Response of Soybean to Hydrochar-Based Rhizobium Inoculation in Loamy Sandy Soil

 ,
, 
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil, Plant, Hydrochar and Rhizobia
2.2. Survival of Bradyrhizobium japonicum in HTC Char
2.3. Pot Experiments
2.4. Analysis of Plant Nutrients
2.5. Plant Growth Under Field Conditions
2.6. Statistical Analyses
3. Results
3.1. Survival of B. japonicum in the HTC Carrier
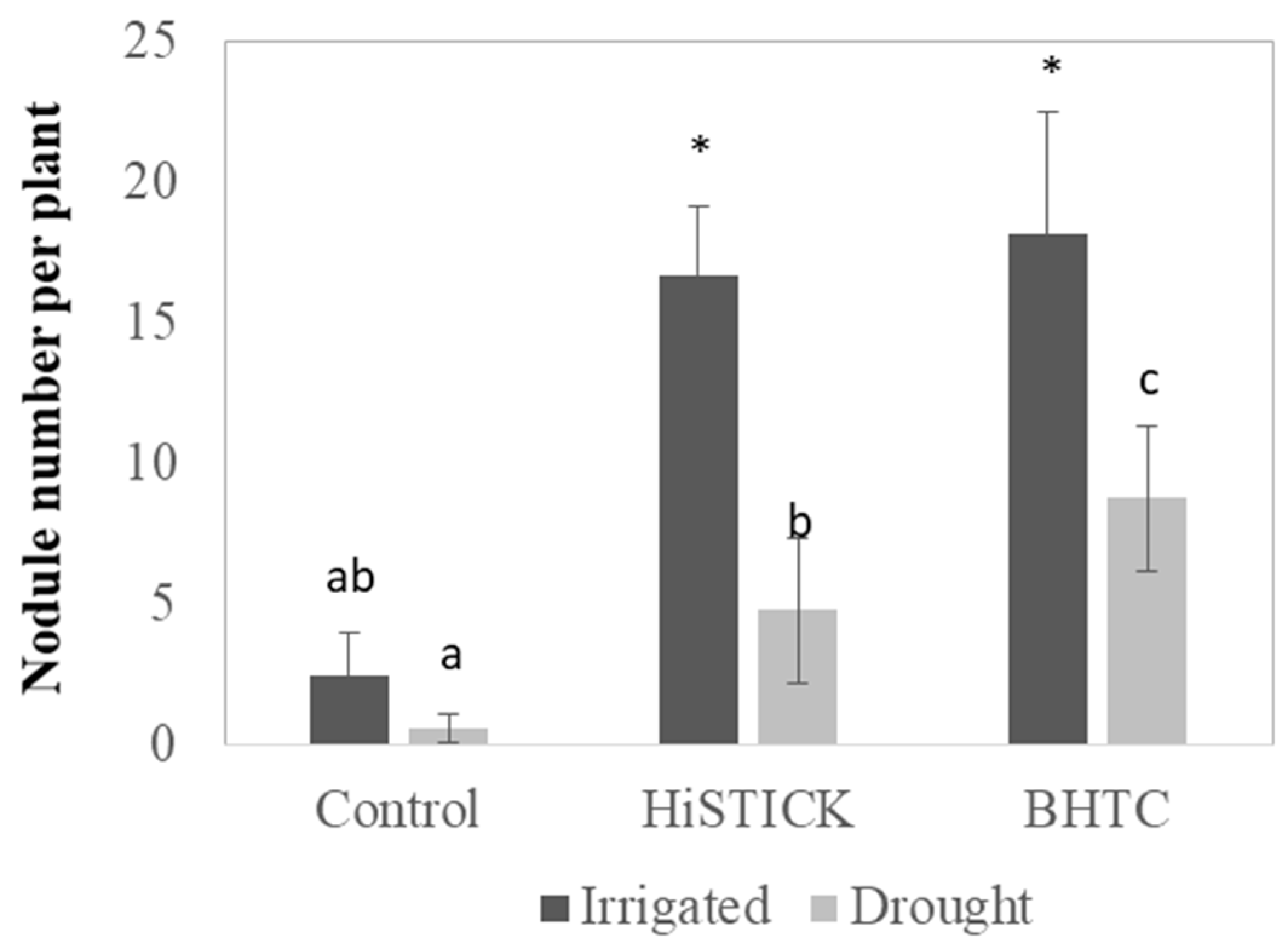
3.2. Plant Growth under Greenhouse Conditions
3.3. Soybean Growth under Field Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wilcox, J. World distribution and trade of soybean. In Soybean: Improvement, Production, and Uses; Boerma, H.R., Specht, J.E., Eds.; American Society of Agronomy: Madison, WI, USA, 2004; pp. 1–14. [Google Scholar]
- Vieira, R.F.; Mendes, I.C.; Reis-Junior, F.B.; Hungria, M. Symbiotic nitrogen fixation in tropical food grain legumes: Current status. In Microbes for Legume Improvement; Khan, M.S., Zaidi, A., Musarrat, J., Eds.; Springer: Vienna, Austria, 2010; pp. 427–472. [Google Scholar]
- Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Al-Huqail, A.A.; Wirth, S.; Egamberdieva, D. Comparing symbiotic performance and physiological responses of two soybean cultivars to arbuscular mycorrhizal fungi under salt stress. Saudi J. Biol. Sci. 2016, 26, 38–48. [Google Scholar] [CrossRef]
- Reckling, M.; Bergkvist, G.; Watson, C.A.; Stoddard, F.L.; Zander, P.M.; Walker, R.; Pristeri, A.; Toncea, I.; Bachinger, J. Trade-offs between economic and environmental impacts of introducing legumes into cropping systems. Front. Plant Sci. 2016, 7, 669. [Google Scholar] [CrossRef] [PubMed]
- Panzieri, M.; Marchettini, N.; Hallam, T.G. Importance of the Bradyrhizobium japonicum symbiosis for the sustainability of a soybean cultivation. Ecol. Model. 2000, 135, 301–310. [Google Scholar] [CrossRef]
- Ma, H.; Egamberdieva, D.; Wirth, S.; Li, Q.; Omari, R.A.; Hou, M.; Bellingrath-Kimura, S.D. Effect of biochar and irrigation on the interrelationships among soybean growth, root nodulation, plant p uptake, and soil nutrients in a sandy field. Sustainability 2019, 11, 6542. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Egamberdieva, D.; Wirth, S.; Bellingrath-Kimura, S.D. Effect of biochar and irrigation on soybean-rhizobium symbiotic performance and soil enzymatic activity in field rhizosphere. Agronomy 2019, 9, 626. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Navarro, D.N.; Margaret Oliver, I.; Albareda Contreras, M.; Ruiz-Sainz, J.E. Soybean interactions with soil microbes, agronomical and molecular aspects. Agron. Sustain. Dev. 2011, 31, 173–190. [Google Scholar] [CrossRef] [Green Version]
- Bouhmouch, I.; Souad-Mouhsine, B.; Brhada, F.; Aurag, J. Influence of host cultivars and Rhizobium species on the growth and symbiotic performance of Phaseolus vulgaris under salt stress. J. Plant Physiol. 2005, 162, 1103–1113. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D.; Li, L.; Lindström, K.; Räsänen, L. A synergistic interaction between salt tolerant Pseudomonas and Mezorhizobium strains improves growth and symbiotic performance of liquorice (Glycyrrhiza uralensis Fish.) under salt stress. Appl. Microbiol. Biotechnol. 2015, 100, 2829–2841. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D.; Davranov, K.; Wirth, S.; Hashem, A.; Abd_Allah, E.F. Impact of soil salinity on the plant-growth–promoting and biological control abilities of root associated bacteria. Saudi J. Biol. Sci. 2017, 24, 1601–1608. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Pandey, A.; Palni, L.M. Carrier-based preparations of plant growth-promoting bacterial inoculants suitable for use in cooler regions. World J. Microb. Biotechnol. 2005, 21, 941–945. [Google Scholar] [CrossRef]
- Abd El-Fattah, D.A.; Eweda, W.E.; Zayed, M.S.; Hassanein, M.K. Effect of carrier materials, sterilization method, and storage temperature on survival and biological activities of Azotobacter chroococcum inoculant. Ann. Agric. Sci. 2013, 58, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Tamreihao, K.; Ningthoujam, D.S.; Nimaichand, S.; Singh, E.S.; Reena, P.; Singh, S.H.; Nongthomba, U. Biocontrol and plant growth promoting activities of a Streptomyces corchorusii strain UCR3-16 and preparation of powder formulation for application as biofertilizer agents for rice plant. Microbiol. Res. 2016, 192, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Bashan, Y.; de-Bashan, L.E.; Prabhu, S.R.; Hernandez, J.P. Advances in plant growth-promoting bacterial inoculant technology: Formulations and practical perspectives (1998–2013). Plant Soil 2014, 378, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Ardakani, S.S.; Hedari, A.; Tayebi, L.; Mohammadi, M. Promotion of cotton seedlings growth characteristics by development and use of new bioformulations. Int. J. Bot. 2010, 6, 95–100. [Google Scholar] [CrossRef]
- Hynes, R.K.; Kraig, K.A.; Covert, D.; Smith, R.S.; Rennie, R.J. Liquid rhizobial inoculants for lentil and field pea. J. Prod. Agric. 1995, 8, 547–552. [Google Scholar] [CrossRef]
- Atieno, M.; Wilson, N.; Casteriano, A.; Crossett, B.; Lesueur, D.; Deaker, R. Aqueous peat extract exposes rhizobia to sub-lethal stress which may prime cells for improved desiccation tolerance. Appl. Microbiol. Biotechnol. 2018, 102, 7521–7539. [Google Scholar] [CrossRef]
- Ronchi, A.L.; Grassano, A.; Balatti, A.P. Perlite as a carrier for legume inocu-lants. Agrochimica 1997, 41, 186–195. [Google Scholar]
- Daza, A.; Santamarı, C.; Rodriguez-Navarro, D.N.; Camacho, M.; Orive, R.; Temprano, F. Perlite as a carrier for bacterial inoculants. Soil Biol. Biochem. 2000, 32, 567–572. [Google Scholar] [CrossRef]
- Albareda, M.; Dulce, N.; Navarro, R.; Camacho, M.; Tenprano, F.J. Alternatives to peat as carrier for rhizobia inoculants, solid and liquid formulations. Soil Biol. Biochem. 2008, 40, 2771–2779. [Google Scholar] [CrossRef]
- Libra, J.A.; Ro, K.S.; Kammann, C.; Funke, A.; Berge, N.D.; Neubauer, Y. Hydrothermal carbonization of biomass residuals: A comparative review of the chemistry, processes and applications of wet and dry pyrolysis. Biofuels 2011, 2, 71–106. [Google Scholar] [CrossRef] [Green Version]
- Egamberdieva, D.; Wirth, S.; Behrendt, U.; Abd Allah, E.F.; Berg, G. Biochar treatment resulted in a combined effect on soybean growth promotion and a shift in plant growth promoting rhizobacteria. Front. Microbiol. 2016, 7, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gai, C.; Chen, M.; Liu, T.; Peng, N.; Liu, Z. Gasification characteristics of hydrochar and pyrochar derived from sewage sludge. Energy 2016, 113, 957–965. [Google Scholar] [CrossRef]
- Gasco, G.; Paz-Ferreiro, J.; Alvarez, M.L.; Saa, A.; Mendez, A. Biochars and hydrochars prepared by pyrolysis and hydrothermal carbonisation of pig manure. Waste Manag. 2018, 79, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Sun, H.; Han, L.; Xue, L.; Chen, Y.; Yang, L.; Xing, B. Fabrication of hydrochar based on food waste (FWHTC) and its application in aqueous solution rare earth ions adsorptive removal: Process, mechanisms and disposal methodology. J. Clean. Prod. 2019, 212, 1423–1433. [Google Scholar] [CrossRef]
- Yu, O.Y.; Raichle, B.; Sink, S. Impact of biochar on the water holding capacity of loamy sand soil. Int. J. Energy Environ. Eng. 2013, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Feng, Y.; Xue, L.; Sun, H.; Han, L.; Yang, L.; Sun, Q.; Chu, Q. Biowaste to treasure: Application of microbial-aged hydrochar in rice paddy could improve nitrogen use efficiency and rice grain free amino acids. J. Clean. Prod. 2019, 240, 118180. [Google Scholar] [CrossRef]
- Ji, M.; Sang, W.; Tsang, D.C.W.; Usman, M.; Zhang, S.; Luo, G. Molecular and microbial insights towards understanding the effects of hydrochar on methane emission from paddy soil. Sci. Total Environ. 2020, 714, 136769. [Google Scholar] [CrossRef] [PubMed]
- Malghani, S.; Jüschke, E.; Baumert, J.; Thuille, A.; Antonietti, M.; Trumbore, S.; Gleixner, G. Carbon sequestration potential of hydrothermal carbonization char (hydrochar) in two contrasting soils; results of a 1-year field study. Biol. Fertil. Soils 2015, 51, 123–134. [Google Scholar] [CrossRef]
- Zimmer, S.; Messmer, M.; Haase, T.; Piepho, H.P.; Mindermann, A.; Schulz, H.; Habekuß, A.; Ordon, F.; Wilbois, K.-P.; Heß, J. Effects of soybean variety and Bradyrhizobium strains on yield, protein content and biological nitrogen fixation under cool growing conditions in Germany. Eur. J. Agron. 2016, 72, 38–46. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Reckling, M.; Wirth, S. Biochar-based inoculum of Bradyrhizobium sp. improves plant growth and yield of lupin (Lupinus albus L.) under drought stress. Eur. J. Soil Biol. 2017, 78, 38–42. [Google Scholar] [CrossRef]
- Reckling, M.; Bergkvist, G.; Watson, C.A.; Stoddard, F.L.; Bachinger, J. Re-designing organic grain legume cropping systems using systems agronomy. Eur. J. Agron. 2000, 112, 125951. [Google Scholar] [CrossRef]
- Brahmaprakash, G.P.; Sahu, P.K. Biofertilizers for Sustainability. J. Indian Inst. Sci. 2012, 92, 37–62. [Google Scholar]
- Egamberdieva, D.; Ma, H.; Reckling, M.; Wirth, S.; Bellingrath-Kimura, S.D. Potential effects of biochar-based microbial inoculants in agriculture. Environ. Sustain. 2018, 1, 19–24. [Google Scholar] [CrossRef]
- Tripti; Kumar, A.; Usmani, Z.; Kumar, V.; Anshumali. Biochar and flyash inoculated with plant growth promoting rhizobacteria act as potential biofertilizer for luxuriant growth and yield of tomato plant. J. Environ. Manag. 2017, 190, 20–27. [Google Scholar] [CrossRef]
- Saxena, J.; Rana, G.; Pandey, M. Impact of addition of biochar along with Bacillus sp. on growth and yield of French beans. Sci. Hortic. 2013, 162, 351–356. [Google Scholar] [CrossRef]
- Khavazi, K.; Rejali, F.; Seguin, P.; Miransari, M. Effects of carrier, sterilization method, and incubation on survival of Bradyrhizobium japonicum in soybean (Glycine max L.) inoculants. Enzym. Microb. Technol. 2007, 41, 780–784. [Google Scholar] [CrossRef]
- Głodowska, M.; Husk, B.; Schwinghamer, T.; Smith, D.L. Biochar is a growth-promoting alternative to peat moss for the inoculation of corn with a pseudomonad. Agron. Sustain. Dev. 2016, 36, 21. [Google Scholar] [CrossRef] [Green Version]
- Saranya, K.; Kumutha, K.; Krishnan, P.S. Influence of biochar and Azospirillum application on the growth of maize. Madras Agric. J. 2011, 98, 158–164. [Google Scholar]
- Chen, J.; Liu, X.; Zheng, J.; Zhang, B.; Lu, H.; Chi, Z.; Pan, G.; Li, L.; Zhen, J.; Zhang, X.; et al. Biochar soil amendment increased bacterial but decreased fungal gene abundance with shifts in community structure in a slightly acid rice paddy from Southwest China. Appl. Soil Ecol. 2013, 71, 33–44. [Google Scholar] [CrossRef]
- Li, W.; Zhang, X.; Xiong, X.; Zhang, B.; Wang, L. Determination of the best conditions for modified biochar immobilized petroleum hydrocarbon degradation microorganism by orthogonal test. IOP Conf. Ser. Earth Environ. Sci. 2017, 94, 012191. [Google Scholar] [CrossRef]
- Ghazi, A.A. Potential for biochar as an alternate carrier to peat moss for the preparation of Rhizobia bio inoculum. Microbiol. Res. J. Int. 2017, 18, 1–9. [Google Scholar] [CrossRef]
- Sangeetha, D. Survival of plant growth promoting bacterial inoculants in different carrier materials. Int. J. Pharm. Biol. Arch. 2002, 3, 170–178. [Google Scholar]
- Steiner, C.; Teixeira, W.G.; Lehmann, J.; Zech, W. Microbial response to charcoal amendments of highly weathered soils and Amazonian dark earths in Central Amazonia preliminary results. In Amazonian Dark Earths: Explorations in Space and Time; Glaser, B., Woods, W.I., Eds.; Springer: Heidelberg, Germany, 2004; pp. 195–212. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Watered | Drought | |||||
|---|---|---|---|---|---|---|
| N | P | K | N | P | K | |
| Control | 2.263c | 0.271a | 1.920b | 1.803a | 0.254a | 1.459a |
| HISTICK | 2.782d | 0.277a | 2.086b | 1.923ab | 0.270a | 1.511a |
| BHTC | 2.983d | 0.278a | 1.954b | 2.149bc | 0.285a | 1.502a |
| Greenhouse | N | P | K | Shoot | Root | Nodule |
|---|---|---|---|---|---|---|
| Treatments × Irrigation | ns | ns | ns | ns | ns | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Egamberdieva, D.; Ma, H.; Alimov, J.; Reckling, M.; Wirth, S.; Bellingrath-Kimura, S.D. Response of Soybean to Hydrochar-Based Rhizobium Inoculation in Loamy Sandy Soil. Microorganisms 2020, 8, 1674. https://doi.org/10.3390/microorganisms8111674
Egamberdieva D, Ma H, Alimov J, Reckling M, Wirth S, Bellingrath-Kimura SD. Response of Soybean to Hydrochar-Based Rhizobium Inoculation in Loamy Sandy Soil. Microorganisms. 2020; 8(11):1674. https://doi.org/10.3390/microorganisms8111674
Chicago/Turabian StyleEgamberdieva, Dilfuza, Hua Ma, Jakhongir Alimov, Moritz Reckling, Stephan Wirth, and Sonoko Dorothea Bellingrath-Kimura. 2020. "Response of Soybean to Hydrochar-Based Rhizobium Inoculation in Loamy Sandy Soil" Microorganisms 8, no. 11: 1674. https://doi.org/10.3390/microorganisms8111674
APA StyleEgamberdieva, D., Ma, H., Alimov, J., Reckling, M., Wirth, S., & Bellingrath-Kimura, S. D. (2020). Response of Soybean to Hydrochar-Based Rhizobium Inoculation in Loamy Sandy Soil. Microorganisms, 8(11), 1674. https://doi.org/10.3390/microorganisms8111674