steward, C.; Hill, B.; Medeiros, A.A.; Tenover, F.C. Evolution of extended-spectrum beta-lactam resistance (SHV-8) in a strain of Escherichia coli during multiple episodes of bacteremia. Antimicrob. Agents Chemother. 1997, 41, 647–653. [Google Scholar] [PubMed]
- Mabilat, C.; Goussard, S. PCR detection and identification of genes for extended spectrum beta-lactamase. In Diagnostic Molecular Microbiology: Principles and Applications; Persing, D.H., Smith, T.F., Tenover, F.C., White, T.J., Eds.; American Society for Microbiology: Washington, DC, USA, 1993; pp. 553–563. [Google Scholar]
- Kumaran, S.; Deivasigamani, B.; Alagappan, K.; Sakthivel, M.; Karthikeyan, R. Antibiotic resistant Esherichia coli strains from seafood and its susceptibility to seaweed extracts. Asian Pac. J. Trop. Med. 2010, 3, 977–981. [Google Scholar] [CrossRef]
- Nordmann, P.; Poirel, L.; Carrër, A.; Toleman, M.A.; Walsh, T.R. How to detect NDM-1 producers. J. Clin. Microbiol. 2011, 49, 718–721. [Google Scholar] [CrossRef] [PubMed]
- Chandran, A.; Hatha, A.A.M.; Varghese, S.; Sheeja, K.M. Prevalence of Multiple Drug Resistant Escherichia coli Serotypes in a Tropical Estuary, India. Microbes Environ. 2008, 23, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Paterson, D.L. Resistance in gram-negative bacteria: Enterobacteriaceae. Am. J. Infect. Control 2006, 34, S20–S28. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, R. Growing group of extended-spectrum beta-lactamases: The CTX-M enzymes. Antimicrob. Agents Chemother. 2004, 48, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.-H.; Lee, J.-H.; Park, S.-H.; Song, M.-O.; Park, S.-H.; Jung, H.-W.; Park, G.-Y.; Choi, S.-M.; Kim, M.-S.; Chae, Y.-Z.; et al. Antimicrobial resistance profiles among Escherichia coli strains isolated from commercial and cooked foods. Int. J. Food Microbiol. 2012, 159, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Miranda, C.D.; Zemelman, R. Antibiotic resistant bacteria in fish from the Concepción Bay, Chile. Mar. Pollut. Bull. 2001, 42, 1096–1102. [Google Scholar] [CrossRef]
- Walsh, T.R.; Toleman, M.A. The emergence of pan-resistant Gram-negative pathogens merits a rapid global political response. J. Antimicrob. Chemother. 2012, 67, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Talbot, G.H.; Bradley, J.; Edwards, J.E.; Gilbert, D.; Scheld, M.; Bartlett, J.G.; Antimicrobial Availability Task Force of the Infectious Diseases Society of America. Bad bugs need drugs: An update on the development pipeline from the Antimicrobial Availability Task Force of the Infectious Diseases Society of America. Clin. Infect. Dis. 2006, 42, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Wattal, C.; Goel, N.; Oberoi, J.K.; Raveendran, R.; Prasad, K.J. A ten year analysis of multi-drug resistant blood stream infections caused by Escherichia coli & Klebsiella pneumoniae in a tertiary care hospital. Indian J. Med. Res. 2012, 135, 907–912. [Google Scholar] [PubMed]
- Bradford, P.A. Extended-spectrum beta-lactamases in the 21st century: Characterization, epidemiology, and detection of this important resistance threat. Clin. Microbiol. Rev. 2001, 14, 933–951. [Google Scholar] [CrossRef] [PubMed]
- Djordjevic, S.P.; Stokes, H.W.; Roy Chowdhury, P. Mobile elements, zoonotic pathogens and commensal bacteria: Conduits for the delivery of resistance genes into humans, production animals and soil microbiota. Front. Microbiol. 2013, 4, 86. [Google Scholar] [CrossRef] [PubMed]
- Berglund, B. Environmental dissemination of antibiotic resistance genes and correlation to anthropogenic contamination with antibiotics. Infect. Ecol. Epidemiol. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Mesa, R.J.; Blanc, V.; Blanch, A.R.; Cortés, P.; González, J.J.; Lavilla, S.; Miró, E.; Muniesa, M.; Saco, M.; Tórtola, M.T.; et al. Extended-spectrum beta-lactamase-producing Enterobacteriaceae in different environments (humans, food, animal farms and sewage). J. Antimicrob. Chemother. 2006, 58, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Van, T.T.H.; Chin, J.; Chapman, T.; Tran, L.T.; Coloe, P.J. Safety of raw meat and shellfish in Vietnam: An analysis of Escherichia coli isolations for antibiotic resistance and virulence genes. Int. J. Food Microbiol. 2008, 124, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Maltezou, H.C. Metallo-beta-lactamases in Gram-negative bacteria: introducing the era of pan-resistance? Int. J. Antimicrob. Agents 2009, 33, 405.e1–405.e7. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Martínez, L.; González-López, J.J. Carbapenemases in Enterobacteriaceae: Types and molecular epidemiology. Enfermedades Infecciosas y Microbiologia Clinica 2014, 32, 4–9. [Google Scholar] [CrossRef]
- Khan, A.U.; Maryam, L.; Zarrilli, R. Structure, Genetics and Worldwide Spread of New Delhi Metallo-β-lactamase (NDM): A threat to public health. BMC Microbiol. 2017, 17, 101. [Google Scholar] [CrossRef] [PubMed]
- Castanheira, M.; Deshpande, L.M.; Mathai, D.; Bell, J.M.; Jones, R.N.; Mendes, R.E. Early dissemination of NDM-1- and OXA-181-producing Enterobacteriaceae in Indian hospitals: Report from the SENTRY Antimicrobial Surveillance Program, 2006–2007. Antimicrob. Agents Chemother. 2011, 55, 1274–1278. [Google Scholar] [CrossRef] [PubMed]
- Kazi, M.; Drego, L.; Nikam, C.; Ajbani, K.; Soman, R.; Shetty, A.; Rodrigues, C. Molecular characterization of carbapenem-resistant Enterobacteriaceae at a tertiary care laboratory in Mumbai. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 467–472. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Species | Number Isolated (ESBL+) |
|---|---|
| Escherichia coli | 66 (53) |
| Klebsiella oxytoca | 34 (27) |
| Klebsiella pneumoniae | 30 (23) |
| Citrobacter diversus | 28 (22) |
| Erwinia chrysanthemi | 16 (12) |
| Proteus vulgaris | 13 (10) |
| Proteus mirabilis | 8 (6) |
| Providencia stuartii | 8 (6) |
| Enterobacter intermedius | 7 (5) |
| Edwardsiella tarda | 2 (2) |
| Shigella sonnei | 2 (2) |
| Salmonella enterica | 1 (1) |
| Total | 215 (169) |
| Sample Types (Number Analyzed) | No. of Enterobacteria Isolated | No. (%) of ESBL+ Enterobacteria |
|---|---|---|
| Fish (14) | ||
| Sardinella albella (5) | 41 | 31 (75.60) |
| Coilia dussumieri (3) | 39 | 29 (74.35) |
| Escualosa thoracata (1) | 14 | 14 (100) |
| Epinephelus diacanthus (1) | 15 | 15 (100) |
| Harpadon nehereus (1) | 11 | 9 (81.81) |
| Terapon jarbua (1) | 7 | 7 (100) |
| Scomberomorus commerson (1) | 10 | 10 (100) |
| Otolithes cuvieri (1) | 16 | 16 (100) |
| Shrimp (3) | ||
| Acetes indicus (1) | 12 | 12(100) |
| Metapenaeus dobsonii (2) | 19 | 14 (73) |
| Clam (1) | ||
| Meretrix meretrix | 16 | 8 (50) |
| Squid (1) | ||
| Loligo duvauceli | 15 | 4 (26) |
| Total | 215 | 169 |
| Antibiotics Tested | No. (%) Resistant a |
|---|---|
| Cefoxitin (CX) | 34 (20.12%) |
| Cefotaxime (CTX) | 165 (97.63%) |
| Ceftazidime (CAZ) | 154 (91.12%) |
| Cefpodoxime (CPD) | 156 (92.30%) |
| Imipenem (IPM) | 18 (10.65%) |
| Ertapenem (ETP) | 69 (40.82%) |
| Meropenem (MRP) | 53 (31.36%) |
| Ciprofloxacin (CIP) | 27 (15.98%) |
| Aztreonam (AT) | 110 (65.08%) |
| Amox/clav (AMC) | 65 (38.46%) |
| Piperacillin/Tazobactam (TZP) | 69 (40.82%) |
| Species | Number of Isolates Tested | Distribution of ESBL Genes | ||
|---|---|---|---|---|
| blaCTX | blaSHV | blaTEM | ||
| Escherichia coli | 53 | 41 | 29 | 24 |
| Klebsiella oxytoca | 27 | 24 | 24 | 16 |
| Klebsiella pneumoniae | 23 | 15 | 17 | 15 |
| Citrobacter diversus | 22 | 15 | 11 | 8 |
| Erwinia chrysanthemi | 12 | 6 | 7 | 6 |
| Proteus vulgaris | 10 | 9 | 3 | 0 |
| Proteus mirabilis | 6 | 6 | 1 | 3 |
| Providencia stuartii | 6 | 6 | 6 | 3 |
| Enterobacter intermedius | 5 | 3 | 4 | 0 |
| Edwardsiella tarda | 2 | 2 | 2 | 0 |
| Shigella sonnei | 2 | 2 | 2 | 0 |
| Salmonella enterica | 1 | 1 | 1 | 0 |
| Total | 169 | 130 | 107 | 75 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanjit Singh, A.; Lekshmi, M.; Prakasan, S.; Nayak, B.B.; Kumar, S. Multiple Antibiotic-Resistant, Extended Spectrum-β-Lactamase (ESBL)-Producing Enterobacteria in Fresh Seafood. Microorganisms 2017, 5, 53. https://doi.org/10.3390/microorganisms5030053
Sanjit Singh A, Lekshmi M, Prakasan S, Nayak BB, Kumar S. Multiple Antibiotic-Resistant, Extended Spectrum-β-Lactamase (ESBL)-Producing Enterobacteria in Fresh Seafood. Microorganisms. 2017; 5(3):53. https://doi.org/10.3390/microorganisms5030053
Chicago/Turabian StyleSanjit Singh, Asem, Manjusha Lekshmi, Sreepriya Prakasan, Binaya Bhusan Nayak, and Sanath Kumar. 2017. "Multiple Antibiotic-Resistant, Extended Spectrum-β-Lactamase (ESBL)-Producing Enterobacteria in Fresh Seafood" Microorganisms 5, no. 3: 53. https://doi.org/10.3390/microorganisms5030053
APA StyleSanjit Singh, A., Lekshmi, M., Prakasan, S., Nayak, B. B., & Kumar, S. (2017). Multiple Antibiotic-Resistant, Extended Spectrum-β-Lactamase (ESBL)-Producing Enterobacteria in Fresh Seafood. Microorganisms, 5(3), 53. https://doi.org/10.3390/microorganisms5030053