Enzymatic Preparation of 2,5-Furandicarboxylic Acid (FDCA)—A Substitute of Terephthalic Acid—By the Joined Action of Three Fungal Enzymes


Abstract
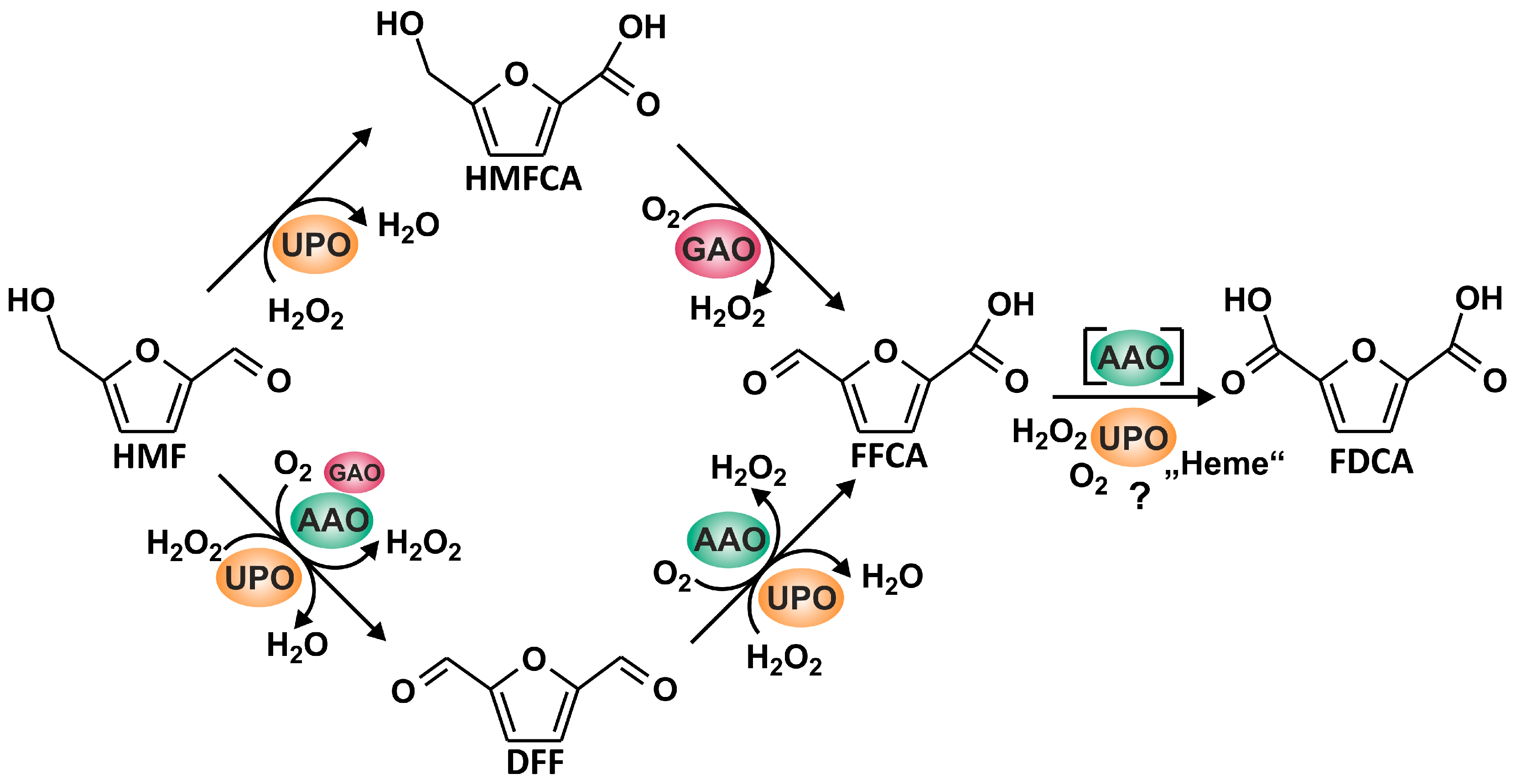
:1. Introduction
2. Materials and Methods
2.1. Enzyme Preparations
2.2. Chemicals
2.3. HPLC Analyses
2.4. Reaction Setups
3. Results
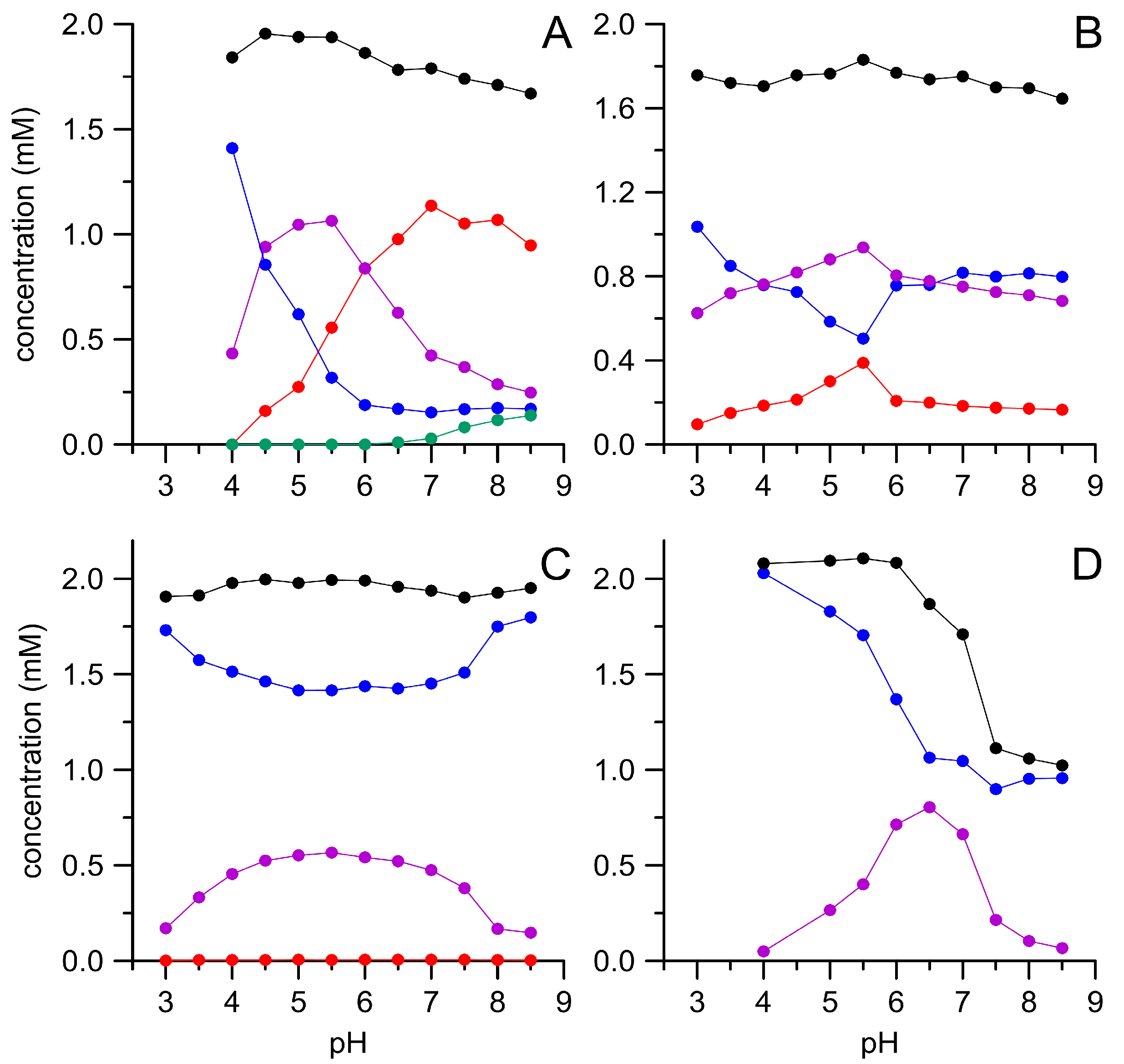
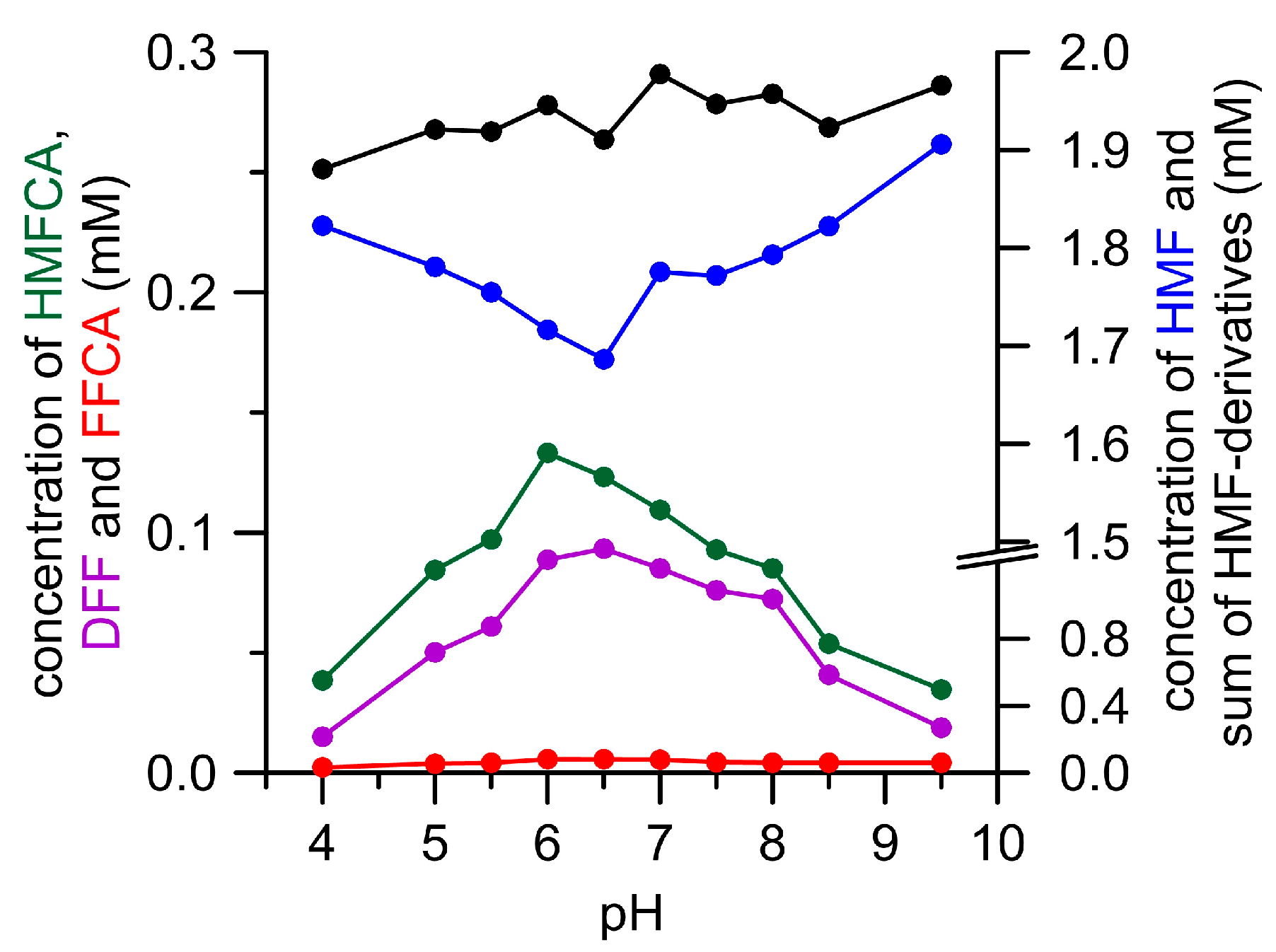
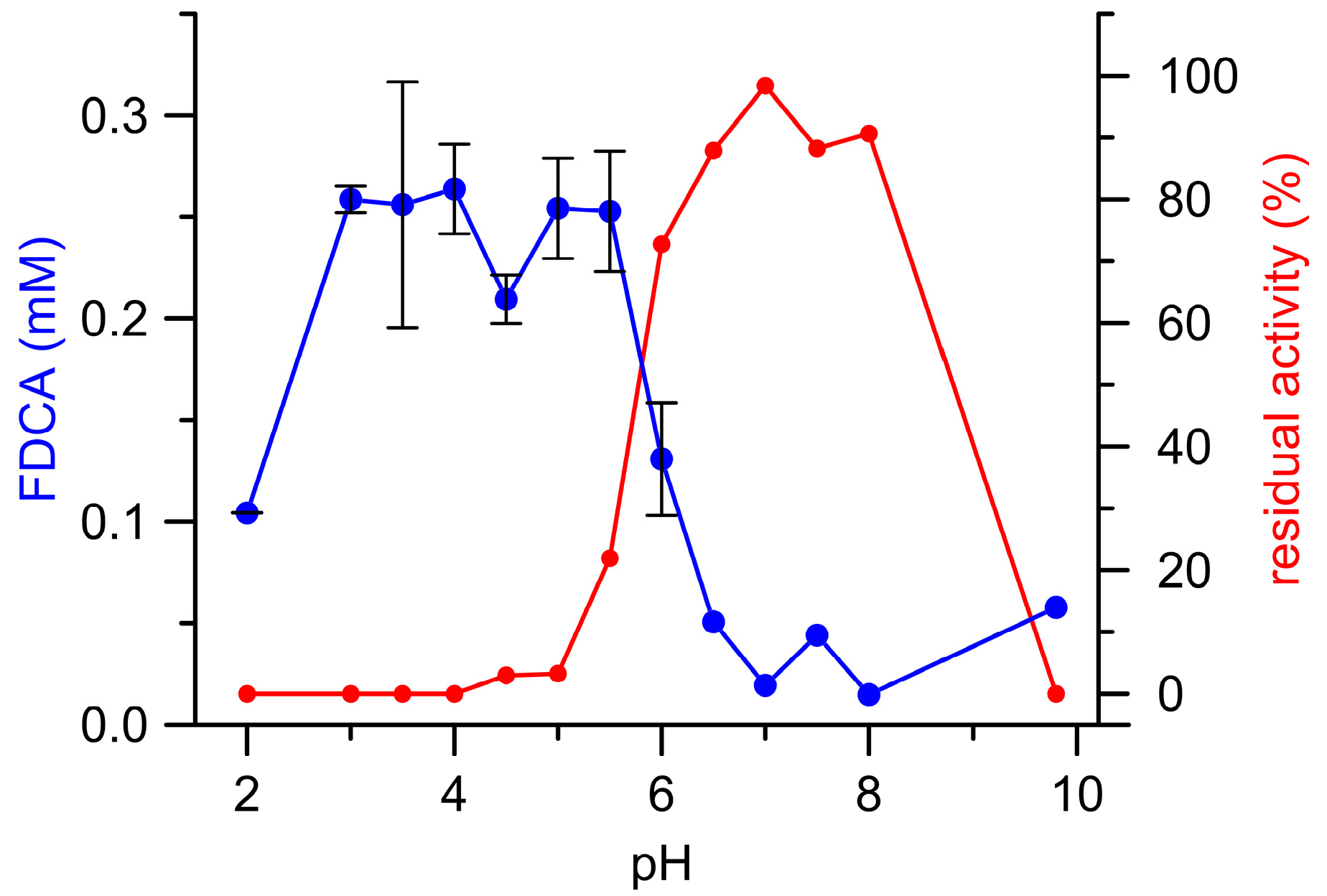
3.1. Oxidation of HMF and Its Derivatives
3.2. FFCA Oxidation
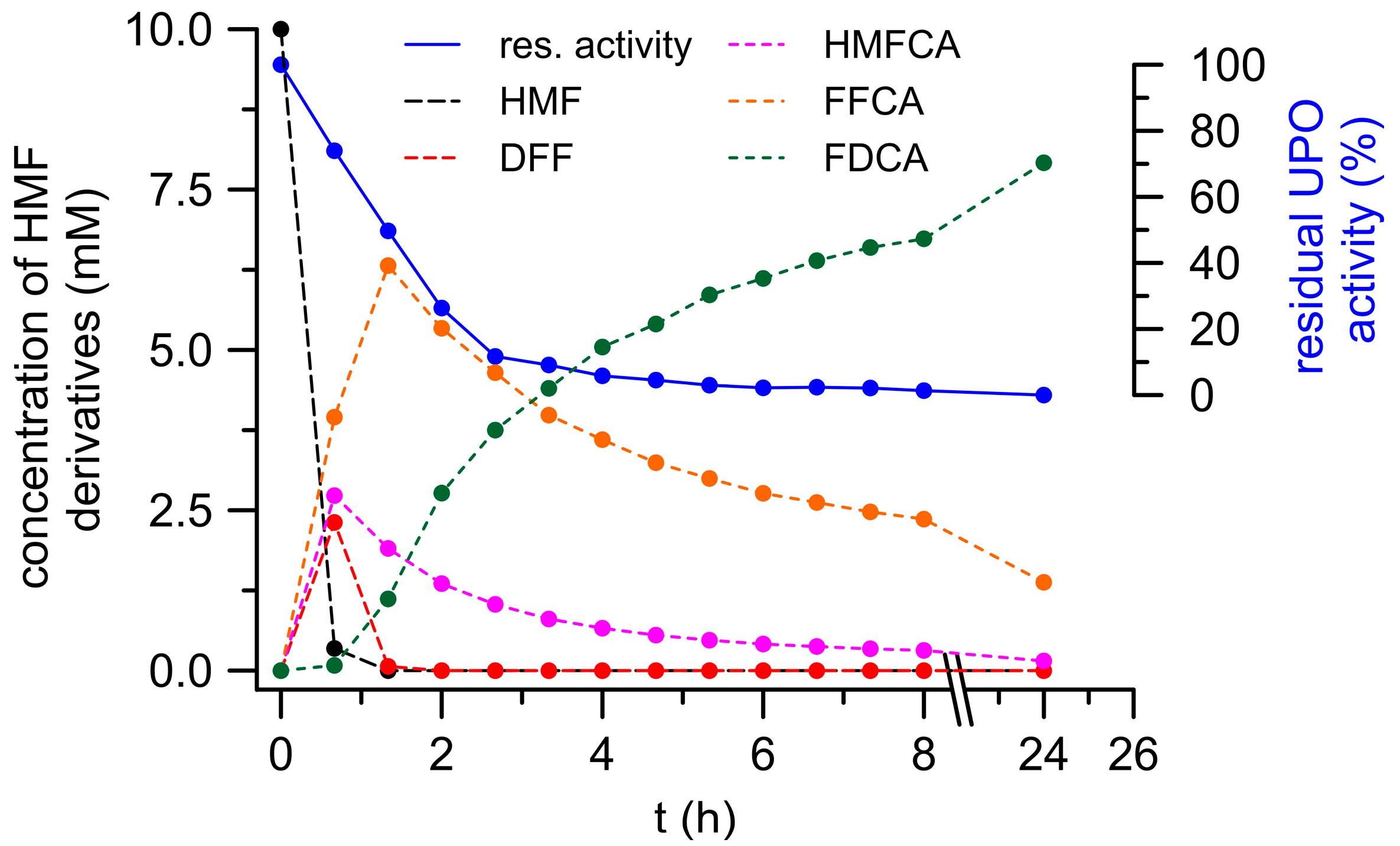
3.3. Combined Cascade Reaction
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Siddique, R.; Khatib, J.; Kaur, I. Use of recycled plastic in concrete: A review. Waste Manag. 2008, 28, 1835–1852. [Google Scholar] [CrossRef] [PubMed]
- Global Polyethylene Terephthalate Production 2014–2020. Available online: https://www.statista.com/statistics/650191/global-polyethylene-terephthalate-production-outlook/ (accessed on 30 November 2017).
- Eriksson, O.; Finnveden, G. Plastic waste as a fuel—CO2-neutral or not? Energy Environ. Sci. 2009, 2, 907–914. [Google Scholar] [CrossRef]
- Papageorgiou, G.Z.; Tsanaktsis, V.; Bikiaris, D.N. Synthesis of poly(ethylene furandicarboxylate) polyester using monomers derived from renewable resources: Thermal behavior comparison with PET and PEN. Phys. Chem. Chem. Phys. 2014, 16, 7946–7958. [Google Scholar] [CrossRef] [PubMed]
- Rosatella, A.A.; Simeonov, S.P.; Frade, R.F.M.; Afonso, C.A.M. 5-hydroxymethylfurfural (HMF) as a building block platform: Biological properties, synthesis and synthetic applications. Green Chem. 2011, 13, 754–793. [Google Scholar] [CrossRef]
- Wierckx, N.; Elink Schuurman, T.D.; Blank, L.M.; Ruijssenaars, H.J. Whole-cell biocatalytic production of 2,5-furandicarboxylic acid. In Microorganisms in Biorefineries; Kamm, B., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 207–223. [Google Scholar]
- Koopman, F.; Wierckx, N.; de Winde, J.H.; Ruijssenaars, H.J. Efficient whole-cell biotransformation of 5-(hydroxymethyl)furfural into FDCA, 2,5-furandicarboxylic acid. Bioresour. Technol. 2010, 101, 6291–6296. [Google Scholar] [CrossRef] [PubMed]
- Dijkman, W.P.; Groothuis, D.E.; Fraaije, M.W. Enzyme-catalyzed oxidation of 5-hydroxymethylfurfural to furan-2,5-dicarboxylic acid. Angew. Chem. Int. Ed. 2014, 53, 6515–6518. [Google Scholar] [CrossRef] [PubMed]
- McKenna, S.M.; Leimkuhler, S.; Herter, S.; Turner, N.J.; Carnell, A.J. Enzyme cascade reactions: Synthesis of furandicarboxylic acid (FDCA) and carboxylic acids using oxidases in tandem. Green Chem. 2015, 17, 3271–3275. [Google Scholar] [CrossRef]
- Carro, J.; Ferreira, P.; Rodríguez, L.; Prieto, A.; Serrano, A.; Balcells, B.; Ardá, A.; Jiménez-Barbero, J.; Gutiérrez, A.; Ullrich, R.; et al. 5-hydroxymethylfurfural conversion by fungal aryl-alcohol oxidase and unspecific peroxygenase. FEBS J. 2015, 282, 3218–3229. [Google Scholar] [CrossRef] [PubMed]
- Van Deurzen, M.P.J.; van Rantwijk, F.; Sheldon, R.A. Chloroperoxidase-catalyzed oxidation of 5-hydroxymethylfurfural. J. Carbohydr. Chem. 1997, 16, 299–309. [Google Scholar] [CrossRef]
- Whittaker, J.W. Galactose oxidase. In Advances in Protein Chemistry; Academic Press: Cambridge, MA, USA, 2002; Volume 60, pp. 1–49. [Google Scholar]
- Sigoillot, C.; Camarero, S.; Vidal, T.; Record, E.; Asther, M.; Pérez-Boada, M.; Martínez, M.J.; Sigoillot, J.-C.; Asther, M.; Colom, J.F.; et al. Comparison of different fungal enzymes for bleaching high-quality paper pulps. J. Biotechnol. 2005, 115, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Hofrichter, M.; Kellner, H.; Pecyna, M.; Ullrich, R. Fungal unspecific peroxygenases: Heme-thiolate proteins that combine peroxidase and cytochrome p450 properties. In Monooxygenase, Peroxidase and Peroxygenase Properties and Mechanisms of Cytochrome p450; Hrycay, E.G., Bandiera, S.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; Volume 851, pp. 341–368. [Google Scholar]
- Reynolds, M.P.; Baron, A.J.; Wilmot, C.M.; Vinecombe, E.; Stevens, C.; Phillips, S.E.V.; Knowles, P.F.; McPherson, M.J. Structure and mechanism of galactose oxidase: Catalytic role of tyrosine 495. JBIC J. Biol. Inorg. Chem. 1997, 2, 327–335. [Google Scholar] [CrossRef]
- Correia, M.A.S.; Otrelo-Cardoso, A.R.; Schwuchow, V.; Sigfridsson Clauss, K.G.V.; Haumann, M.; Romão, M.J.; Leimkühler, S.; Santos-Silva, T. The Escherichia coli periplasmic aldehyde oxidoreductase is an exceptional member of the xanthine oxidase family of molybdoenzymes. ACS Chem. Biol. 2016, 11, 2923–2935. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, R.; Nüske, J.; Scheibner, K.; Spantzel, J.; Hofrichter, M. Novel haloperoxidase from the agaric basidiomycete Agrocybe aegerita oxidizes aryl alcohols and aldehydes. Appl. Environ. Microbiol. 2004, 70, 4575–4581. [Google Scholar] [CrossRef] [PubMed]
- Kirk, T.K.; Farrell, R.L. Enzymatic “combustion”: The microbial degradation of lignin. Annu. Rev. Microbiol. 1987, 41, 465–501. [Google Scholar] [CrossRef] [PubMed]
- Feldman, D.; Kowbel, D.J.; Glass, N.L.; Yarden, O.; Hadar, Y. Detoxification of 5-hydroxymethylfurfural by the Pleurotus ostreatus lignolytic enzymes aryl alcohol oxidase and dehydrogenase. Biotechnol. Biofuels 2015, 8, 63. [Google Scholar] [CrossRef] [PubMed]
- Kalum, L.; Morant, M.D.; Lund, H.; Jensen, J.; Lapainaite, I.; Soerensen, N.H.; Pedersen, S.; Østergaard, L.H.; Xu, F. Enzymatic Oxidation of 5-hydroxymethylfurfural and Derivatives Thereof. U.S. Patent WO2014-015256A2, 23 January 2014. [Google Scholar]
- Riley, R.; Salamov, A.A.; Brown, D.W.; Nagy, L.G.; Floudas, D.; Held, B.W.; Levasseur, A.; Lombard, V.; Morin, E.; Otillar, R.; et al. Extensive sampling of basidiomycete genomes demonstrates inadequacy of the white-rot/brown-rot paradigm for wood decay fungi. Proc. Natl. Acad. Sci. USA 2014, 111, 9923–9928. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, I.S.; Ruiz-Duenas, F.J.; Santillana, E.; Ferreira, P.; Martinez, M.J.; Martinez, A.T.; Romero, A. Novel structural features in the gmc family of oxidoreductases revealed by the crystal structure of fungal aryl-alcohol oxidase. Acta Crystallogr. Sect. D 2009, 65, 1196–1205. [Google Scholar] [CrossRef] [PubMed]
- Vonck, J.; Parcej, D.N.; Mills, D.J. Structure of alcohol oxidase from Pichia pastoris by cryo-electron microscopy. PLoS ONE 2016, 11, e0159476. [Google Scholar] [CrossRef] [PubMed]
- Tanner, W.; Lehle, L. Protein glycosylation in yeast. Biochim. Biophys. Acta (BBA) Rev. Biomembr. 1987, 906, 81–99. [Google Scholar] [CrossRef]
- Whittaker, M.M.; Whittaker, J.W. Expression of recombinant galactose oxidase by Pichia pastoris. Protein Expr. Purif. 2000, 20, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Karich, A.; Scheibner, K.; Ullrich, R.; Hofrichter, M. Exploring the catalase activity of unspecific peroxygenases and the mechanism of peroxide-dependent heme destruction. J. Mol. Catal. B Enzym. 2016, 134, 238–246. [Google Scholar] [CrossRef]
- Kalko, S.G.; Gelpí, J.L.; Fita, I.; Orozco, M. Theoretical study of the mechanisms of substrate recognition by catalase. J. Am. Chem. Soc. 2001, 123, 9665–9672. [Google Scholar] [CrossRef] [PubMed]
- Hofrichter, M.; Ziegenhagen, D.; Vares, T.; Friedrich, M.; Jäger, M.G.; Fritsche, W.; Hatakka, A. Oxidative decomposition of malonic acid as basis for the action of manganese peroxidase in the absence of hydrogen peroxide. FEBS Lett. 1998, 434, 362–366. [Google Scholar] [CrossRef]
- Urzúa, U.; Kersten, P.J.; Vicuña, R. Manganese peroxidase-dependent oxidation of glyoxylic and oxalic acids synthesized by Ceriporiopsis subvermispora produces extracellular hydrogen peroxide. Appl. Environ. Microbiol. 1998, 64, 68–73. [Google Scholar] [PubMed]
- Ortiz de Montellano, P.R.; David, S.K.; Ator, M.A.; Tew, D. Mechanism-based inactivation of horseradish peroxidase by sodium azide. Formation of meso-azidoprotoporphyrin ix. Biochemistry 1988, 27, 5470–5476. [Google Scholar] [PubMed]
- Suzuki, Y.; Kawarada, A. Products of peroxidase catalyzed oxidation of indolyl-3-acetic acid. Agric. Biol. Chem. 1978, 42, 1315–1321. [Google Scholar]
- Gazaryan, I.G.; Lagrimini, L.M.; Ashby, G.A.; Thorneley, R.N. Mechanism of indole-3-acetic acid oxidation by plant peroxidases: Anaerobic stopped-flow spectrophotometric studies on horseradish and tobacco peroxidases. Biochem. J. 1996, 313, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Zorn, H.; Langhoff, S.; Scheibner, M.; Berger, R.G. Cleavage of β,β-carotene to flavor compounds by fungi. Appl. Microbiol. Biotechnol. 2003, 62, 331–336. [Google Scholar] [CrossRef] [PubMed]
- McKenna, S.M.; Mines, P.; Law, P.; Kovacs-Schreiner, K.; Birmingham, W.R.; Turner, N.J.; Leimkuhler, S.; Carnell, A.J. The continuous oxidation of HMF to FDCA and the immobilisation and stabilisation of periplasmic aldehyde oxidase (PaoABC). Green Chem. 2017, 19, 4660–4665. [Google Scholar] [CrossRef]
- Mayer, S.F.; Kroutil, W.; Faber, K. Enzyme-initiated domino (cascade) reactions. Chem. Soc. Rev. 2001, 30, 332–339. [Google Scholar] [CrossRef]
- Ricca, E.; Brucher, B.; Schrittwieser, J.H. Multi-enzymatic cascade reactions: Overview and perspectives. Adv. Synth. Catal. 2011, 353, 2239–2262. [Google Scholar] [CrossRef]
- Gibson, Q.H.; Swoboda, B.E.P.; Massey, V. Kinetics and mechanism of action of glucose oxidase. J. Biol. Chem. 1964, 239, 3927–3934. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Substrate | KM (mM) | kcat (min−1) | kcat/KM (M−1 s−1) |
|---|---|---|---|---|
| AaeUPO | HMF | 6.07 | 13,333 | 36,610 |
| DFF | 0.82 | 1752 | 35,622 | |
| PeryAAO | HMF | 36.3 | 219 | 100 |
| PostAAO | HMF | 7.2 | 177 | 411 |
| GAO | HMF | 142 | 42 | 4.9 |
| Reaction Setup | FDCA (mM) |
|---|---|
| AaeUPO | 0.31 ± 0.01 |
| AaeUPO/H2O2 | 0.40 ± 0.05 |
| H2O2 | 0.05 ± 0.01 |
| Hemoglobin/H2O2 | 0.06 ± 0.00 |
| Hemin/H2O2 | 0.06 ± 0.00 |
| AaeUPO/Cat | 1.88 ± 0.11 |
| AaeUPO/SOD | 0.33 ± 0.02 |
| AaeUPO/Cat/SOD | 1.98 ± 0.09 |
| AaeUPO/Cat/SOD/H2O2 | 2.08 ± 0.07 |
| Cat | 1.39 ± 0.09 |
| Cat/SOD | 1.37 ± 0.08 |
| AaeUPO/Cat/GOD | 0.61 ± 0.03 |
| Cat/GOD | 0.08 ± 0.01 |
| HRP | 0.52 ± 0.01 |
| UPO/sodium azide | 0.00 ± 0.00 |
| UPO (boiled) | 0.00 ± 0.00 |
| Cat/sodium azide | 0.00 ± 0.00 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karich, A.; Kleeberg, S.B.; Ullrich, R.; Hofrichter, M. Enzymatic Preparation of 2,5-Furandicarboxylic Acid (FDCA)—A Substitute of Terephthalic Acid—By the Joined Action of Three Fungal Enzymes. Microorganisms 2018, 6, 5. https://doi.org/10.3390/microorganisms6010005
Karich A, Kleeberg SB, Ullrich R, Hofrichter M. Enzymatic Preparation of 2,5-Furandicarboxylic Acid (FDCA)—A Substitute of Terephthalic Acid—By the Joined Action of Three Fungal Enzymes. Microorganisms. 2018; 6(1):5. https://doi.org/10.3390/microorganisms6010005
Chicago/Turabian StyleKarich, Alexander, Sebastian B. Kleeberg, René Ullrich, and Martin Hofrichter. 2018. "Enzymatic Preparation of 2,5-Furandicarboxylic Acid (FDCA)—A Substitute of Terephthalic Acid—By the Joined Action of Three Fungal Enzymes" Microorganisms 6, no. 1: 5. https://doi.org/10.3390/microorganisms6010005
APA StyleKarich, A., Kleeberg, S. B., Ullrich, R., & Hofrichter, M. (2018). Enzymatic Preparation of 2,5-Furandicarboxylic Acid (FDCA)—A Substitute of Terephthalic Acid—By the Joined Action of Three Fungal Enzymes. Microorganisms, 6(1), 5. https://doi.org/10.3390/microorganisms6010005