Green Technology: Bacteria-Based Approach Could Lead to Unsuspected Microbe–Plant–Animal Interactions



Abstract
:1. Introduction
2. Plant Microbiome: A Fascinating Source of BCAs
3. Human and Animal Pathogens Associated with Plants: Simple Contamination or Survival/Spreading Strategies
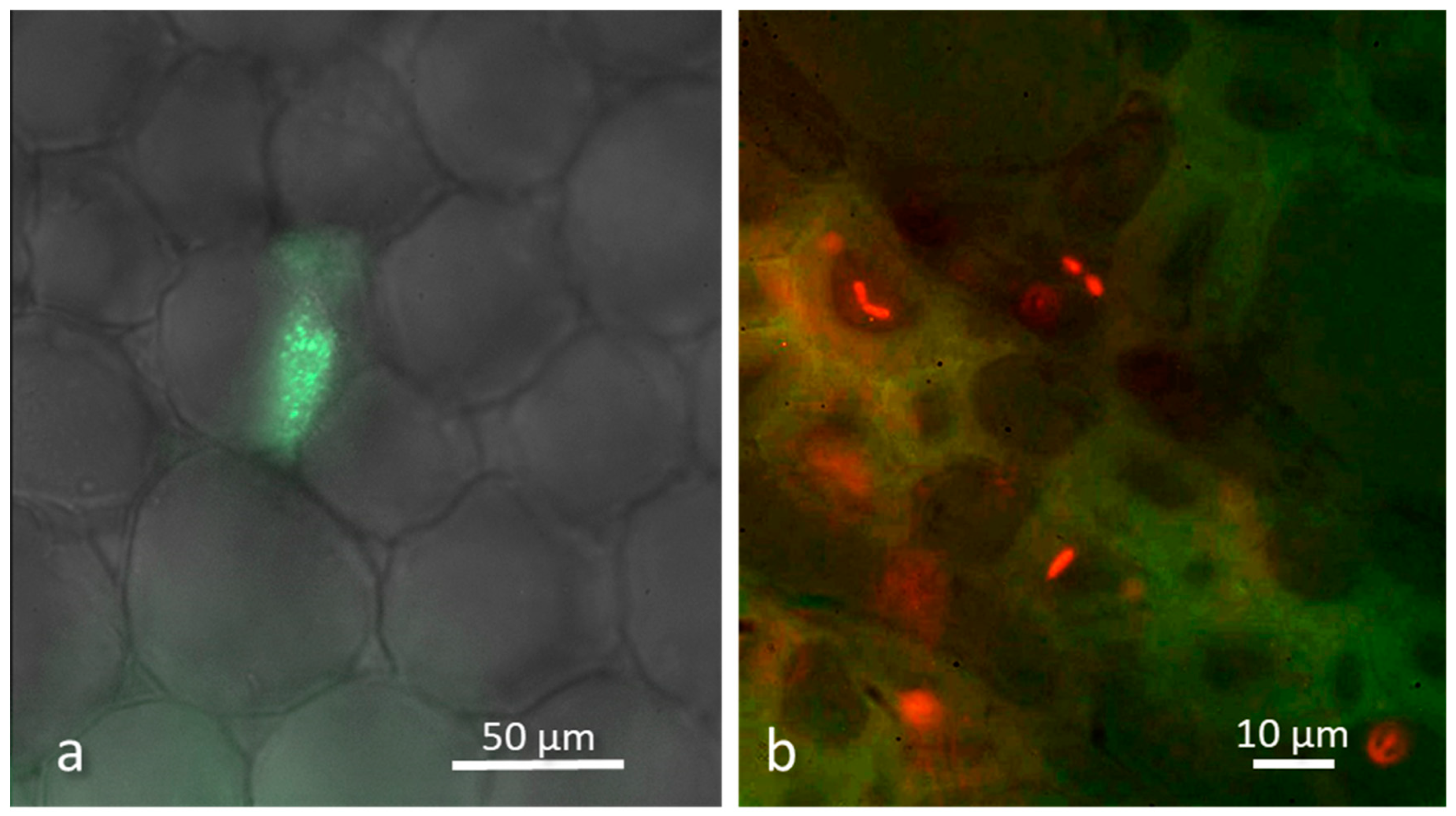
4. Bacteria Can Overcome Kingdom Barriers
5. Possible Unfavorable Consequences of the Massive Use of Bacteria as Biocontrol or Biofertilizer Agents
6. Conclusions
Funding
Conflicts of Interest
References
- Flint, M.L.; Dreistad, S.H. Natural Enemies Handbook: The Illustrated Guide to Biological Pest Control; University of California Press: Oakland, CA, USA, 1998. [Google Scholar]
- Francis, M.; Holsters, M.; Vereecke, D. The Gram-positive side of plant-microbe interactions. Environ. Microbiol. 2009, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [PubMed]
- Glare, T.; Caradus, J.; Gelernter, W.; Jackson, T.; Keyhani, N.; Köhl, J.; Marrone, P.; Reardon, R.C.; Podgwaite, J.; Zerillo, R. Have biopesticides come of age? Trends Biotechnol. 2012, 30, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Martínez-Viveros, O.; Jorquera, M.A.; Crowley, D.E.; Gajardo, G.; Mora, M.L. Mechanisms and practical considerations involved in plant growth promotion by rhizobacteria. J. Soil Sci. Plant Nutr. 2010, 10, 293–319. [Google Scholar] [CrossRef]
- Pirofski, L.A.; Casadevall, A. Q&A: What is a pathogen? A question that begs the point. BMC Biol. 2012, 10, 6. [Google Scholar] [CrossRef]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The Hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed]
- Rout, M.E. The Plant Microbiome. In Advances in Botanical Research; Paterson, A.H., Ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2014; Volume 69, pp. 280–302. [Google Scholar]
- Chow, J.; Tang, H.; Mazmanian, S.K. Pathobionts of the gastrointestinal microbiota and inflammatory disease. Curr. Opin. Immunol. 2011, 23, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Vayssier-Taussat, M.; Albina, E.; Citti, C.; Cosson, J.F.; Jacques, M.-A.; Lebrun, M.-H.; Le Loir, Y.; Ogliastro, M.; Petit, M.A.; Roumagnac, P.; et al. Shifting the paradigm from pathogens to pathobiome: New concepts in the light of meta-omics. Front. Cell. Infect. Microbiol. 2014, 4, 29. [Google Scholar] [CrossRef] [PubMed]
- Van der Riet, F.D. Diseases of plants transmissible between plants and man (phytonoses) exist. Med. Hypotheses 1997, 49, 359–361. [Google Scholar]
- Van Overbeek, L.S.; van Doorn, J.; Wicher, J.H.; Amerongen, A.; van Roermund, H.J.W.; Willemsen, P.T.J. The arable ecosystem as battle ground for emergence of new human pathogens. Front. Microbiol. 2014, 4, 104. [Google Scholar] [CrossRef]
- Bais, H.P.; Park, S.W.; Weir, T.L.; Callaway, R.M.; Vivanco, J.M. How plants communicate using the underground information superhighway. Trends Plant. Sci. 2004, 9, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.G. Can microbiome studies lead to effective biocontrol of plant disease? Mol. Plant Microbe Interact. 2017, 30, 190–193. [Google Scholar] [CrossRef] [PubMed]
- Bashan, Y.; de-Bashan, L.E.; Prabhu, S.R.; Hernandez, J.P. Advances in plant growth-promoting bacterial inoculant technology: Formulations and practical perspectives (1998–2013). Plant Soil 2014, 378, 1–33. [Google Scholar] [CrossRef]
- O’Brien, P.A. Biological control of plant diseases. Australas. Plant Pathol. 2017, 46, 293–304. [Google Scholar] [CrossRef]
- Bernal, P.; Allsopp, L.P.; Filloux, A.; Llamas, M.A. The Pseudomonas putida T6SS is a plant warden against phytopathogens. ISME J. 2017, 11, 972–987. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Bullen, N.P.; Ahmad, S.; Whitney, J.C. Diverse NADase effector families mediate interbacterial antagonism via the type VI secretion system. J. Biol. Chem. 2018, 293, 1504–1514. [Google Scholar] [CrossRef]
- Vettiger, A.; Basler, M. Type VI secretion system substrates are transferred and reused among sister cells. Cell 2016, 167, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Ho, B.T.; Fu, Y.; Dong, T.G.; Mekalanos, J.J. Vibrio cholerae type 6 secretion system effector trafficking in target bacterial cells. Proc. Natl. Acad. Sci. USA 2017, 114, 9427–9432. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.J.; Kuo, T.Y.; Hsieh, F.C.; Chen, P.Y.; Wang, C.S.; Shih, Y.L.; Lai, Y.M.; Liu, J.R.; Yang, Y.L.; Shih, M.C. Involvement of type VI secretion system in secretion of iron chelator pyoverdine in Pseudomonas taiwanensis. Sci Rep. 2016, 8, 32950. [Google Scholar] [CrossRef] [PubMed]
- Milner, J.L.; Silo-Suh, L.; Lee, J.C.; He, H.Y.; Clardy, J.; Handelsman, J. Production of kanosamine by Bacillus cereus UW85. Appl. Environ. Microbiol. 1996, 62, 3061–3065. [Google Scholar] [PubMed]
- Nakayama, T.; Homma, Y.; Hashidoko, Y.; Mizutani, J.; Tahara, S. Possible role of xanthobaccins produced by Stenotrophomonas sp. strain SB-K88 in suppression of sugar beet damping-off disease. Appl. Environ. Microbiol. 1999, 65, 4334–4339. [Google Scholar] [PubMed]
- Raaijmakers, J.M.; Vlami, M.; de Souza, J.T. Antibiotic production by bacterial biocontrol agents. Antonie Van Leeuwenhoek 2002, 81, 537. [Google Scholar] [CrossRef] [PubMed]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol Microbiol. 2005, 56, 845–885. [Google Scholar] [CrossRef] [PubMed]
- Grady, E.N.; MacDonald, J.; Liu, L.; Richman, A.; Yuan, Z.-C. Current knowledge and perspectives of Paenibacillus: A review. Microb. Cell Fact. 2016, 15, 203. [Google Scholar] [CrossRef] [PubMed]
- Compant, S.; Duffy, B.; Nowak, J.; Clement, C.; Barka, E.A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef]
- Tyc, O.; van den Berg, M.; Gerards, S.; van Veen, J.A.; Raaijmakers, J.M.; de Boer, W.; Garbeva, P. Impact of interspecific interactions on antimicrobial activity among soil bacteria. Front. Microbiol. 2014, 5, 567. [Google Scholar] [CrossRef]
- Gozzo, F.; Faoro, F. Systemic Acquired Resistance (50 years after discovery): Moving from the lab to the field. J. Agric. Food Chem. 2013, 61, 12473–12491. [Google Scholar] [CrossRef]
- Conrath, U.; Beckers, G.J.; Flors, V.; Garcia-Agustin, P.; Jakab, G.; Mauch, F.; Newman, M.A.; Pieterse, C.M.; Poinssot, B.; Pozo, M.J.; et al. Priming: Getting ready for battle. Mol. Plant Microbe Interact. 2006, 19, 1062–1071. [Google Scholar] [CrossRef]
- Yuan, J.; Li, B.; Zhang, N.; Waseem, R.; Shen, Q.; Huang, Q. Production of bacillomycin- and macrolactin-type antibiotics by Bacillus amyloliquefaciens NJN-6 for suppressing soilborne plant pathogens. J. Agric. Food Chem. 2012, 60, 2976–2981. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, A.; Shihara, H.; Nakajima, H. Isolation of anteiso-C17, iso-C17, iso-C16, and iso-C15 bacillomycin D from Bacillus amyloliquefaciens SD-32 and their antifungal activities against plant pathogens. J. Agric. Food Chem. 2014, 62, 1469–1476. [Google Scholar] [CrossRef] [PubMed]
- Jin, P.; Wang, H.; Liu, W.; Fan, Y.; Miao, W. A new cyclic lipopeptide isolated from Bacillus amyloliquefaciens HAB-2 and safety evaluation. Pest. Biochem. Physiol. 2017, 147, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Peterson, J.; Garges, S.; Giovanni, M.; McInnes, P.; Wang, L.; Schloss, J.A.; Bonazzi, V.; McEwen, J.E.; Wetterstrand, K.A.; Deal, C. The NIH Human Microbiome Project. Genome Res. 2009, 19, 2317–2323. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraiser-Liggett, C.M.; Knight, R.; Gordon, J.I. The human microbiome project: A strategy to understand the microbial components of the human genetic and metabolic landscape and how they contribute to normal physiology and predisposition to disease. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Greenblum, S.; Turnbaugh, P.J.; Borenstein, E. Metagenomic systems biology of the human gut microbiome reveals topological shifts azssociated with obesity and inflammatory bowel disease. Proc. Natl. Acad. Sci. USA 2012, 109, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat. Med. 2013, 19, 576–585. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.A.; Hartmann, M. Networking in the Plant Microbiome. PLoS Biol. 2016, 14, e1002378. [Google Scholar] [CrossRef]
- Berg, G.; Grube, M.; Schloter, M.; Smalla, K. Unraveling the plant microbiome: Looking back and future perspectives. Front. Microbiol. 2014, 5, 148. [Google Scholar] [CrossRef]
- Hallmann, J.; QuadtHallmann, A.; Mahaffee, W.F.; Kloepper, J.W. Bacterial endophytes in agricultural crops. Can. J. Microbiol. 1997, 43, 895–914. [Google Scholar] [CrossRef]
- Eevers, N.; Gielen, M.; Sánchez-López, A.; Jaspers, S.; White, C.; Vangronsveld, J.; Weyens, N. Optimization of isolation and cultivation of bacterial endophytes through addition of plant extract to nutrient media. Microb. Biotechnol. 2015, 8, 707–715. [Google Scholar] [CrossRef]
- Sessitsch, A.; Hardoim, P.; Döring, J.; Weilharter, A.; Krause, A.; Woyke, T.; Mitter, B.; Hauberg-Lotte, L.; Friedrich, F.; Rahalkar, M.; et al. Functional characteristics of an endophyte community colonizing rice roots as revealed by metagenomics analysis. Mol. Plant Microbe Interact. 2012, 25, 28–36. [Google Scholar] [CrossRef]
- Knief, C. Analysis of plant microbe interactions in the era of next generation sequencing technologies. Front. Plant Sci. 2014, 5, 216. [Google Scholar] [CrossRef]
- Yousaf, S.; Bulgari, D.; Bergna, A.; Pancher, M.; Quaglino, F.; Casati, P.; Campisano, A. Pyrosequencing detects human and animal pathogenic taxa in the grapevine endosphere. Front. Microbiol. 2014, 5, 327. [Google Scholar] [CrossRef] [PubMed]
- Jansson, J.K.; Neufeld, J.D.; Moran, M.A.; Gilbert, J.A. Omics for understanding microbial functional dynamics. Environ. Microbiol. 2012, 14, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Zachow, C.; Müller, H.; Philipps, J.; Tilcher, R. Next generation bio-products sowing the seeds of success for sustainable agriculture. Agronomy 2013, 3, 648–656. [Google Scholar] [CrossRef]
- Finkel, O.M.; Delmont, T.O.; Post, A.F.; Belkin, S. Metagenomic signatures of bacterial adaptation to life in the phyllosphere of a salt-secreting desert tree. Appl. Environ. Microbiol. 2016, 82, 2854–2861. [Google Scholar] [CrossRef]
- Proença, D.N.; Francisco, R.; Kublik, S.; Schöler, A.; Vestergaard, G.; Schloter, M.; Morais, P.V. The microbiome of endophytic, wood colonizing bacteria from pine trees as affected by pine wilt disease. Sci. Rep. 2017, 7, 4205. [Google Scholar] [CrossRef]
- Hallmann, J.; Berg, G. Spectrum and population dynamics of bacterial root endophytes. In Microbial Root Endophytes; Schulz, B.J.E., Boyle, C.J.C., Sieber, T.N., Eds.; Springer: Dordercht, The Netherlands, 2006; pp. 15–31. [Google Scholar]
- Van Overbeek, L.S.; van Elsas, J.D. Effects of plant genotype and growth stage on the structure of bacterial communities associated with potato (Solanum tuberosum L.). FEMS Microbiol. Ecol. 2008, 64, 283–296. [Google Scholar] [CrossRef]
- Bulgari, D.; Bozkurt, A.I.; Casati, P.; Caglayan, K.; Quaglino, F.; Bianco, P.A. Endophytic bacterial community living in roots of healthy and ‘Candidatus Phytoplasma mali’-infected apple (Malus domestica, Borkh.) trees. Antonie Van Leeuwenhoek 2012, 102, 677–687. [Google Scholar] [CrossRef]
- Bulgari, D.; Casati, P.; Quaglino, F.; Bianco, P.A. Endophytic bacterial community of grapevine leaves influenced by sampling date and phytoplasma infection process. BMC Microbiol. 2014, 14, 198. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Rott, M.; Schlaeppi, K.; van Themaat, E.V.L.; Ahmadinejad, N.; Assenza, F.; Rauf, P.; Huettel, B.; Reinhardt, R.; Schmelzer, E.; et al. Revealing structure and assembly cues for Arabidopsis root-inhabiting bacterial microbiota. Nature 2012, 488, 91–95. [Google Scholar] [CrossRef]
- Ryan, R.P.; Germaine, K.; Franks, A.; Ryan, D.J.; Dowling, D.N. Bacterial endophytes: Recent developments and applications. FEMS Microbiol. Lett. 2008, 278, 1–9. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; van Themaat, E.V.L.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef]
- Pitzschke, A.; Hirt, H. New insights into an old story: Agrobacterium-induced tumour formation in plants by plant transformation. EMBO J. 2010, 29, 1021–1032. [Google Scholar] [CrossRef]
- Salomon, D.; Kinch, L.N.; Trudgian, D.C.; Guo, X.; Klimko, J.A.; Grishin, N.V.; Mirzaei, H.; Orth, K. Marker for type VI secretion system effectors. Proc. Natl. Acad. Sci. USA 2014, 111, 9271–9276. [Google Scholar] [CrossRef]
- Tyler, H.L.; Triplett, E.W. Plants as a Habitat for beneficial and/or human pathogenic bacteria. Annu. Rev. Phytopathol. 2008, 46, 53–73. [Google Scholar] [CrossRef]
- Holden, N.; Pritchard, L.; Toth, I. Colonization out with the colon: Plants as an alternative environmental reservoir for human pathogenic Enterobacteria. FEMS Microbiol. Rev. 2009, 33, 689–703. [Google Scholar] [CrossRef]
- Compant, S.; Clément, C.; Sessitsch, A. Plant growth-promoting bacteria in the rhizo- and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 2010, 42, 669–678. [Google Scholar] [CrossRef]
- Lugtenberg, B.J.J.; Dekkers, L.C. What makes Pseudomonas bacteria rhizosphere competent? Environ. Microbiol. 1999, 1, 9–13. [Google Scholar] [CrossRef]
- Walker, L.R.; del Moral, R. Primary Succession and Ecosystem Rehabilitation; Cambridge University Press: New York, NY, USA, 2003. [Google Scholar]
- Turner, T.R.; James, E.K.; Poole, P.S. The plant microbiome. Genome Biol. 2013, 14, 209. [Google Scholar] [CrossRef]
- Bulgari, D.; Casati, P.; Faoro, F. Fluorescence in situ hybridization for phytoplasma and endophytic bacteria localization in plant tissues. J. Microbiol. Methods 2011, 87, 220–223. [Google Scholar] [CrossRef]
- Hansen, M.L.; Kregelund, L.; Nybroe, O.; Sørensen, J. Early colonization of barley roots by Pseudomonas fluorescens studied by immunofluorescence technique and confocal laser scanning microscopy. FEMS Microbiol. Ecol. 1997, 23, 353–360. [Google Scholar] [CrossRef]
- Hurek, T.; Reinhold-Hurek, B.; van Montagu, M.; Kellemberg, E. Root colonization and systemic spreeding of Azoarcus sp. Strain BH72 in grasses. J. Bacteriol. 1994, 176, 1913–1923. [Google Scholar] [CrossRef]
- Mahaffee, W.F.; Kloepper, J.W.; van Vurdee, J.W.L.; van der Wolf, J.M.; van den Brink, M. Endophytic colonization of Phaseolus vulgaris by Pseudomonas fluorescent strain 89B-27 and Enterobacter asburiae strain JM22. In Improve Plant Productivity in Rhizosphere Bacteria; Ryder, M.H., Stephens, P.M., Bowen, G.D., Eds.; CSIRO Division of Soils: Melbourne, Australia, 1994; p. 180. [Google Scholar]
- Di Fiore, S.; Del Gallo, M. Endophytic Bacteria: Their Possible Role in the Host Plant. In Azospirillum VI and Related Microorganisms; Fendrik, I., Del Gallo, M., Vanderleyden, J., de Zamaroczy, M., Eds.; NATO ASI Series (Series G: Ecological Sciences); Springer: Berlin/Heidelberg, Germany, 1995; Volume 37. [Google Scholar]
- Mitter, B.; Petric, A.; Shin, M.W.; Chain, P.S.G.; Hauberg-Lotte, L.; Reinhold-Hurek, B.; Nowak, J.; Sessitsch, A. Comparative genome analysis of Burkholderia phytofirmans PsJN reveals a wide spectrum of endophytic lifestyles based on interaction strategies with host plants. Front. Plant Sci. 2013, 4, 120. [Google Scholar] [CrossRef]
- Hardoim, P.R.; van Overbeek, L.S.; van Elsas, J.D. Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 2008, 16, 463–471. [Google Scholar] [CrossRef]
- Millet, Y.A.; Danna, C.H.; Clay, N.K.; Songnuan, W.; Simon, M.D.; Werck-Reichhart, D.; Ausubela, F.M. Innate immune responses activated in Arabidopsis roots by Microbe-Associated Molecular Patterns. Plant Cell 2010, 22, 973–990. [Google Scholar] [CrossRef]
- Zamioudis, C.; Pieterse, C.M.J. Modulation of host immunity by beneficial microbes. Mol. Plant Microbe Interact. 2012, 25, 139–150. [Google Scholar] [CrossRef]
- Bulgari, D.; Casati, P.; Brusetti, L.; Quaglino, F.; Brasca, M.; Daffonchio, D.; Bianco, P.A. Endophytic bacterial diversit y in grapevine (Vitis vinifera L.) leaves described by 16S rRNA gene sequence analysis and Length Heterogeneity-PCR. J. Microbiol. 2009, 47, 393–401. [Google Scholar] [CrossRef]
- Tondello, A. Endophyte Watching: Combining Molecular and Microscopy Approaches to Isolate, Identify, Tag, and Monitor Fungi and Bacteria Inside Plants. Ph.D. Thesis, Università di Padova, Padua, Italy, 2009. [Google Scholar]
- Ryan, E.T. The intestinal pathobiome: Its reality and consequences among infants and young children in resource-limited settings. J. Infect. Dis. 2013, 208, 1732–1733. [Google Scholar] [CrossRef]
- Kirzinger, M.W.B.; Nadarasah, G.; Stavrinides, J. Insights into Cross-Kingdom Plant Pathogenic Bacteria. Genes 2011, 2, 980–997. [Google Scholar] [CrossRef]
- Campisano, A.; Ometto, L.; Compant, S.; Pancher, M.; Antonielli, L.; Yousaf, S.; Varotto, C.; Anfora, G.; Pertot, I.; Sessitsch, A.; et al. Interkingdom transfer of the acne-causing agent, Propionibacterium acnes, from human to grapevine. Mol. Biol. Evol. 2014, 31, 1059–1065. [Google Scholar] [CrossRef]
- Nithya, A.; Babu, S. Prevalence of plant beneficial and human pathogenic bacteria isolated from salad vegetables in India. BMC Microbiol. 2017, 17, 64. [Google Scholar] [CrossRef]
- Sharma, K.; Goss, E.M.; Dickstein, E.R.; Smith, M.; Southwick, F.; van Bruggen, A.H.C. Exserohilum rostratum: Characterization of a cross-kingdom pathogen. PLoS ONE 2014, 9, e108691. [Google Scholar] [CrossRef]
- Van Diepeningen, A.D.; de Hoog, G.S. Challenges in Fusarium, a trans-kingdom pathogen. Mycopathologia 2016, 181, 161–163. [Google Scholar] [CrossRef]
- Hayward, A.C.; Waterston, J.M. Agrobacterium Tumefaciens. Descriptions of Pathogenic Fungi and Bacteria; No. 42; Commonwealth Mycological Institute, Kew: Surrey, UK, 1965. [Google Scholar]
- Paphitou, N.I.; Rolston, K.V. Catheter-related bacteremia caused by Agrobacterium radiobacter in a cancer patient: Case report and literature review. Infection 2003, 31, 421–424. [Google Scholar]
- Lacroix, B.; Tzfira, T.; Vainstein, A.; Citovsky, V. A case of promiscuity: Agrobacterium’s endless hunt for new partners. Trends Genet. 2006, 22, 29–37. [Google Scholar] [CrossRef]
- Van Baarlen, P.; van Belkum, A.; Summerbell, R.C.; Crous, P.W.; Thomma, B.P.H.J. Molecular mechanisms of pathogenicity: How do pathogenic microorganisms develop cross-kingdom host jumps? FEMS Microbiol. Rev. 2007, 31, 239–277. [Google Scholar] [CrossRef]
- Suárez-Moreno, Z.R.; Caballero-Mellado, J.; Coutinho, B.C.; Mendonça-Previato, L.; James, E.K.; Venturi, V. Common features of environmental and potentially beneficial plant-associated Burkholderia. Microb. Ecol. 2012, 63, 249–266. [Google Scholar] [CrossRef]
- Weisskopf, L.; Heller, S.; Eberl, L. Burkholderia species are major inhabitants of white lupin cluster roots. Appl. Environ. Microbiol. 2011, 77, 7715–7720. [Google Scholar] [CrossRef]
- Estrada-de los Santos, P.; Vinuesa, P.; Martínez-Aguilar, L.; Hirsch, A.M.; Caballero-Mellado, J. Phylogenetic analysis of Burkholderia species by multilocus sequence analysis. Curr. Microbiol. 2013, 67, 51–60. [Google Scholar] [CrossRef]
- Eberl, L.; Vandamme, P. Members of the genus Burkholderia: Good and bad guys [version 1; referees: 3 approved]. F1000 Res. 2016, 5. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Lee, H.H.; Park, J.; Park, I.; Seo, Y.S. Computational identification and comparative analysis of secreted and transmembrane proteins in six Burkholderia species. Plant Pathol. J. 2017, 33, 148–162. [Google Scholar] [CrossRef]
- Estrada-de los Santos, P.; Rojas-Rojas, F.U.; Tapia-García, E.Y.; Vásquez-Murrieta, M.S.; Hirsch, A.M. To split or not to split: An opinion on dividing the genus Burkholderia. Ann. Microbiol. 2016, 66, 1303–1314. [Google Scholar] [CrossRef]
- Lòpez-Fernàndez, S.; Sonego, P.; Moretto, M.; Pancher, M.; Engelen, K.; Pertot, I.; Campisano, A. A Whole-genome comparative analysis of virulence genes unveils similarities and differences between endophytes and other symbiotic bacteria. Front. Microbiol. 2015, 6, 419. [Google Scholar] [CrossRef]
- Sheibani-Tezerji, R.; Rattei, T.; Sessitsch, A.; Trognitz, F.; Mitter, B. Transcriptome Profiling of the Endophyte Burkholderia phytofirmans PsJN Indicates Sensing of the Plant Environment and Drought Stress. MBio 2015, 6, e00621-15. [Google Scholar] [CrossRef]
- Berg, G.; Eberl, L.; Hartmann, A. The rhizosphere as a reservoir for opportunistic human pathogenic bacteria. Environ. Microbiol. 2005, 7, 1673–1685. [Google Scholar] [CrossRef]
- Teplitski, M.; Warriner, K.; Bartz, J.; Schneider, K.R. Untangling metabolic and communication networks: Interactions of enterics with phytobacteria and their implications in produce safety. Trends Microbiol. 2011, 19, 121–127. [Google Scholar] [CrossRef]
- Gu, G.; Hu, J.; Cevallos-Cevallos, J.M.; Richardson, S.M.; Bartz, J.A.; van Bruggen, A.H.C. Internal colonization of Salmonella enterica serovar Typhimurium in tomato plants. PLoS ONE 2011, 6, e27340. [Google Scholar] [CrossRef]
- Cevallos-Cevallos, J.M.; Gu, G.; Danyluk, M.D.; van Bruggen, A.H.C. Adhesion and splash dispersal of Salmonella enterica Typhimurium on tomato leaflets: Effects of rdar morphotype and trichome density. Int. J. Food Microbiol. 2012, 160, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Gu, G.; Cevallos-Cevallos, J.M.; van Bruggen, A.H.C. Ingress of Salmonella enteric Typhimurium into tomato leaves through hydathodes. PLoS ONE 2013, 8, e53470. [Google Scholar] [CrossRef]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, M. The role of root exudates in rhizosphere interactions with plant and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [PubMed]
- Franz, E.; van Bruggen, A.H.C. Ecology of E. coli O157:H7 and Salmonella enterica in the primary vegetable production chain. Crit. Rev. Microbiol. 2008, 34, 143–161. [Google Scholar] [CrossRef] [PubMed]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef]
- Lynch, M.F.; Tauxe, R.V.; Hedberg, C.W. The growing burden of food-borne outbreaks due to contaminated fresh produce: Risks and opportunities. Epidemiol. Infect. 2009, 137, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Barak, J.D.; Schroeder, B.K. Interrelationships of food safety and plant pathology: The life cycle of human pathogens on plants. Annu. Rev. Phytopathol. 2012, 50, 241–266. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Jiang, X. Microbiological safety of chicken litter or chicken litter-based organic fertilizers: A review. Agriculture 2014, 4, 1–29. [Google Scholar] [CrossRef]
- Seo, K.H.; Frank, J.F. Attachment of Escherichia coli O157:H7 to lettuce leaf surface and bacterial viability in response to chlorine treatment as demonstrated by using confocal scanning laser microscopy. J. Food Prot. 1999, 62, 3–9. [Google Scholar] [CrossRef]
- Saldaña, Z.; Sánchez, E.; Xicohtencatl-Cortes, J.; Puente, J.L.; Girón, J.A. Surface structures involved in plant stomata and leaf colonization by shiga-toxigenic Escherichia coli O157:H7. Front. Microbiol. 2011, 2, 119. [Google Scholar] [CrossRef]
- Hofmann, A.; Fischer, D.; Hartmann, A.; Schmid, M. Colonization of plants by human pathogenic bacteria in the course of organic vegetable production. Front. Microbiol. 2014, 5, 191. [Google Scholar] [CrossRef] [PubMed]
- Melotto, M.; Panchal, S.; Roy, D. Plant innate immunity against human bacterial pathogens. Front. Microbiol. 2014, 5, 411. [Google Scholar] [CrossRef] [PubMed]
- Gomes, C.; Da Silva, P.; Moreira, R.G.; Castell-Perez, E.; Ellis, E.A.; Pendleton, M. Understanding E. coli internalization in lettuce leaves for optimization of irradiation treatment. Int. J. Food Microbiol. 2009, 135, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Bilek, S.E.; Turantas, F. Decontamination efficiency of high power ultrasound in the fruit and vegetable industry, a review. Int. J. Food Microbiol. 2013, 166, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Ziuzin, D.; Han, L.; Cullen, P.J.; Bourke, P. Cold plasma inactivation of internalized bacteria and biofilms for Salmonella enterica serovar Typhimurium, Listeria monocytogenes and Escherichia coli. Int. J. Food Microbiol. 2015, 210, 53–61. [Google Scholar] [CrossRef]
- Cooley, M.B.; Miller, W.G.; Mandrell, R.E. Colonization of Arabidopsis thaliana with Salmonella enterica and enterohemorrhagic Escherichia coli O157:H7 and competition by Enterobacter asburiae. Appl. Environ. Microbiol. 2003, 69, 4915–4926. [Google Scholar] [CrossRef]
- Wright, K.M.; Crozier, L.; Marshall, J.; Merget, B.; Holmes, A.; Holden, N.J. Differences in internalization and growth of Escherichia coli O157:H7 within the apoplast of edible plants, spinach and lettuce, compared with the model species Nicotiana benthamiana. Microbial Biotechnol. 2017, 10, 555–569. [Google Scholar] [CrossRef]
- Girardin, S.E.; Sansonetti, P.J.; Philpott, D.J. Intracellular vs extracellular recognition of pathogens—Common concepts in mammals and flies. Trends Microbiol. 2002, 10, 193–199. [Google Scholar] [CrossRef]
- Green, E.; Mecsas, J. Bacterial Secretion Systems: An Overview. In Virulence Mechanisms of Bacterial Pathogens, 5th ed.; Kudva, I., Cornick, N., Plummer, P., Zhang, Q., Nicholson, T., Bannantine, J., Bellaire, B.H., Eds.; ASM Press: Washington, DC, USA, 2016; pp. 215–239. [Google Scholar]
- Ronald, P.C.; Beutler, B. Plant and animal sensors of conserved microbial signatures. Science 2010, 330, 1061–1064. [Google Scholar] [CrossRef]
- Jacob, F.; Vernaldi, S.; Maekawa, T. Evolution and conservation of plant NLR functions. Front. Immunol. 2013, 4, 297. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Elinav, T.; Strowig, J.; Henao-Mejia, J.; Flavell, R.A. Regulation of the antimicrobial response by NLR proteins. Immunity 2011, 34, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.V.; Charriera, A.; Schikorab, A.; Bigearda, J.; Pateyronc, S.; de Tauzia-Moreaua, M.L.; Evrard, A.; Mithöfer, A.; Martin-Magniette, M.L.; Virlogeux-Payant, I.; et al. Salmonella enterica Flagellin Is Recognized via FLS2 and Activates PAMP-Triggered Immunity in Arabidopsis thaliana. Mol. Plant 2014, 7, 657–674. [Google Scholar] [CrossRef] [PubMed]
- Iriti, M.; Franco, F. Review of innate and specific immunity in plants and plant. Mycopathologia 2007, 164, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Thomma, B.P.H.J.; Cammue, B.P.A.; Thevissen, K. Plant defensins. Planta 2002, 216, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Kroupitski, Y.; Golberg, D.; Belausov, E.; Pinto, R.; Swartzberg, D.; Granot, D.; Sela, S. Internalization of Salmonella enterica in leaves is induced by light and involves chemotaxis and penetration through open stomata. Appl. Environ. Microbiol. 2009, 75, 6076–6086. [Google Scholar] [CrossRef] [PubMed]
- Berger, C.N.; Shaw, R.K.; Brown, D.J.; Mather, H.; Clare, S.; Dougan, G.; Pallen, M.J.; Frankel, G. Interaction of Salmonella enterica with basil and other salad leaves. ISME J. 2009, 3, 261–265. [Google Scholar] [CrossRef]
- López-Gálvez, F.; Gil, M.I.; Allende, A. Impact of relative humidity, inoculum carrier and size, and native microbiota on Salmonella ser. Typhimurium survival in baby lettuce. Food Microbiol. 2018, 7, 155–161. [Google Scholar] [CrossRef]
- Brunner, F.; Fraiture, M. Killing two birds with one stone: Trans-kingdom suppression of PAMP/MAMP-induced immunity by T3E from enteropathogenic bacteria. Front. Microbiol. 2014, 5, 320. [Google Scholar] [CrossRef]
- Neumann, C.; Fraiture, M.; Hernàndez-Reyes, C.; Akum, F.N.; Virlogeux-Payant, I.; Chen, Y.; Pateyron, S.; Colcombet, J.; Kogel, K.H.; Hirt, H.; et al. The Salmonella effector protein SpvC, a phosphothreonine lyase is functional in plant cells. Front. Microbiol. 2014, 5, 548. [Google Scholar] [CrossRef]
- Brown, N.A.; Urban, M.; Hammond-Kosack, K.E. The trans-kingdom identification of negative regulators of pathogen hypervirulence. FEMS Microbiol. Rev. 2016, 40, 19–40. [Google Scholar] [CrossRef]
- Heffron, F.; Niemann, G.; Yoon, H.; Kidwai, A.; Brown, R.; McDemrott, J.; Smith, R.D.; Adkins, J.N. Salmonella-secreted virulence factors. In Salmonella from Genome to Function; Porwollik, S., Ed.; Caister Academic Press: San Diego, CA, USA, 2011; pp. 187–223. [Google Scholar]
- Schikora, A.; Virlogeux-Payant, I.; Bueso, E.; Garcia, A.V.; Nilau, T.; Charrier, A.; Pelletier, S.; Menanteau, P.; Baccarini, M.; Velge, P.; et al. Conservation of Salmonella infection mechanisms in plants and animals. PLoS ONE 2011, 6, e24112. [Google Scholar] [CrossRef] [PubMed]
- Shirron, N.; Yaron, S. Active suppression of early immune response in tobacco by the human pathogen Salmonella Typhimurium. PLoS ONE 2011, 6, e18855. [Google Scholar] [CrossRef] [PubMed]
- Galan, J.E. Common themes in the design and function of bacterial effectors. Cell Host Microbe 2009, 5, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Potnis, N.; Soto-Arias, J.P.; Cowles, K.; van Bruggen, A.H.C.; Jones, J.B.; Barak, J.D. Stealth colonization of tomato leaf by Xanthomonas perforans enhances persistence of Salmonella enterica in the phyllosphere. Appl. Environ. Microbiol. 2014, 80, 3173–3180. [Google Scholar] [CrossRef]
- Monack, D.M.; Mueller, A.; Falkow, S. Persistent bacterial infections: The interface of the pathogen and the host immune system. Nat. Rev. Microbiol. 2004, 2, 747–765. [Google Scholar] [CrossRef] [PubMed]
- Parker, C.T.; Sperandio, V. Cell-to-cell signalling during pathogenesis. Cell Microbiol. 2009, 11, 363–369. [Google Scholar] [CrossRef]
- Bulgari, D.; Casati, P.; Crepaldi, P.; Daffonchio, D.; Quaglino, F.; Brusetti, L.; Bianco, P.A. Restructuring of endophytic bacterial communities in grapevine yellows-diseased and recovered Vitis vinifera L. plants. Appl. Environ. Microbiol. 2011, 77, 5018–5022. [Google Scholar] [CrossRef]
- Cardenas, A.; Rodriguez, R.L.; Pizarro, V.; Cadavid, L.F.; Arevalo-Ferro, C. Shifts in bacterial communities of two Caribbean reef-building coral species affected by white plague disease. ISME J. 2012, 6, 502–512. [Google Scholar] [CrossRef]
- Casadevall, A.; Pirofski, L.A. Host-Pathogen interactions: Basic concepts of microbial commensalism, colonization, infection, and disease. Infect. Immun. 2000, 68, 6511–6518. [Google Scholar] [CrossRef]
- Rosas-Magallanes, V.; Deschavanne, P.; Quintana-Murci, L.; Brosch, R.; Gicquel, B.; Neyrolles, O. Horizontal Transfer of a Virulence Operon to the Ancestor of Mycobacterium tuberculosis. Mol. Biol. Evol. 2006, 23, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Kelly, B.G.; Vespermann, A.; Bolton, D.J. Horizontal gene transfer of virulence determinants in selected bacterial food borne pathogens. Food Chem. Toxicol. 2009, 47, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Qin, S.; Lan, Y.; Liu, M.; Cao, X.; Qiao, D.; Cao, Y.; Cao, Y. Comparative genomic analysis of Paenibacillus sp. SSG-1 and its closely related strains reveals the effect of glycometabolism on environmental adaptation. Sci. Rep. 2017, 7, 5720. [Google Scholar] [CrossRef] [PubMed]
- Padda, K.P.; Puri, A.; Chanway, C.P. Paenibacillus polymyxa: A prominent biofertilizer and biocontrol agent for sustainable agriculture. In Agriculturally Important Microbes for Sustainable Agriculture; Meena, V., Mishra, P., Bisht, J., Pattanayak, A., Eds.; Springer: Singapore, 2017; Volume 2, pp. 165–191. [Google Scholar]
- Mikkola, R.; Andersson, M.A.; Grigoriev, P.; Heinonen, M.; Salkinoja-Salonen, M.S. The toxic mode of action of cyclic lipodepsipeptide fusaricidins, produced by Paenibacillus polymyxa, toward mammalian cells. J. Appl. Microbiol. 2017, 123, 436–449. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Term | Definition | Reference |
|---|---|---|
| Biocontrol | The use of living organisms to control plant pathogens and pests resulting in plant growth promotion | [2] |
| Biostimulants | Formulated products with novel, or emergent properties due the complex of constituents, that improve plant productivity not as a sole consequence of the presence of known essential plant nutrients, plant growth regulators, or plant protective compounds | [3] |
| Biopesticide | Biological pesticides are derived from natural materials including plants, animals and microbe, and some minerals | [4] |
| Biofertilizer | A biofertilizer is any bacterial or fungal inoculant applied to plants with the aim of increasing the availability of nutrients and their utilization by plants, regardless of the nutrient content of the inoculant itself. Biofertilizers may also be defined as microbial biostimulants improving plant nutrition efficiency. | [5] |
| Phytostimulator | Microorganism with the ability to produce or change the concentration of growth regulators, such as indole acetic acid, gibberellic acid, cytokinins, and ethylene | [6] |
| Pathogen | A microbe that can cause physiological and structural damages in a host | [7] |
| Biocontrol agents | Living organisms that show the ability to directly or indirectly antagonize plant pathogens and pests | here defined |
| Endophytic bacteria | Bacteria that habit for all or part of their lifetime, in the internal part of a plant | [8] |
| Plant microbiome | A community of microbes associated with a plant and their crosstalk with the plant genome and proteome | [9] |
| Pathobiome | The complex interactions of pathogenic microbes which may influence or drive disease processes and their relationship to the ‘normal’ microbiome of the organism in question | [10,11] |
| Phytonoses | New group of diseases caused by human pathogens that are transmitted via consumption of fresh produce | [12,13] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bulgari, D.; Montagna, M.; Gobbi, E.; Faoro, F. Green Technology: Bacteria-Based Approach Could Lead to Unsuspected Microbe–Plant–Animal Interactions. Microorganisms 2019, 7, 44. https://doi.org/10.3390/microorganisms7020044
Bulgari D, Montagna M, Gobbi E, Faoro F. Green Technology: Bacteria-Based Approach Could Lead to Unsuspected Microbe–Plant–Animal Interactions. Microorganisms. 2019; 7(2):44. https://doi.org/10.3390/microorganisms7020044
Chicago/Turabian StyleBulgari, Daniela, Matteo Montagna, Emanuela Gobbi, and Franco Faoro. 2019. "Green Technology: Bacteria-Based Approach Could Lead to Unsuspected Microbe–Plant–Animal Interactions" Microorganisms 7, no. 2: 44. https://doi.org/10.3390/microorganisms7020044
APA StyleBulgari, D., Montagna, M., Gobbi, E., & Faoro, F. (2019). Green Technology: Bacteria-Based Approach Could Lead to Unsuspected Microbe–Plant–Animal Interactions. Microorganisms, 7(2), 44. https://doi.org/10.3390/microorganisms7020044