1. Introduction
The timid nature of Pekin ducks makes them easily stressed. Intensively reared ducks are constantly exposed to various adverse influences, such as noise, crowding, handling, transport, high temperature, and immune challenges, which leads to chronic stress [
1,
2]. Stress in poultry can activate the hypothalamic pituitary adrenal axis, resulting in increased corticosterone (CORT) secretion [
3]. Therefore, the serum CORT level is considered a reliable stress indicator in poultry [
4]. Recently, numerous publications have demonstrated that a CORT challenge can reliably induce physiological stress in rodents and birds [
5,
6]. Additionally, a CORT challenge can also lead to decreased body weight, and can influence lipid metabolism and immune system responses [
7,
8,
9].
The liver plays an important role in maintaining metabolic homeostasis [
10]. A fatty liver, oxidative stress, and inflammation are the major mechanisms underlying the pathogenesis of liver injury induced by CORT [
8,
11,
12]. Hepatic inflammation and lipogenesis can be affected by a number of hormones, including glucocorticoids, which modulate metabolic homeostasis and stress responses [
13,
14]. Glucocorticoids are important regulators of lipid metabolism, and chronic low level exposure to glucocorticoids promotes lipogenesis to produce fatty liver [
15,
16]. CORT is the primary active form of glucocorticoids and is considered as an induced indicator of fatty liver and stress [
17]. Chronic high levels of CORT are closely related to fatty liver progression and inflammatory responses [
18]. Exogenous CORT administration to animals can induce abnormal lipid accumulation and an inflammatory response in the liver that can progress to fatty liver diseases [
19,
20]. However, no relevant studies in Pekin ducks have been reported in the literature.
Clostridium butyricum (
C. butyricum), regarded as a beneficial probiotic, exists in the intestinal tract and acts to protect against pathogenic bacteria and intestinal injury [
21]. Direct feeding of
C. butyricum to different animals (such as broilers, Pekin ducks and shrimps) can enhance growth performance, antioxidant properties, as well as meat quality, and modulate intestinal microflora, balance intestinal health, and regulate the immune system by activating related genes and signaling pathways [
22,
23,
24]. Beyond its beneficial effects in the digestive tract,
C. butyricum can protect mice against non-alcoholic fatty liver disease (NAFLD) induced by a high fat diet [
25] and the anti-oxidation and anti-inflammation protective effects of
C. butyricum have also been demonstrated in previous studies for other diseases [
26,
27]. Although the probiotic
C. butyricum can improve the intestinal health and immune status of weaned piglets challenged with lipopolysaccharide (LPS) [
28] and suppress hepatic damage degree in mice challenged with CORT [
25], it is largely unknown whether probiotic
C. butyricum intervention can attenuate liver injury in Pekin ducks under CORT challenge. Transcriptome analysis is an effective technique to identify the genes that are differentially expressed and signaling pathways that are affected by environmental endocrine disruptors and we have already used transcriptome analysis to demonstrate the effects of dietary
C. butyricum on the breast muscle of Pekin ducks [
29]. Therefore, it is interesting to use the RNA-sequencing (RNA-seq) method in order to determine the underlying hepatoprotective mechanisms active in Pekin ducks supplemented with dietary
C. butyricum under CORT challenge through a combined analysis of lipid metabolism and inflammation response.
In this study, we aimed to investigate whether dietary C. butyricum intervention can alleviate CORT-induced oxidative stress, inflammation, and lipid accumulation in the liver of Pekin ducks, and to explore the underlying mechanisms of these effects using the RNA-sequencing technique. We hope our study can help to solve the stress problems of Pekin duck in the breeding process for the future, and eventually enhance the associated economic income.
2. Materials and Methods
2.1. Ethics Statement
This study was carried out in accordance with the recommendations of Animal Care and Use Ethics Committee of China Agricultural University under protocol code CAU20180428-2. The protocol was approved by the Animal Care and Use Ethics Committee of China Agricultural University (approval date: 28 April 2018).
2.2. Animals and Experimental Design
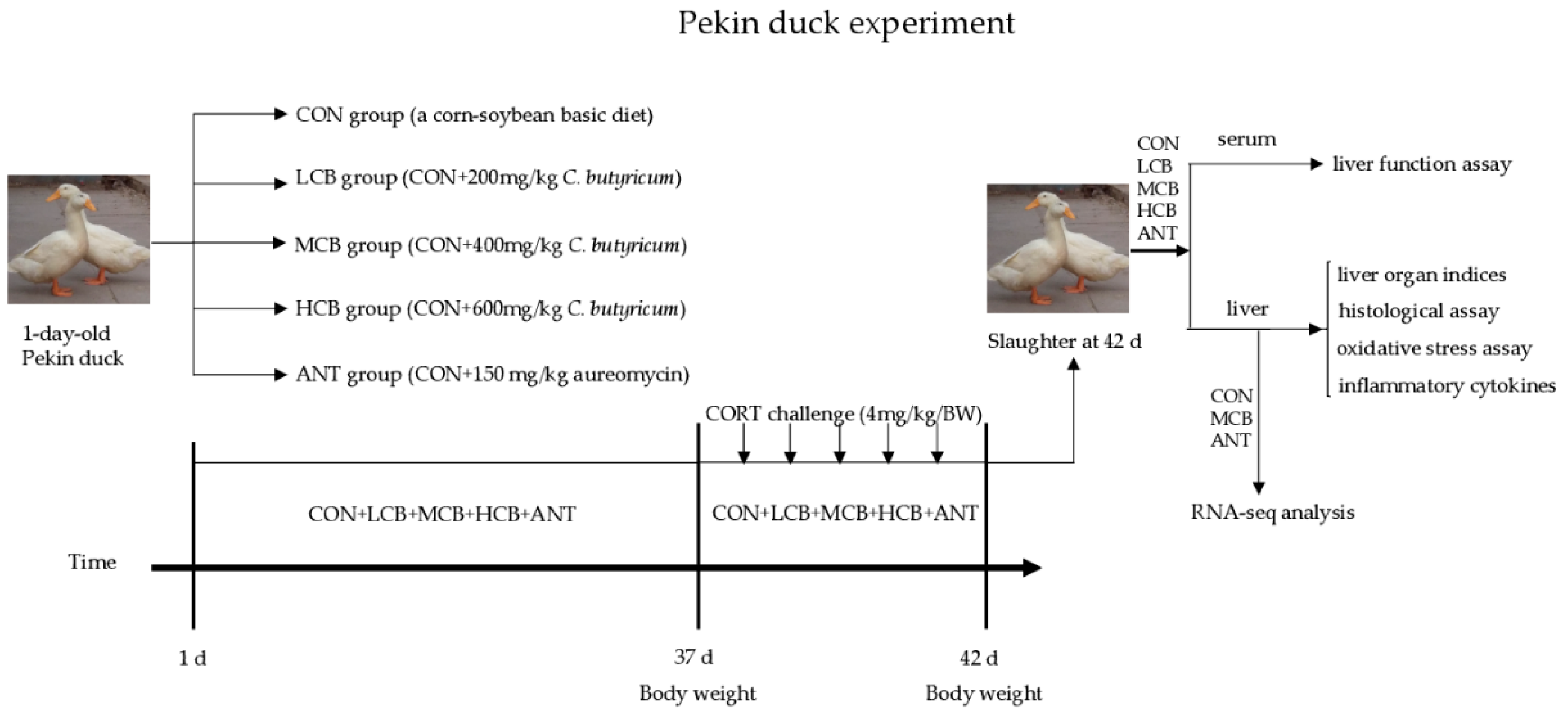
A total of 500 male Pekin ducks (1-day-old) were selected from a local commercial hatchery (Beijing Golden Star Duck Co., Ltd.) and the experiment was performed at the Experimental Center of China Agricultural University. The birds were randomly assigned into five groups (five replicates with twenty birds each) and were raised in a temperature-controlled room. The temperature was controlled at 32 to 34 °C during the first 5 days, and then subsequently declined (about 2–3 °C/week) to a final temperature of 22 °C. All ducks had free access to feed and drinking water with a 23L: 1D lighting system during the experiment. The control group (CON) was fed with a corn-soybean basic diet for 42 d. The other four groups were fed with the basic diet supplemented with 200 mg/kg (LCB), 400 mg/kg (MCB), or 600 mg/kg (HCB)
C. butyricum (2.0 × 10
9 CFU/g) or 150 mg/kg aureomycin (ANT), respectively (
Figure 1). The nutrient requirements of the diets were calculated to meet or exceed the National Research Council (NRC, 1994) standards (
Table S1). The
C. butyricum is available as a freeze-dried powder obtained from Beijing Shine Biology Technology Co., Ltd., China (Batch No. 20170325003).
At 37-days-old, twelve ducks were randomly selected from each group and challenged with CORT (4 mg/kg, Sigma-Aldrich, Shanghai, China) for 5 days. CORT was premixed in corn oil (1:1) and injected into the abdominal cavity. The dose of 4 mg/kg was demonstrated previously to enhance serum CORT content. Injections were done once daily between 07:00 and 08:00.
2.3. Sample Collection
The body weight of the selected 12 ducks from each group was recorded after feed fasting for 6 h at 37- and 42-days old. On day 42, blood samples (5 mL) were taken from the wing vein for biochemical analysis. Subsequently, all 12 ducks were euthanized by giving sodium pentobarbitone solution (30 mg/kg/BW) intravenously through the leg anesthetic to reduce discomfort. After dissection, the entire liver was weighed to calculate the liver organ indices. All liver samples were frozen in liquid nitrogen immediately and stored at –80 °C until further RNA-seq analysis.
2.4. Histological Examination of Hepatic Tissue
Liver samples were freshly dissected and, after appropriate fixation (10% formaldehyde neutral buffer solution) and paraffin embedding, samples were sectioned at a thickness of 4 μm and stained with hematoxylin and eosin (H&E). Frozen liver sections were made at −18 °C using a Leica kryostat (Leica CM3050S, Leica instrument GmbH, Germany), then stained with Oil Red O (O0625; Sigma-Aldrich Co., St. Louis, MO, USA) for 15 min and counterstained with hematoxylin. The slides were observed using an optical microscope (Olympus BX50 microscope; Olympus Corporation, Tokyo, Japan).
2.5. Measurement of Liver Function Indices
The levels of aspartate aminotransferase (AST), alanine aminotransferase (ALT), and gamma-glutamyltranspeptidase (γ-GGT) in serum were measured using the assay kits (Nanjing Jiancheng Inc., Nanjing, China) according to the manufacturer’s instructions. The total protein (TP), albumin (ALB), and globulin (GLB) were detected using a Roche Cobas702 automatic biochemical analyzer (Roche, Basel Switzerland).
2.6. Detection of Oxidative Stress and Cytokines
Liver tissues (200 mg) were homogenized in 0.9% physiological saline to obtain a 10% homogenate. The total antioxidant capacity (T-AOC), total superoxide dismutase (T-SOD), and glutathione peroxidase (GSH-PX) activities as well as the reactive oxygen species (ROS) and malondialdehyde (MDA) contents of the 10% homogenate were measured using commercial kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). The total hepatic protein was detected by the method of Coomassie brilliant blue using the commercial kit (Nanjing Jiancheng Bioengineering Institute, China). The concentrations of interleukin-1β (IL-1β), tumor necrosis factor-α (TNF-α), IL-6, IL-4, IL-10 and transforming growth factor-β (TGF-β) in the liver homogenate were detected using commercial ELISA kits (Nanjing Jiancheng Bioengineering Institute, China). All procedures were performed according to the manufacturer’s instructions. The levels of CORT in serum were determined by the Beijing Huaying Biotechnology Company (Beijing, China) using a radioimmunoassay method.
2.7. RNA Isolation, Library Preparation, and Sequencing
The liver samples of nine randomly selected ducks (three from each of the CON, MCB, and ANT groups) were separately ground for 30 s using the easy grind method in liquid nitrogen. Total RNA was then extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) following the manufacturer’s protocol. RNA quality and purity were determined using a Nano-Drop 2000 spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA) at 260 and 280 nm, and RNA integrity was checked on an Agilent 2100 Bionalyzer (Agilent Technologies, Les Ullis, France) and by 1.2% agarose gel electrophoresis. Only RNA samples with OD260/OD280 ratios between 1.8 and 2.2 and an RNA integrity number (RIN) ≥ 8.0 were selected for cDNA library generation. Next, RNA samples from the nine ducks were then packed in dry ice and sent to Shanghai Majorbio Bio-pharm Technology Co., Ltd. (Shanghai, China) for further library preparation and sequencing. The cDNA library was sequenced using an Illumina HiSeq 4000 (Illumina, San Diego, CA, USA) using paired-end technology.
The quality of raw reads was evaluated using the FastQC program (
http://www.bioinformatics.babraham.ac.uk/projects/fastqc/). Raw reads with a threshold quality < 20, reads shorter than 50 nt, as well as reads containing adapter sequences, poly-N, or the sequencing primer were removed using SeqPrep software (
https://github.com/jstjohn/SeqPrep) and Sickle software (
https://github.com/najoshi/sickle) to obtain clean reads. Additionally, the quality of the clean data was evaluated according to Error%, Q30, GC content%, average read length and sequence duplication level. The high-quality clean reads were used for subsequent analyses and functional annotation. The high quality clean reads of the nine samples were separately mapped to the reference
Anas platyrhynchos genome (
http://asia.ensembl.org/Anas_platyrhynchos/Info/Index?db=core) using TopHat2 (version: 2.0.9) [
30]. The normalization of gene expression levels was obtained by converting the RNA-seq transcript abundance into fragments per kilobase per million mapped reads (FPKM) [
31]. The RNA-seq datasets used and/or analyzed during the study are available in the NCBI Sequence Read Archive (SRA) under the accession number SRP134223.
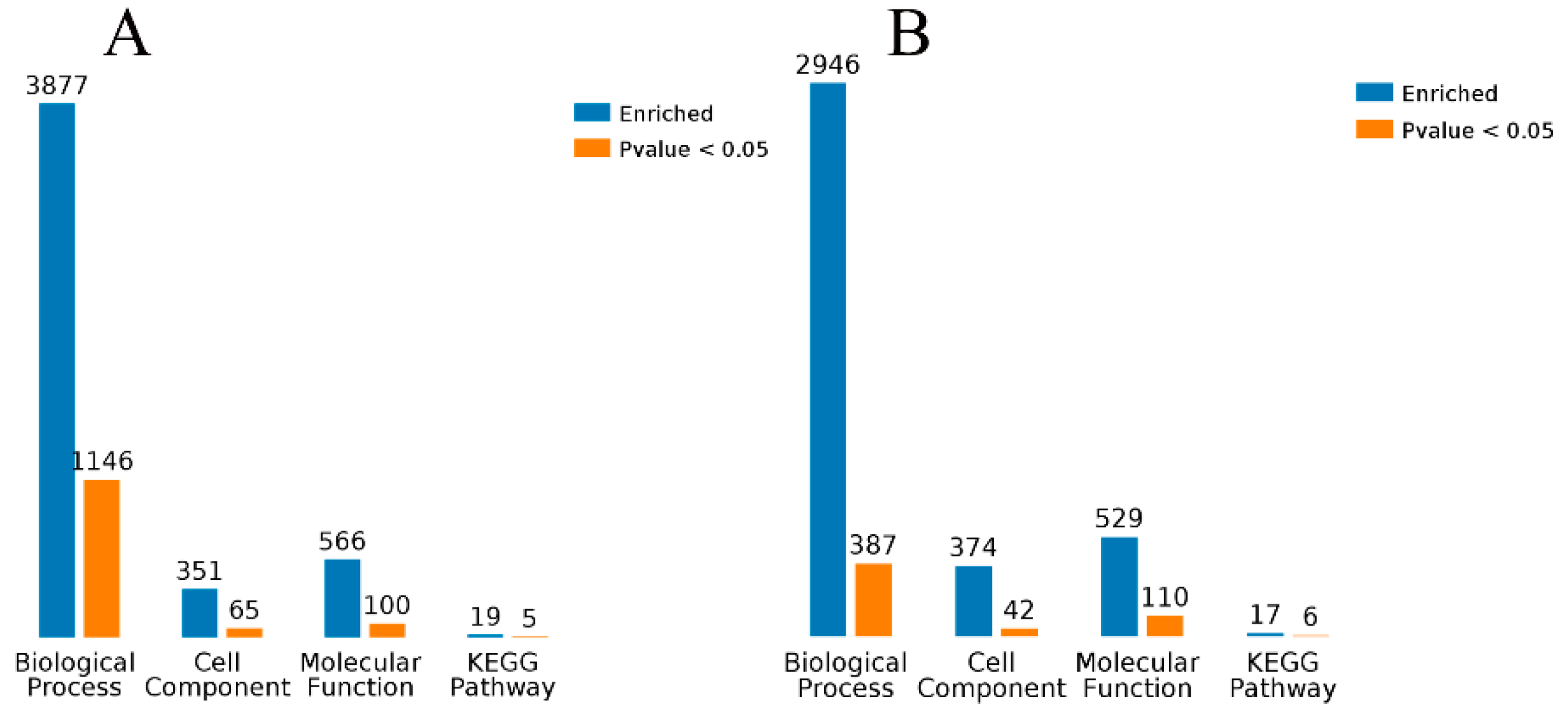
2.8. Enrichment Analysis of GO Terms and KEGG Pathways for DEGs
The edgeR software (
http://www.bioconductor.org/packages/2.12/bioc/html/edgeR.html) was used to identify differentially expressed genes (DEGs) among the three groups and fold change was calculated based on FPKM values. The
p values were adjusted by a false discovery rate (FDR) correction method. Genes with a corrected
p-value < 0.05 and fold-change ≥ 2 or ≤ 0.5 were considered to be significantly differentially expressed.
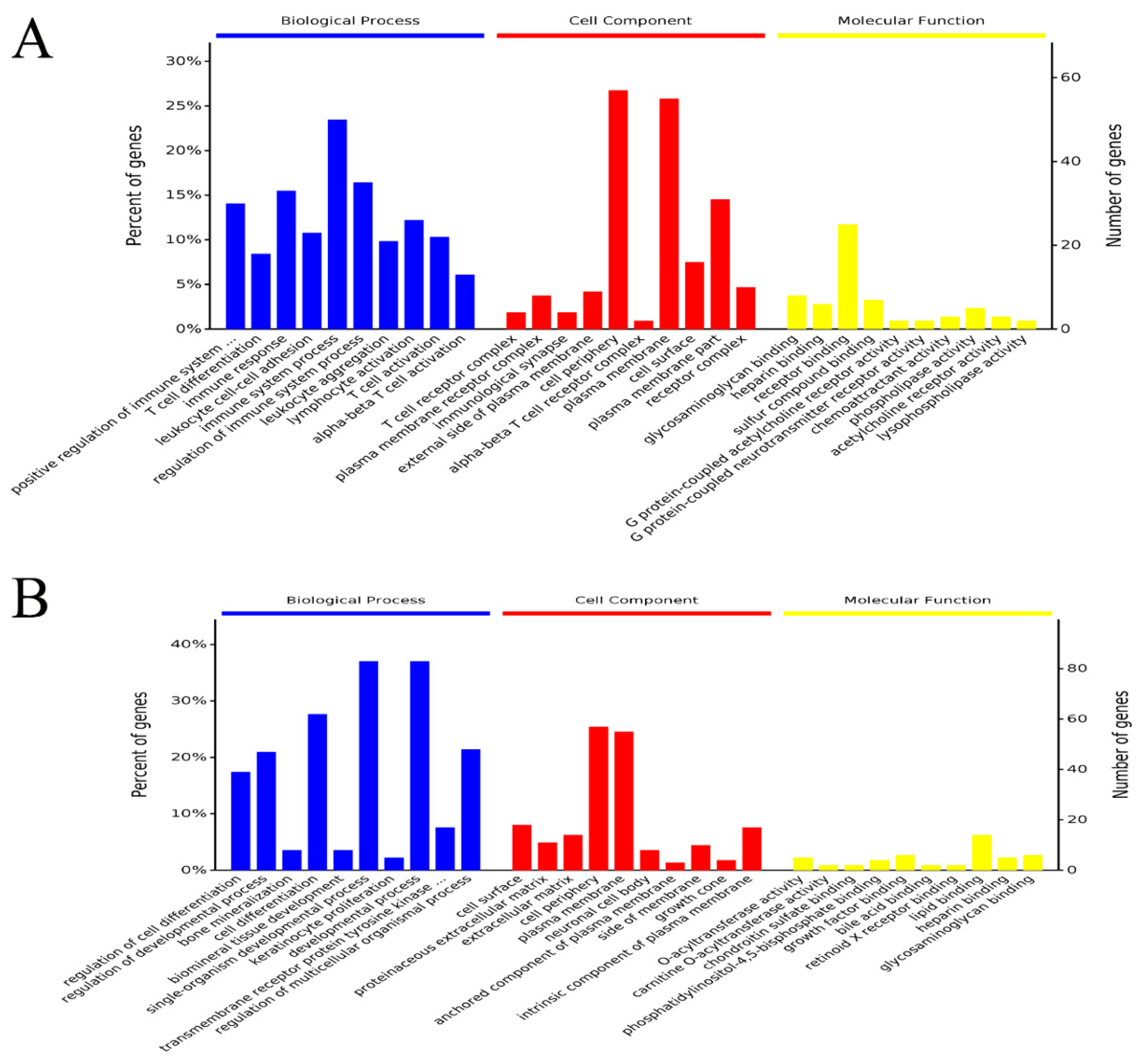
Gene ontology (GO) term enrichment analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis for the identified DEGs were performed using the free online platform Majorbio I-Sanger Cloud Platform (
www.i-sanger.com) to predict the metabolic pathways in which the DEGs are involved. The DEGs of GO term enrichment was classified into the categories of molecular functions, cellular component, and biological process. A corrected
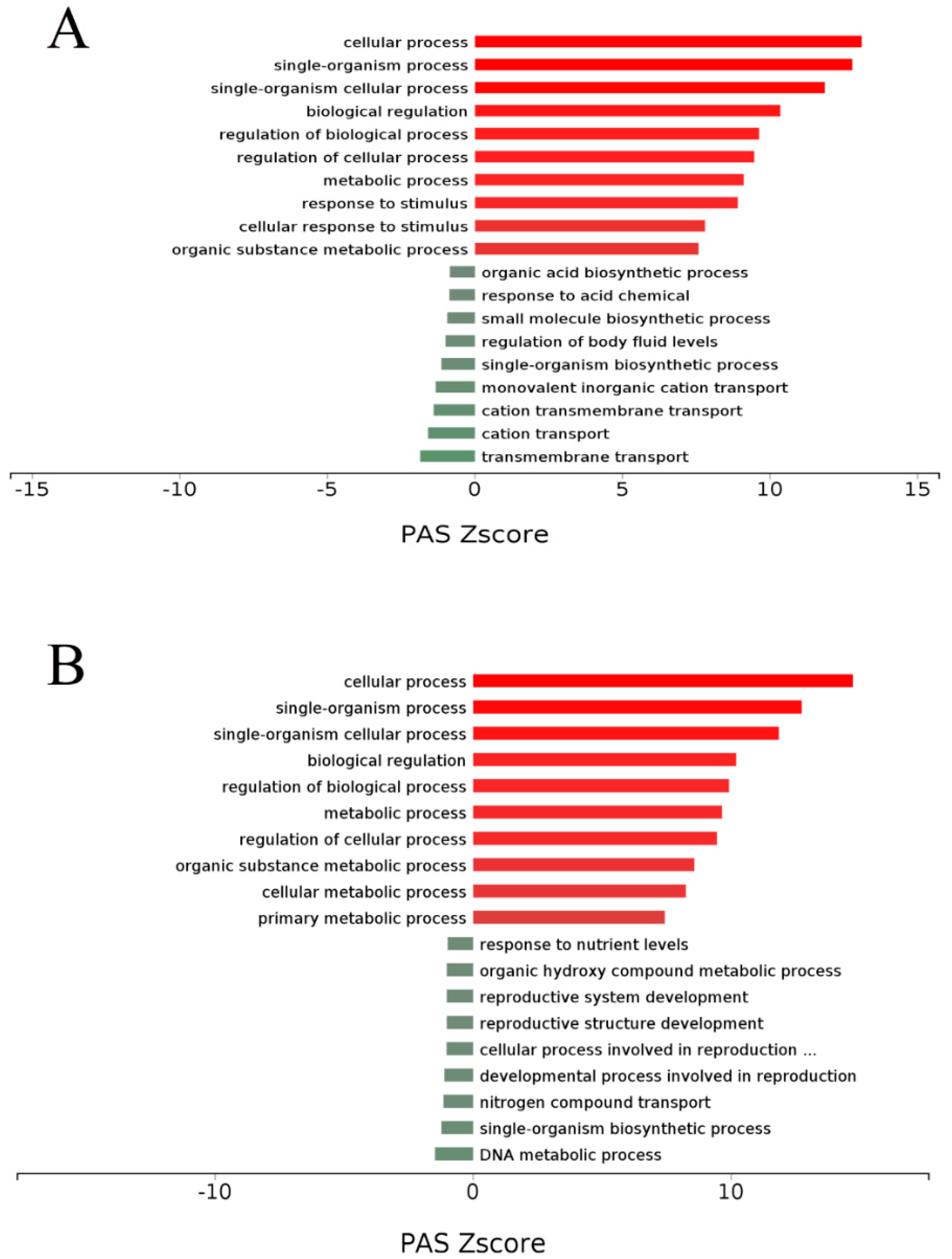
p-value < 0.05 indicated significant enrichment. Additionally, a complementary and more comprehensive pathway activation strength (PAS) value analysis of GO processes based on DEGs was performed using OmicsBean (
http://www.omicsbean.cn) to further identify the deep biological processes and pathways.
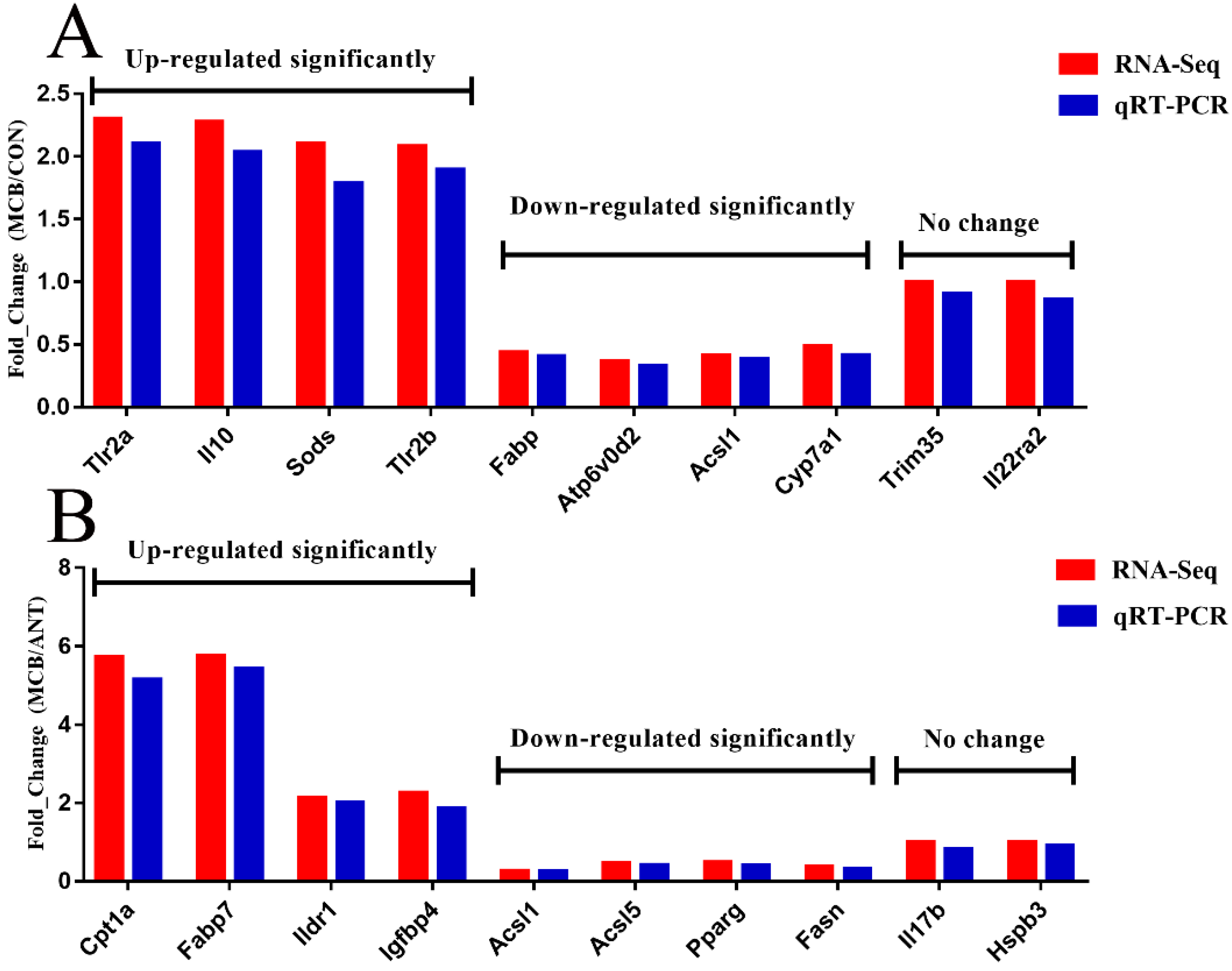
2.9. Confirmation of RNA-seq Data by qRT-PCR
To validate the accuracy of the RNA-seq data, we randomly selected 20 DEGs for qRT-PCR analysis. A total of six RNA samples from each group were reverse transcribed to complementary DNA using a Prime Script RT Master Mix Kit with DNase I (TaKaRa, Dalian, China) following the manufacturer’s instructions. Primers were designed using primer premier 5.0 software and were synthesized by Sangon Biotech Co., Ltd. (Shanghai, China) (
Table S2). Then, qRT-PCR was performed in triplicate reactions in 96 well plates using the Top Green qPCR SuperMix Kit (TransGen Biotech, Beijing, China) according to the manufacturer’s instructions using the following cycling conditions: 95 °C for 30 s, followed by 40 cycles of 95 °C for 10 s, and 55 °C for 42 s, then 72 °C for 10 s, followed by 72 °C for 5 min. A melting curve was then produced using an ABI QuantStudio 7 Flex Sequence Detection System (Applied Biosystems Co. Ltd., Foster City, CA, USA). The fold expression change of DEGs was calculated using the 2
−ΔΔCt method [
32]. The mRNA levels of the DEGs were normalized against an endogenous reference gene, glyceraldehyde-3-phosphate dehydrogenase (
Gapdh).
2.10. Statistical Analysis
The results are presented as the mean ± standard deviation (S.D.). The data were determined by one-way ANOVA followed by Duncan’s multiple comparisons between groups using SPSS25.0 software (SPSS, Chicago, IL, USA). p < 0.05 or less was considered statistically significant.
4. Discussion
C. butyricum is reported to have a variety of beneficial effects in humans and animals, including promoted growth performance [
33], improved meat quality [
22], enhanced immune function [
34], alleviated oxidative stress [
26], attenuated fatty liver disease [
25], and balanced intestinal microflora [
33]. In our study,
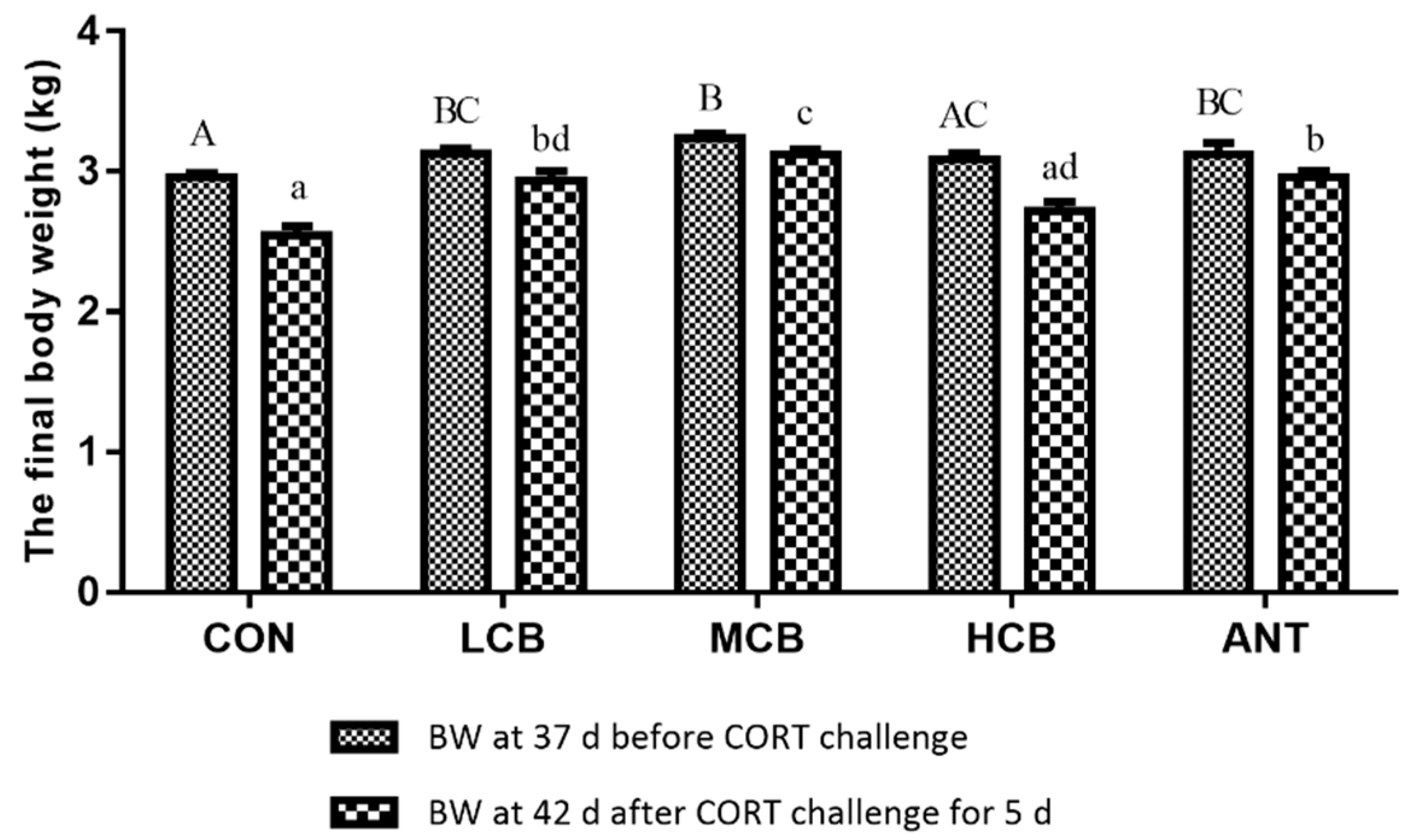
C. butyricum supplementation at 200mg/kg (LCB) and 400 mg/kg (MCB) both significantly increased the body weight of Pekin ducks at 37 d before CORT challenge. However,
C. butyricum supplementation at 600 mg/kg (HCB) did not significantly affect growth performance compared with the CON group, indicating that 600 mg/kg
C. butyricum does not positively promote the growth performance of Pekin ducks. Antibiotic supplementation also had positive effects on Pekin duck’s growth performance. Numerous studies have reported that sub-therapeutic use of antibiotics in diets can promote growth performance and control gastrointestinal bacterial infections in Pekin ducks [
35]. However, the use of antibiotics which has largely led to the emergence of resistant bacteria and the potential for producing drug residues in animal feed and products; therefore, a ban has been issued [
36].
C. butyricum is, therefore, an alternative to improve the growth performance of animals. Exogenous CORT administration significantly suppresses body weight in ducks [
2], which is consistent with the results of our study. In our study, the body weight of ducks at 42 d decreased after CORT challenge, but
C. butyricum intervention (especially 400 mg/kg) significantly attenuated this degree. Although the antibiotic presented the same protective effects as
C. butyricum, we recommend using
C. butyricum instead of antibiotics.
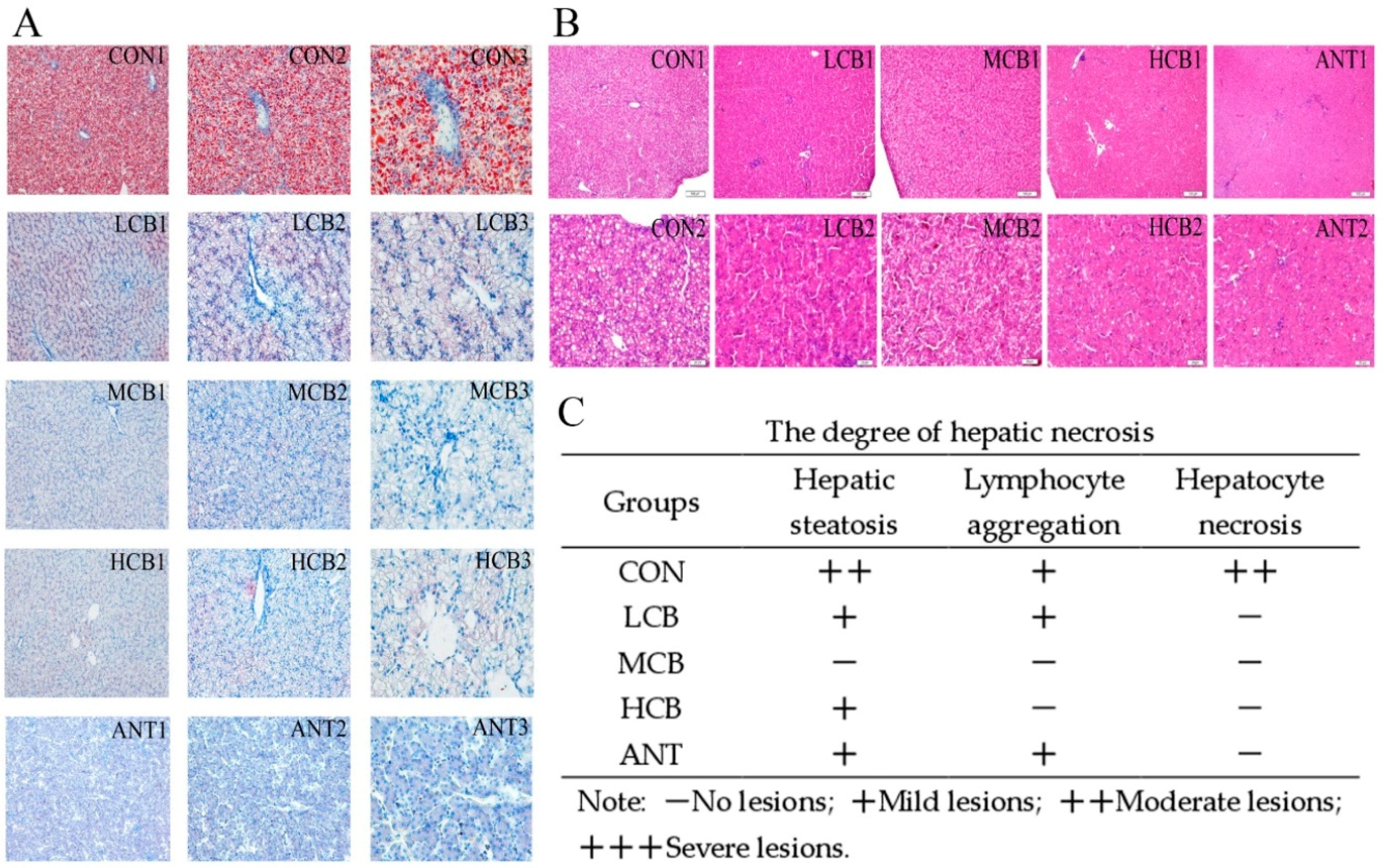
The liver is susceptible to CORT challenges, so it was selected as the research model. To further confirm the protective effects of
C. butyricum on hepatic function in Pekin ducks, we measured the serum levels of aminotransferase protein and pathological changes in the liver, which are classical parameters used to evaluate hepatic damage induced by CORT [
20]. The serum ALT level is used as a special parameter of hepatotoxic effects, while AST activity is considered a less specific biomarker of liver function. However, the elevated AST and ALT levels induced by CORT were drastically attenuated by
C. butyricum intervention, and the 400 mg/kg
C. butyricum intervention had the best effects. In addition,
C. butyricum intervention significantly alleviated the increase in serum ALB levels in MCB and HCB groups. Histological results showed that hepatic steatosis, lymphocyte aggregation and hepatocyte necrosis induced by CORT were also significantly ameliorated by
C. butyricum supplementation and the 400 mg/kg
C. butyricum intervention had the best effects.
C. butyricum supplementation has been demonstrated to alleviate CCl
4-induced liver lesions in mice [
25]. Meanwhile, other findings have revealed that
C. butyricum can effectively attenuate the neurohistopathological changes in dementia mice [
37], indicating that
C. butyricum supplementation indeed attenuates CORT-induced liver injury.
Oxidative stress is also an important mechanism of liver injury in animals that can involve increased lipid peroxidation and decreased activities of antioxidant enzymes [
38]. Our results revealed that
C. butyricum attenuated the increase in hepatic MDA and ROS contents and the decrease in SOD and GSH-PX activities induced by CORT in both MCB and HCB groups, indicating that a certain dose of
C. butyricum may ameliorate liver injury via an anti-oxidative mechanism. MDA has long been considered an indicator of oxidative stress, and the excess production of ROS can induce immunosuppression [
39], but
C. butyricum can enhance the activities of antioxidant enzymes to alleviate oxidative stress in animals. A previous study revealed that
C. butyricum supplementation can significantly increase the activities of SOD and CAT, and reduce the MDA level after CCl
4-induced liver injury in mice [
25]. Oxidative stress plays an important role in the pathogenesis of CORT [
40], and therefore, these key indices can be used to evaluate the status of oxidative stress and thus provide insight into identifying the mechanisms underlying the protective effects of
C. butyricum.
After a CORT challenge for five days, inflammatory reactions play a crucial role. Inflammatory cytokines, such as IL-1β, IL-6 and TNF-α, are associated with CORT-induced hepatic damage. In agreement with previous reports [
23], the pro-inflammatory factors were significantly enhanced after CORT challenge and were alleviated by 400 mg/kg
C. butyricum supplementation in our study. Moreover, anti-inflammatory factors (IL-4, IL-10 and TGF-β) that were decreased by CORT stimulation were significantly increased in the 400 mg/kg
C. butyricum-treated groups. The anti-inflammatory mechanism in this liver injury model is complicated, but it at least involves the attenuation of pro-inflammatory expression and an increase in the levels of IL-4, IL-10 and TGF-β.
Non-alcoholic fatty liver disease (NAFLD) is the result of hepatic steatosis because of lipid accumulation in the liver. Extensive efforts have been made to elucidate the mechanisms underlying hepatic steatosis, which include hepatic fatty acid uptake, de novo lipogenesis, β-oxidation and export [
41,
42]. Fatty liver formation in waterfowl appears very similar to human NAFLD, making Pekin ducks a very interesting model to better understand hepatic steatosis [
43,
44]. Previous publications reported that
C. butyricum supplementation alleviated hepatic lipid accumulation by enhancing fatty acid β-oxidation and inhibiting lipogenesis [
21,
25]. In a recent study,
C. butyricum attenuated NAFLD progression in broilers, confirming that
C. butyricum can protect hepatocytes against chronic damage [
45]. Of greater interest, the Oil red O stain showed that pre-administration of
C. butyricum to Pekin ducks receiving CORT significantly inhibited lipid accumulation in the liver and the 400 mg/kg
C. butyricum intervention showed the best effects. This result indicated that
C. butyricum can effectively prevent CORT-induced hepatic steatosis in Pekin ducks.
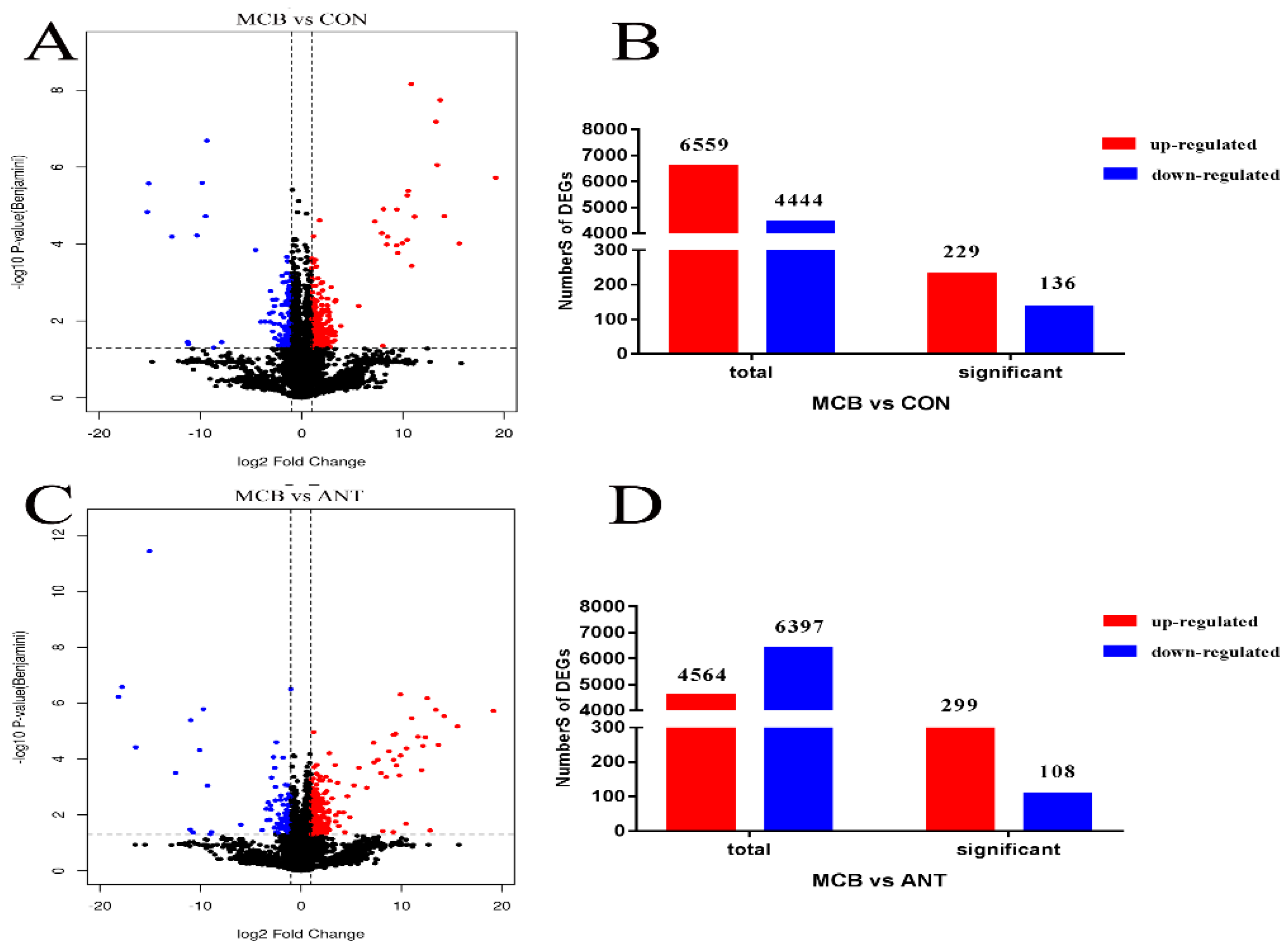
We demonstrated that
C. butyricum can attenuate CORT-induced oxidative stress, inflammatory responses and hepatic steatosis in Pekin ducks and the 400 mg/kg
C. butyricum intervention showed the best effects, however, the alleviated mechanisms involved require further exploration. Recently, Illumina transcriptome profiling, as an efficient, rapid method, has become widely used in stock raising [
46]. Our study analyzed the hepatic transcriptome of Pekin ducks with 400 mg/kg
C. butyricum intervention in order to explore the genes and pathways involved in the protective effects of
C. butyricum against CORT.
C. butyricum supplementation at 400 mg/kg activated the NF-κB, and toll-like receptor signaling pathways in the liver, thereby promoting lymphocyte development and differentiation, decreased the expression of genes related to pro-inflammatory responses, and increased the expression of genes related to anti-inflammatory responses (such as
Tlr2a,
Tlr2b and
Il10) and anti-oxidant capacity (
Sod3). Furthermore, the high level of
Alb expression in both MCB and ANT groups increased the serum ALB levels. These two signaling pathways can regulate inflammatory responses and cytokine secretion [
47,
48]. PI3K-Akt signaling pathway is involved in inflammation and immune regulation and was also significantly enriched in the liver and the
Il10r-β and
Il10r-α genes were enriched in the PI3K-Akt signaling pathway in the MCB group. IL-10 can promote T cell differentiation and proliferation and can induce the secretion of anti-inflammation cytokines, such as IL-4, IL-10, interferon (IFN)-γ, and TGF-β in cytotoxic T cells and NK cells [
49,
50]. Accordingly, dietary
C. butyricum supplementation up-regulated the transcription levels of genes involved in cytokine-cytokine receptor interaction (enriched genes:
Lifr,
Bmpr2,
Tnfsf10,
Ccl5,
Il8, and
Pdgfra). Therefore,
C. butyricum supplementation can improve the antioxidant capacity and inflammatory responses by enhancing the expression of related genes and the enrichment of related pathways to play a hepatoprotective role in Pekin ducks after CORT challenge.
Oil red O staining showed that a CORT challenge induced severe hepatic steatosis, while
C. butyricum supplementation decreased the accumulation of liver fat and alleviated the degree of hepatic steatosis. At the transcriptional level, we found the expression of
Acsl1,
Acsl5,
Cyp7a1,
Acaa2 and
Pck1 were down-regulated; whereas
Fabp7 and
Cpt1a were up-regulated in the MCB group. These genes were demonstrated to decrease gluconeogenesis and increase fatty acid β-oxidation, fatty acid transportation and cholesterol metabolism [
51,
52,
53]. These results indicated that
C. butyricum supplementation in MCB group can modulate hepatic lipid metabolism-related genes in Pekin ducks to alleviate the hepatic steatosis induced by CORT. According to the KEGG analysis, we found that the PPAR signaling pathway, which is associated with lipid metabolism regulation [
54], was activated in the
C. butyricum-treated group in Pekin ducks, indicating that providing
C. butyricum to Pekin ducks can regulate the expression of lipid metabolism-related genes through activation of the PPAR signaling pathway. In the PPAR signaling pathway, the enriched genes associated with lipid transport promoters (
Apoa1) and fatty acid oxidation promoters (
Fabp7,
Cpt1a, and
Cpt2) were up-regulated, whereas peroxisome proliferator activated receptor (
Ppar-γ), gluconeogenesis promoter (
Pck1), fatty acid transport promoter (
Acsl1/5 and
Lpl), lipogenesis (
Me1 and
Scd-1) and cholesterol metabolism (
Cyp7a1) were down-regulated in the
C. butyricum supplementation group compared with the CON and ANT groups.
KEGG pathway analysis revealed that
C. butyricum supplementation activated the glycerophospholipid metabolism signaling pathway and alleviated lipid accumulation, which is consistent with a previous report showing that
C. butyricum can reduce lipid accumulation in CCl
4-challenged mice [
25].
Pla2g3 was observed enriched in glycerophospholipid metabolism.
Pla2g3 is the rate-limiting enzyme in glycerophospholipid metabolism [
55] and is also involved in signal transduction, immune regulation and the inflammatory response [
56,
57]. Therefore,
C. butyricum supplementation modulated glycerophospholipid metabolism by significantly down-regulating
Pla2g3 expression in order to protect the liver from lipid accumulation. Additionally, the adipocytokine signaling pathway also clustered more DEGs (
Socs3,
Pck1,
Acsl5 and
Cpt1a) in the MCB and ANT groups, indicating that compared with antibiotics, feeding
C. butyricum to ducks may have a more obvious protective effect on alleviating lipid accumulation after CORT challenge. Additionally,
C. butyricum treatment significantly modulated the expression of genes related to cholesterol transporter activity (up-regulated
Abcg1, down-regulated
Cetp and
Stard5).
Abcg1 can increase the transmembrane transport of sterols and phospholipids [
58] and
Cetp mainly mediates the selective uptake of high density lipoprotein cholesterol by the liver [
59]. Therefore, moderate
C. butyricum intervention increased the transport and decreased the deposition of cholesterol in the liver, which may alleviate hepatic steatosis induced by CORT. In summary, we demonstrated that
C. butyricum intervention protects Pekin ducks from CORT-induced hepatic steatosis in the liver via the epigenetic modulation of lipogenic and lipophagic genes.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}