Lactobacillus sakei: A Starter for Sausage Fermentation, a Protective Culture for Meat Products

{kind=link}
Abstract
:1. Introduction
2. Adaptation of L. sakei to Its Various Habitats
2.1. Ability to Use Nutrients Encountered in Meat to Produce Metabolites
2.1.1. Use of Amino-Acids
2.1.2. Use of Carbon Sources
2.1.3. Other Nutrients
2.2. Ability to Resist to Various Stresses
3. Intra-species Diversity of L. sakei
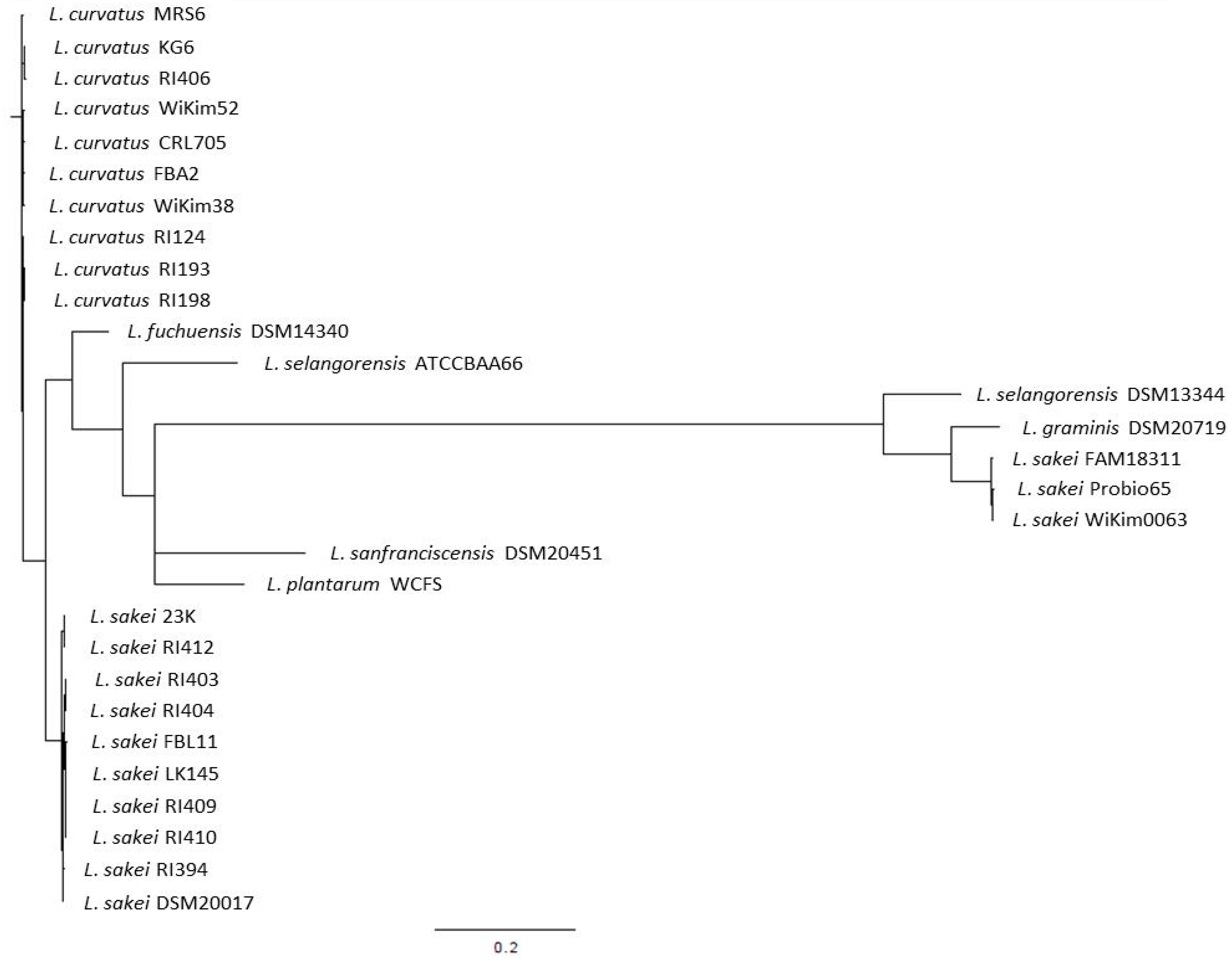
3.1. The L. sakei Clade
3.2. The Genomic Diversity among L. sakei Strains
4. L. sakei: A Starter for Fermented Sausages
5. L. sakei and Its Use as Protective Culture
6. L. sakei: The Bad Boy
7. Conclusions
Author Contributions
Conflicts of Interest
References
- Katagiri, H.; Kitahara, K.; Fukami, K. The characteristics of the lactic acid bacteria isolated from moto, yeast mashes for sake´ manufacture. IV. Classification of the lactic acid bacteria. Bull. Agric. Chem. Soc. 1934, 10, 156–157. [Google Scholar] [CrossRef]
- Moroni, A.V.; Arendt, E.K.; Dal Bello, F. Biodiversity of lactic acid bacteria and yeasts in spontaneously-fermented buckwheat and teff sourdoughs. Food Microbiol. 2011, 28, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Lhomme, E.; Mezaize, S.; Bonnand Ducasse, M.; Chiron, H.; Champomier-Vergès, M.C.; Chaillou, S.; Zagorec, M.; Dousset, X.; Onno, B. A polyphasic approach to study the dynamics of microbial population of an organic wheat sourdough during its conversion to gluten-free sourdough. Int. Microbiol. 2014, 17, 1–9. [Google Scholar] [PubMed]
- Jung, J.Y.; Lee, S.H.; Jeon, C.O. Kimchi microflora: History, Current Status, and Perspectives for Industrial Kimchi Production. Appl. Microbiol. Biotechnol. 2014, 98, 2385–2393. [Google Scholar] [CrossRef] [PubMed]
- Coda, R.; Kianjam, M.; Pontonio, E.; Verni, M.; Di Cagno, R.; Katina, K.; Rizzello, C.G.; Gobbetti, M. Sourdough-type propagation of faba bean flour: Dynamics of Microbial Consortia and Biochemical Implications. Int. J. Food Microbiol. 2017, 248, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Chaillou, S.; Chaulot-Talmon, A.; Caekebeke, H.; Cardinal, M.; Christieans, S.; Denis, C.; Desmonts, M.H.; Dousset, X.; Feurer, C.; Hamon, E.; et al. Origin and ecological selection of core and food-specific bacterial communities associated with meat and seafood spoilage. ISME J. 2015, 9, 1105–1118. [Google Scholar] [CrossRef] [PubMed]
- Najjari, A.; Ouzari, I.; Boudabous, A.; Zagorec, M. Method for reliable isolation of a collection of Lactobacillus sakei strains originating from Tunisian seafood and meat products. Int. J. Food Microbiol. 2008, 121, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Lucquin, I.; Zagorec, M.; Champomier-Vergès, M.C.; Chaillou, S. Taxonomic structure of lactic acid bacteria community in non-fermented beef meat ecosystems and natural population dynamic of the meat starter species Lactobacillus sakei assessed by genotype fingerprinting. Food Microbiol. 2012, 29, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Dal Bello, F.; Walter, J.; Hammes, W.; Hertel, C. Increased complexity of the species composition of lactic acid bacteria in human feces revealed by alternative incubation condition. Microb. Ecol. 2003, 45, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Chiaramonte, F.; Blugeon, S.; Chaillou, S.; Langella, P.; Zagorec, M. Behavior of the meat-borne bacterium Lactobacillus sakei during its transit through the gastrointestinal tracts of axenic and conventional mice. Appl. Environ. Microbiol. 2009, 75, 4498–4505. [Google Scholar] [CrossRef] [PubMed]
- Chiaramonte, F.; Anglade, P.; Baraige, F.; Langella, P.; Champomier-Vergès, M.C.; Zagorec, M. Characteristics of Lactobacillus sakei mutants selected by adaptation to the gastrointestinal tract of axenic mice. Appl. Environ. Microbiol. 2010, 76, 2932–2939. [Google Scholar] [CrossRef] [PubMed]
- Chaillou, S.; Champomier-Vergès, M.C.; Cornet, M.; Crutz Le Coq, A.M.; Dudez, A.M.; Martin, V.; Beaufils, S.; Bossy, R.; Darbon-Rongère, E.; Loux, V.; et al. Complete genome sequence of the meat-borne lactic acid bacterium Lactobacillus sakei 23K. Nat. Biotechnol. 2005, 23, 1527–1533. [Google Scholar] [CrossRef] [PubMed]
- McLeod, A.; Brede, D.A.; Rud, I.; Axelsson, L. Genome sequence of Lactobacillus sakei subsp. sakei LS25, a commercial starter culture strain for fermented sausage. Genome Announc. 2013, 1, e00475-13. [Google Scholar] [PubMed]
- Lim, H.I.; Lee, J.; Jang, J.Y.; Park, H.W.; Choi, H.J.; Kim, T.W.; Kang, M.R.; Lee, J.H. Draft genome sequence of Lactobacillus sakei strain wikim 22, isolated from kimchi in Chungcheong Province, South Korea. Genome Announc. 2014, 2, e01296-14. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, E.; Kim, C.G.; Choo, D.W.; Kim, H.Y. Draft genome sequence of Lactobacillus sakei strain FBL1, a probiotic bacterium isolated from mukeunji, a long-fermented kimchi, in South Korea. Genome Announc. 2016, 4, e00365-16. [Google Scholar] [CrossRef] [PubMed]
- McLeod, A.; Nyquist, O.L.; Snipen, L.; Naterstad, K.; Axelsson, L. Diversity of Lactobacillus sakei strains investigated by phenotypic and genotypic methods. Syst. Appl. Microbiol. 2008, 31, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Chaillou, S.; Daty, M.; Baraige, F.; Dudez, A.M.; Anglade, P.; Jones, R.; Alpert, C.-A.; Champomier-Vergès, M.C.; Zagorec, M. Intra-species genomic diversity and natural population structure of the meat-borne lactic acid bacterium Lactobacillus sakei. Appl. Environ. Microbiol. 2009, 75, 970–980. [Google Scholar] [CrossRef] [PubMed]
- Nyquist, O.L.; McLeod, A.; Brede, D.A.; Snipen, L.; Aakra, A.; Nes, I.F. Comparative genomics of Lactobacillus sakei with emphasis on strains from meat. Mol. Genet. Genom. 2011, 285, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Chaillou, S.; Lucquin, I.; Najjari, A.; Zagorec, M.; Champomier-Vergès, M.C. Population genetics of Lactobacillus sakei reveals three lineages with distinct evolutionary histories. PLoS ONE 2013, 8, e73253. [Google Scholar] [CrossRef] [PubMed]
- Lauret, R.; Morel-Deville, F.; Berthier, F.; Champomier-Vergès, M.; Postma, P.; Ehrlich, S.D.; Zagorec, M. Carbohydrate utilization in Lactobacillus sake. Appl. Environ. Microbiol. 1996, 62, 1922–1927. [Google Scholar] [PubMed]
- Xu, H.Q.; Gao, L.; Jiang, Y.S.; Tian, Y.; Peng, J.; Xa, Q.Q.; Chen, Y. Transcriptome response of Lactobacillus sakei to meat protein environment. J. Basic Microbiol. 2015, 55, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Fadda, S.; Anglade, P.; Baraige, F.; Zagorec, M.; Talon, R.; Vignolo, G.; Champomier-Vergès, M.C. Adaptive response of Lactobacillus sakei 23K during growth in the presence of meat extracts: A proteomic approach. Int. J. Food Microbiol. 2010, 142, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Aristoy, M.C.; Toldrá, F. Concentration of free amino acids and dipeptides in porcine skeletal muscles with different oxidative patterns. Meat Sci. 1998, 50, 327–332. [Google Scholar] [CrossRef]
- Zuñiga, M.; Champomier-Vergès, M.C.; Zagorec, M.; Pérez-Martinez, G. Structural and functional analysis of the gene cluster encoding the enzymes of the arginine deiminase pathway of Lactobacillus sake. J. Bacteriol. 1998, 180, 4154–4159. [Google Scholar] [PubMed]
- Zúñiga, M.; Miralles, M.C.; Pérez-Martínez, G. The product of arcR, the sixth gene of the arc operon of Lactobacillus sakei, is essential for expression of the arginine deiminase pathway. Appl. Environ. Microbiol. 2002, 68, 6051–6058. [Google Scholar] [CrossRef] [PubMed]
- Rimaux, T.; Rivière, A.; Hebert, E.M.; Mozzi, F.; Weckx, S.; De Vuyst, L.; Leroy, F. A putative transport protein is involved in citrulline excretion and re-uptake during arginine deiminase pathway activity by Lactobacillus sakei. Res. Microbiol. 2013, 164, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Rimaux, T.; Vrancken, G.; Pothakos, V.; Maes, D.; De Vuyst, L.; Leroy, F. The kinetics of the arginine deiminase pathway in the meat starter culture Lactobacillus sakei CTC 494 are pH-dependent. Food Microbiol. 2011, 28, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Rimaux, T.; Riviere, A.; Illeghems, K.; Weckx, S.; De Vuyst, L.; Leroy, F. Expression of the arginine deiminase pathway genes in Lactobacillus sakei is strain dependent and is affected by the environmental pH. Appl. Environ. Microbiol. 2012, 78, 4874–4883. [Google Scholar] [CrossRef] [PubMed]
- Champomier-Vergès, M.C.; Zuñiga, M.; Morel-Deville, F.; Pérez-Martinez, G.; Zagorec, M.; Ehrlich, S.D. Relationships between arginine degradation, pH and survival in Lactobacillus sakei. FEMS Microbiol. Lett. 1999, 180, 297–304. [Google Scholar] [CrossRef]
- Stentz, R.; Zagorec, M. Ribose utilization in Lactobacillus sakei: Analysis of the Regulation of the rbs Operon and Putative Involvement of a New Transporter. J. Mol. Microbiol. Biotechnol. 1999, 1, 165–173. [Google Scholar] [PubMed]
- McLeod, A.; Snipen, L.; Naterstad, K.; Axelsson, L. Global transcriptome response in Lactobacillus sakei during growth on ribose. BMC Microbiol. 2011, 11, 145. [Google Scholar] [CrossRef] [PubMed]
- Stentz, R.; Cornet, M.; Chaillou, S.; Zagorec, M. Adaptation of Lactobacillus sakei to meat: A new regulatory mechanism of ribose utilization? Le Lait 2001, 81, 131–138. [Google Scholar] [CrossRef]
- Rimaux, T.; Vrancken, G.; Vuylsteke, B.; De Vuyst, L.; Leroy, F. The pentose moiety of adenosine and inosine is an important energy source for the fermented-meat starter culture Lactobacillus sakei CTC 494. Appl. Environ. Microbiol. 2011, 77, 6539–6550. [Google Scholar] [CrossRef] [PubMed]
- Anba-Mondoloni, J.; Chaillou, S.; Zagorec, M.; Champomier-Vergès, M.C. Catabolism of N-Acetylneuraminic acid, as a fitness function for the food-borne lactic acid bacterium Lactobacillus sakei, involves two new proteins. Appl. Environ. Microbiol. 2013, 79, 2012–2018. [Google Scholar] [CrossRef] [PubMed]
- Garcıa-Garcıa, M.I.; Gil-Ortiz, F.; Garcıa-Carmona, F.; Sanchez-Ferrer, A. First functional and mutational analysis of group 3 N-Acetylneuraminate lyases from Lactobacillus antri and Lactobacillus sakei 23K. PLoS ONE 2014, 9, e96976. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Rhee, Y.K.; Gu, E.J.; Kim, D.W.; Jang, G.J.; Song, S.H.; Lee, J.I.; Kim, B.M.; Lee, H.J.; Hong, H.D.; et al. Mass-based metabolomic analysis of Lactobacillus sakei and its growth media at different growth phases. J. Microbiol. Biotechnol. 2017, 27, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Knauf, H.J.; Vogel, R.F.; Hammes, W.P. Cloning, sequence, and phenotypic expression of katA, which encodes the catalase of Lactobacillus sakei LTH677. Appl. Environ. Microbiol. 1992, 58, 832–839. [Google Scholar] [PubMed]
- Hertel, C.; Schmidt, G.; Fischer, M.; Oellers, K.; Hammes, W.P. Oxygen-dependent regulation of the expression of the catalase gene katA of Lactobacillus sakei LTH677. Appl. Environ. Microbiol. 1998, 64, 1359–1365. [Google Scholar] [PubMed]
- Duhutrel, P.; Bordat, C.; Wu, T.D.; Zagorec, M.; Guerquin-Kern, J.L.; Champomier-Vergès, M. Iron sources used by the nonpathogenic lactic acid bacterium Lactobacillus sakei revealed by electron energy loss spectroscopy and secondary ions mass spectrometry. Appl. Environ. Microbiol. 2010, 76, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Ammor, S.; Dufour, E.; Zagorec, M.; Chaillou, S.; Chevallier, I. Characterization and selection of Lactobacillus sakei strains isolated from traditional dry-sausage. Food Microbiol. 2005, 22, 529–538. [Google Scholar] [CrossRef]
- Parente, E.; Ciocia, F.; Ricciardi, A.; Zotta, T.; Felis, G.E.; Torriani, S. Diversity of stress tolerance in Lactobacillus plantarum, Lactobacillus pentosus and Lactobacillus paraplantarum: A multivariate screening study. Int. J. Food Microbiol. 2010, 144, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Guilbaud, M.; Zagorec, M.; Chaillou, S.; Champomier-Vergès, M.C. Intraspecies diversity of Lactobacillus sakei response to oxidative stress and variability of strain performance in mixed strains challenges. Food Microbiol. 2012, 29, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Zotta, T.; Parente, E.; Ricciardi, A. Aerobic metabolism in the genus Lactobacillus: Impact on stress response and potential applications in the food industry. J. Appl. Microbiol. 2017, 122, 1365–2672. [Google Scholar] [CrossRef] [PubMed]
- Marceau, A.; Zagorec, M.; Champomier-Vergès, M.C. Positive effects of growth at suboptimal temperature and high salt concentration on long-term survival of Lactobacillus sakei. Res. Microbiol. 2003, 154, 37–42. [Google Scholar] [CrossRef]
- Bonomo, M.G.; Sico, M.A.; Grieco, S.; Salzano, G. Fluorescent differential display analysis of Lactobacillus sakei strains under stress conditions. J. Microbiol. Methods 2010, 82, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Pot, B.; Felis, G.E.; De Bruyne, K.; Tsakalidou, E.; Papadimitriou, K.; Leisner, J.; Vandamme, P. The genus Lactobacillus. In Lactic Acid Bacteria: Biodiversity and Taxonomy; Holzapfel, W.P., Wood, B.J.B., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; pp. 249–353. [Google Scholar]
- Zheng, J.; Ruan, L.; Sun, M.; Gänzle, M. A genomic view of lactobacilli and pediococci demonstrates that phylogeny matches ecology and physiology. Appl. Environ. Microbiol. 2015, 81, 7233–7243. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Harris, H.M.B.; McCann, A.; Guo, C.; Argimón, S.; Zhang, W.; Yang, X.; Jeffery, I.B.; Cooney, J.C.; Kagawa, T.F.; et al. Expanding the biotechnology potential of lactobacilli through comparative genomics of 213 strains and associated genera. Nat. Commun. 2015, 6, 8322. [Google Scholar] [CrossRef] [PubMed]
- Leisner, J.J.; Pot, B.; Christensen, H.; Rusul, G.; Olsen, J.E.; Wee, B.W.; Ghazali, H.M. Identification of lactic acid bacteria from Chili Bo, a Malaysian food ingredient. Appl. Environ. Microbiol. 1999, 65, 599–605. [Google Scholar] [PubMed]
- Adékambi, T.; Drancourt, M.; Raoult, D. The rpoB gene as a tool for clinical microbiologists. Trends Microbiol. 2009, 17, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Torriani, S.; Van Reenen, G.A.; Klein, G.; Reuter, G.; Dellaglio, F.; Dicks, L.M. Lactobacillus curvatus subsp. curvatus subsp. nov. and Lactobacillus curvatus subsp. melibiosus subsp. nov. and Lactobacillus sake subsp. sake subsp. nov. and Lactobacillus sake subsp. carnosus subsp. nov., new subspecies of Lactobacillus curvatus Abo-Elnaga and Kandler 1965 and Lactobacillus sake Katagiri, Kitahara, and Fukami 1934 (Klein et al. 1996, emended descriptions), respectively. Int. J. Syst. Bacteriol. 1996, 46, 1158–1163. [Google Scholar]
- Microbial Genomes. Available online: https://www.ncbi.nlm.nih.gov/genome/microbes/ (accessed on 17 August 2017).
- Leroy, F.; Verluyten, J.; De Vuyst, L. Functional meat starter cultures for improved sausage fermentation. Int. J. Food Microbiol. 2006, 106, 270–285. [Google Scholar] [CrossRef] [PubMed]
- Talon, R.; Leroy, S.; Lebert, I.; Giammarinaro, P.; Chacornac, J.P.; Latorre-Moratalla, M.; Vidal-Carou, C.; Zanardi, E.; Conter, M.; Lebecque, A. Safety improvement and preservation of typical sensory qualities of traditional dry fermented sausages using autochthonous starter cultures. Int. J. Food Microbiol. 2008, 126, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Tamang, J.P.; Watanabe, K.; Holzapfel, W.H. Diversity of microorganisms in global fermented foods and beverages. Front. Microbiol. 2016, 7, 377. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Elegado, F.B.; Librojo-Basilio, N.T.; Mabesa, R.C.; Dizon, E.I. Isolation and characterisation of selected lactic acid bacteria for improved processing of Nem chua, a traditional fermented meat from Vietnam. Benef. Microbes 2010, 1, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Wanangkarn, A.; Liu, D.C.; Swetwiwathana, A.; Jindaprasert, A.; Phraephaisarn, C.; Chumnqoen, W.; Tan, F.J. Lactic acid bacterial population dynamics during fermentation and storage of Thai fermented sausage according to restriction fragment length polymorphism analysis. Int. J. Food Microbiol. 2014, 186, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Fontana, C.; Bassi, D.; López, C.; Pisacane, V.; Otero, M.C.; Puglisi, E.; Rebecchi, A.; Cocconcelli, P.S.; Vignolo, G. Microbial ecology involved in the ripening of naturally fermented llama meat sausages. A focus on lactobacilli diversity. Int. J. Food Microbiol. 2016, 236, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Hüfner, E.; Markieton, T.; Chaillou, S.; Crutz Le Coq, A.M.; Zagorec, M.; Hertel, C. Identification of Lactobacillus sakei genes induced in meat fermentation and their role in survival and growth. Appl. Environ. Microbiol. 2007, 73, 2522–2531. [Google Scholar] [CrossRef] [PubMed]
- Hüfner, E.; Hertel, C. Improvement of raw sausage fermentation by stress-conditioning of the starter organism Lactobacillus sakei. Curr. Microbiol. 2008, 57, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Ojha, K.S.; Kerry, J.P.; Alvarez, C.; Walsh, D.; Tiwari, B.K. Effect of high intensity ultrasound on the fermentation profile of Lactobacillus sakei in a meat model system. Ultrason. Sonochem. 2016, 31, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Tremonte, P.; Reale, A.; Di Renzo, T.; Tipaldi, L.; Di Luccia, A.; Coppola, R.; Sorrentino, E.; Succi, M. Interactions between Lactobacillus sakei and CNC (Staphylococcus xylosus and Kocuria varians) and their influence on proteolytic activity. Lett. Appl. Microbiol. 2010, 51, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Song, J.H.; Park, S.H.; Lee, J.H.; Park, H.W.; Kim, T.W. Effects of brown rice extract treated with Lactobacillus sakei Wikim001 on osteoblast differentiation and osteoclast formation. Prev. Nutr. Food Sci. 2014, 19, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Yun, B.; Moon, J.H.; Park, D.J.; Lim, K.; Oh, S. Characterization of selected Lactobacillus strains for use as probiotic. Korean J. Food Sci. Anim. Resour. 2015, 35, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.D.; Hong, Y.F.; Jeon, B.; Jung, B.J.; Chung, D.K.; Kim, H. Differential cytokine regulatory effect of three Lactobacillus strains isolated from fermented foods. J. Microbiol. Biotechnol. 2016, 26, 1517–1526. [Google Scholar] [CrossRef] [PubMed]
- Møretrø, T.; Naterstad, C.; Wang, E.; Aasen, I.; Chaillou, S.; Zagorec, M.; Axelsson, L. Sakacin P non-producing Lactobacillus sakei strains contain homologues of the sakacin P gene cluster. Res. Microbiol. 2005, 156, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Castellano, P.; Belfiore, C.; Fadda, S.; Vignolo, G. A review of bacteriocinogenic lactic acid bacteria used as bioprotective cultures in fresh meat produced in Argentina. Meat Sci. 2008, 79, 483–499. [Google Scholar] [CrossRef] [PubMed]
- Castellano, P.; Aristoy, M.C.; Sentandreu, M.A.; Vignolo, G.; Toldrá, F. Lactobacillus sakei CRL1862 improves safety and protein hydrolysis in meat systems. J. Appl. Microbiol. 2012, 113, 1407–1416. [Google Scholar] [CrossRef] [PubMed]
- Bredholt, S.; Nesbakken, T.; Holck, A. Protective cultures inhibit growth of Listeria monocytogenes and Escherichia coli O157:H7 in cooked, sliced, vacuum- and gas-packaged meat. Int. J. Food Microbiol. 1999, 53, 43–52. [Google Scholar] [CrossRef]
- Bredholt, S.; Nesbakken, T.; Holck, A. Industrial application of an antilisterial strain of Lactobacillus sakei as a protective culture and its effect on the sensory acceptability of cooked, sliced, vacuum-packaged meats. Int. J. Food Microbiol. 2001, 66, 191–196. [Google Scholar] [CrossRef]
- Vermeiren, L.; Devlieghere, F.; Debevere, J. Evaluation of meat born lactic acid bacteria as protective cultures for the biopreservation of cooked meat products. Int. J. Food Microbiol. 2004, 96, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Vermeiren, L.; Devlieghere, F.; Debevere, J. Co-culture experiments demonstrate the usefulness of Lactobacillus sakei 10A to prolong the shelf-life of a model cooked ham. Int. J. Food Microbiol. 2006, 108, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Vermeiren, L.; Devlieghere, F.; Vandekinderen, I.; Debevere, J. The interaction of the non-bacteriocinogenic Lactobacillus sakei 10A and lactocin S producing Lactobacillus sakei 148 towards Listeria monocytogenes on a model cooked ham. Food Microbiol. 2006, 23, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Vermeiren, L.; Devlieghere, F.; Vandekinderen, I.; Rajtak, U.; Debevere, J. The sensory acceptability of cooked meat products treated with a protective culture depends on glucose content and buffering capacity: A case study with Lactobacillus sakei 10A. Meat Sci. 2006, 74, 532–545. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.J.; Hussein, H.M.; Zagorec, M.; Brightwell, G.; Tagg, J.R. Isolation of lactic-acid bacteria with inhibitory activity against pathogens and spoilage organisms associated with fresh meat. Food Microbiol. 2008, 25, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.J.; Zagorec, M.; Tagg, J.R. Inhibition by Lactobacillus sakei of other species in the flora of vacuum packaged raw meats during prolonged storage. Food Microbiol. 2009, 26, 876–881. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.J.; Wilklund, E.; Zagorec, M.; Tagg, J.R. Evaluation of stored lamb bio-preserved using a three-strain cocktail of Lactobacillus sakei. Meat Sci. 2010, 86, 955–959. [Google Scholar] [CrossRef] [PubMed]
- Chaillou, S.; Christieans, S.; Rivollier, M.; Lucquin, I.; Champomier-Vergès, M.C.; Zagorec, M. Quantification and efficiency of lactic acid bacteria used as bioprotective cultures in ground beef. Meat Sci. 2014, 97, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Xu, X.L.; Zhou, G.H.; Han, Y.Q.; Xu, B.C.; Liu, J.C. Study of the Lactobacillus sakei protective effect towards spoilage bacteria in vacuum packed cooked ham analyzed by PCR-DGGE. Meat Sci. 2008, 80, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Comi, G.; Andyanto, D.; Manzano, M.; Iacumin, L. Lactococcus lactis and Lactobacillus sakei as bio-protective culture to eliminate Leuconostoc mesenteroides spoilage and improve the shelf life and sensorial characteristics of commercial cooked bacon. Food Microbiol. 2016, 58, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Huh, C.K.; Hwang, T.Y. Identification of antifungal substances of Lactobacillus sakei subsp. ALI033 and antifungal cctivity against Penicillium brevicompactum Strain FI02. Prev. Nutr. Food Sci. 2016, 21, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Zavaleta, O.; López-Malo, A.; Hernández-Mendoza, A.; García, H.S. Antifungal activity of lactobacilli and its relationship with 3-phenyllactic acid production. Int. J. Food Microbiol. 2014, 173, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Doulgeraki, A.I.; Ercolini, D.; Villani, F.; Nychas, G.J. Spoilage microbiota associated to the storage of raw meat in different conditions. Int. J. Food Microbiol. 2012, 157, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Schirmer, B.C.; Heir, E.; Langsrud, S. Characterization of the bacterial spoilage flora in marinated pork products. J. Appl. Microbiol. 2009, 106, 2106–2116. [Google Scholar] [CrossRef] [PubMed]
- Ammor, M.S.; Gueimonde, M.; Danielsen, M.; Zagorec, M.; van Hoeck, A.H.A.M.; de los Reyes-Gavilan, C.G.; Mayo, B.; Margolles, A. Two different tetracycline resistance mechanisms, plasmid-carried tet (L) and chromosomally-encoded transposon-associated tet (M), coexist in Lactobacillus sakei Tits 9. Appl. Environ. Microbiol. 2008, 74, 1394–1401. [Google Scholar] [CrossRef] [PubMed]
- Gevers, D.; Danielsen, M.; Huys, G.; Swings, J. Molecular characterization of tet (M) genes in Lactobacillus isolates from different types of fermented dry sausage. Appl. Environ. Microbiol. 2003, 69, 1270–1275. [Google Scholar] [CrossRef] [PubMed]
- Zonenschain, D.; Rebecchi, A.; Morelli, L. Erythromycin-and tetracycline-resistant lactobacilli in Italian fermented dry sausages. J. Appl. Microbiol. 2009, 107, 1559–1568. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Tsai, C.Y.; Lin, C.F.; Wang, Y.C.; Wang, I.K.; Chung, T.C. Characterization of tetracycline resistance lactobacilli isolated from swine intestines at western area of Taiwan. Anaerobe 2011, 17, 239–245. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zagorec, M.; Champomier-Vergès, M.-C. Lactobacillus sakei: A Starter for Sausage Fermentation, a Protective Culture for Meat Products. Microorganisms 2017, 5, 56. https://doi.org/10.3390/microorganisms5030056
Zagorec M, Champomier-Vergès M-C. Lactobacillus sakei: A Starter for Sausage Fermentation, a Protective Culture for Meat Products. Microorganisms. 2017; 5(3):56. https://doi.org/10.3390/microorganisms5030056
Chicago/Turabian StyleZagorec, Monique, and Marie-Christine Champomier-Vergès. 2017. "Lactobacillus sakei: A Starter for Sausage Fermentation, a Protective Culture for Meat Products" Microorganisms 5, no. 3: 56. https://doi.org/10.3390/microorganisms5030056
APA StyleZagorec, M., & Champomier-Vergès, M.-C. (2017). Lactobacillus sakei: A Starter for Sausage Fermentation, a Protective Culture for Meat Products. Microorganisms, 5(3), 56. https://doi.org/10.3390/microorganisms5030056