Molecular Characteristics of Extraintestinal Pathogenic E. coli (ExPEC), Uropathogenic E. coli (UPEC), and Multidrug Resistant E. coli Isolated from Healthy Dogs in Spain. Whole Genome Sequencing of Canine ST372 Isolates and Comparison with Human Isolates Causing Extraintestinal Infections

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. E. coli Isolates
2.2. Phylogenetic Grouping
2.3. Serotyping
2.4. Multilocus Sequence Typing (MLST)
2.5. CH Typing
2.6. Virulence Genotyping
2.7. Antimicrobial Susceptibility and ESBL and pAmpC Typing
2.8. Whole Genome Sequencing (WGS)
2.9. Statistical Analysis
3. Results
3.1. Phylogenetic Groups of the 197 Canine Isolates
3.2. Virulence Factor (VF)-Encoding Genes in the 197 Canine Isolates
3.3. Antimicrobial Resistance in the 197 Canine Isolates
3.4. Sequence Types, Clones and Serotypes Displayed by the 91 Canine ExPEC and/or UPEC Isolates and 28 MDR Isolates
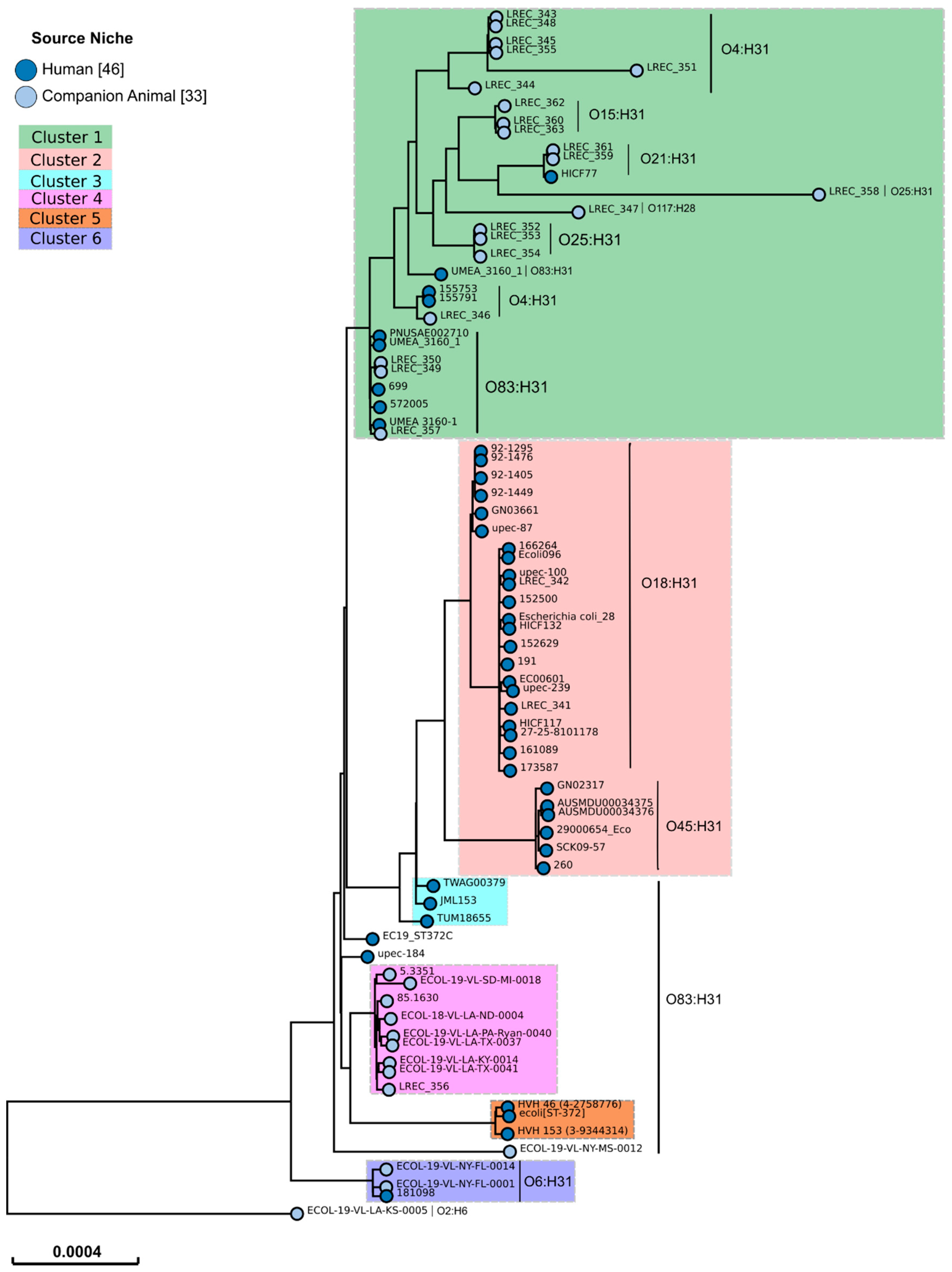
3.5. Whole Genome Sequencing (WGS) and Molecular Characterisation of ST372 Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Boehmer, T.; Vogler, A.J.; Thomas, A.; Sauer, S.; Hergenroether, M.; Straubinger, R.K.; Birdsell, D.; Keim, P.; Sahl, J.W.; Williamson, C.H.D.; et al. Phenotypic characterization and whole genome analysis of extended-spectrum beta-lactamase-producing bacteria isolated from dogs in Germany. PLoS ONE 2018, 13, e0206252. [Google Scholar] [CrossRef]
- Bourne, J.A.; Chong, W.L.; Gordon, D.M. Genetic structure, antimicrobial resistance and frequency of human associated Escherichia coli sequence types among faecal isolates from healthy dogs and cats living in Canberra, Australia. PLoS ONE 2019, 14, e0212867. [Google Scholar] [CrossRef] [Green Version]
- Flament-Simon, S.C.; Nicolas-Chanoine, M.H.; García, V.; Duprilot, M.; Mayer, N.; Alonso, M.P.; García-Meniño, I.; Blanco, J.E.; Blanco, M.; Blanco, J. Clonal structure, virulence factor-encoding genes and antibiotic resistance of Escherichia coli, causing urinary tract infections and other extraintestinal infections in humans in Spain and France during 2016. Antibiotics 2020, 9, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mamani, R.; Flament-Simon, S.C.; García, V.; Mora, A.; Alonso, M.P.; López, C.; García-Meniño, I.; Díaz-Jiménez, D.; Blanco, J.E.; Blanco, M.; et al. Sequence types, clonotypes, serotypes, and virotypes of extended-spectrum β-lactamase-producing Escherichia coli causing bacteraemia in a Spanish hospital over a 12-year period (2000 to 2011). Front. Microbiol. 2019, 10, 1530. [Google Scholar] [CrossRef] [Green Version]
- Thompson, M.F.; Litster, A.L.; Platell, J.L.; Trott, D.J. Canine bacterial urinary tract infections: New developments in old pathogens. Vet. J. 2011, 190, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Whittam, T.S.; Wolfe, M.L.; Wilson, R.A. Genetic relationships among Escherichia coli isolates causing urinary tract infections in humans and animals. Epidemiol. Infect. 1989, 102, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Porter, S.; Johnston, B.; Kuskowski, M.A.; Spurbeck, R.R.; Mobley, H.L.T.; Williamson, D.A. Host characteristics and bacterial traits predict experimental virulence for Escherichia coli bloodstream isolates from patients with urosepsis. Open Forum Infect. Dis. 2015, 2, ofv083. [Google Scholar] [CrossRef] [PubMed]
- Spurbeck, R.R.; Dinh, P.C.; Walk, S.T.; Stapleton, A.E.; Hooton, T.M.; Nolan, L.K.; Kim, K.S.; Johnson, J.R.; Mobley, H.L.T. Escherichia coli isolates that carry vat, fyuA, chuA, and yfcV efficiently colonize the urinary tract. Infect. Immun. 2012, 80, 4115–4122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallonen, T.; Brodrick, H.J.; Harris, S.R.; Corander, J.; Brown, N.M.; Martin, V.; Peacock, S.J.; Parkhill, J. Systematic longitudinal survey of invasive Escherichia coli in England demonstrates a stable population structure only transiently disturbed by the emergence of ST131. Genome Res. 2017, 27, 1437–1449. [Google Scholar] [CrossRef] [Green Version]
- Kidsley, A.K.; O’Dea, M.; Saputra, S.; Jordan, D.; Johnson, J.R.; Gordon, D.M.; Turni, C.; Djordjevic, S.P.; Abraham, S.; Trott, D.J. Genomic analysis of phylogenetic group B2 extraintestinal pathogenic E. coli causing infections in dogs in Australia. Vet. Microbiol. 2020, 248, 108783. [Google Scholar] [CrossRef] [PubMed]
- LeCuyer, T.E.; Byrne, B.A.; Daniels, J.B.; Diaz-Campos, D.V.; Hammac, G.K.; Miller, C.B.; Besser, T.E.; Davis, M.A. Population structure and antimicrobial resistance of canine uropathogenic Escherichia coli. J. Clin. Microbiol. 2018, 56, e00788-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manges, A.R.; Geum, H.M.; Guo, A.; Edens, T.J.; Fibke, C.D.; Pitout, J.D.D. Global extraintestinal pathogenic Escherichia coli (ExPEC) Lineages. Clin. Microbiol. Rev. 2019, 32, e00135-18. [Google Scholar] [CrossRef] [PubMed]
- Riley, L.W. Pandemic lineages of extraintestinal pathogenic Escherichia coli. Clin. Microbiol. Infect. 2014, 20, 380–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valat, C.; Drapeau, A.; Beurlet, S.; Bachy, V.; Boulouis, H.-J.; Pin, R.; Cazeau, G.; Madec, J.-Y.; Haenni, M. Pathogenic Escherichia coli in dogs reveals the predominance of ST372 and the human-associated ST73 extra-intestinal lineages. Front. Microbiol. 2020, 11, 580. [Google Scholar] [CrossRef] [PubMed]
- Zogg, A.L.; Zurfluh, K.; Schmitt, S.; Nüesch-Inderbinen, M.; Stephan, R. Antimicrobial resistance, multilocus sequence types and virulence profiles of ESBL producing and non-ESBL producing uropathogenic Escherichia coli isolated from cats and dogs in Switzerland. Vet. Microbiol. 2018, 216, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Clabots, C.; Kuskowski, M.A. Multiple-host sharing, long-term persistence, and virulence of Escherichia coli clones from human and animal household members. J. Clin. Microbiol. 2008, 46, 4078–4082. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Johnston, B.; Clabots, C.R.; Kuskowski, M.A.; Roberts, E.; DebRoy, C. Virulence genotypes and phylogenetic background of Escherichia coli serogroup O6 isolates from humans, dogs, and cats. J. Clin. Microbiol. 2008, 46, 417–422. [Google Scholar] [CrossRef] [Green Version]
- Platell, J.L.; Cobbold, R.N.; Johnson, J.R.; Heisig, A.; Heisig, P.; Clabots, C.; Kuskowski, M.A.; Trott, D.J. Commonality among fluoroquinolone-resistant sequence type ST131 extraintestinal Escherichia coli isolates from humans and companion animals in Australia. Antimicrob. Agents Chemother. 2011, 55, 3782–3787. [Google Scholar] [CrossRef] [Green Version]
- Platell, J.L.; Trott, D.J.; Johnson, J.R.; Heisig, P.; Heisig, A.; Clabots, C.R.; Johnston, B.; Cobbold, R.N. Prominence of an O75 clonal group (clonal complex 14) among non-ST131 fluoroquinolone-resistant Escherichia coli causing extraintestinal infections in humans and dogs in Australia. Antimicrob. Agents Chemother. 2012, 56, 3898–3904. [Google Scholar] [CrossRef] [Green Version]
- De Toro, M.; Fernández, J.; García, V.; Mora, A.; Blanco, J.; de la Cruz, F.; Rodicio, M.R. Whole genome sequencing, molecular typing and in vivo virulence of OXA-48-producing Escherichia coli isolates including ST131 H30-Rx, H22 and H41 subclones. Sci. Rep. 2017, 7, 12103. [Google Scholar] [CrossRef] [Green Version]
- Flament-Simon, S.C.; García, V.; Duprilot, M.; Mayer, N.; Alonso, M.P.; García-Meniño, I.; Blanco, J.E.; Blanco, M.; Nicolas-Chanoine, M.H.; Blanco, J. High Prevalence of ST131 subclades C2-H30Rx and C1-M27 among extended-spectrum β-lactamase-producing Escherichia coli causing human extraintestinal infections in patients from two hospitals of Spain and France during 2015. Front. Cell. Infect. Microbiol. 2020, 10, 125. [Google Scholar] [CrossRef] [PubMed]
- Jamborova, I.; Johnston, B.D.; Papousek, I.; Kachlikova, K.; Micenkova, L.; Clabots, C.; Skalova, A.; Chudejova, K.; Dolejska, M.; Literak, I.; et al. Extensive genetic commonality among wildlife, wastewater, community, and nosocomial isolates of Escherichia coli sequence type 131 (H30R1 and H30Rx subclones) that carry blaCTX-M-27 or blaCTX-M-15. Antimicrob. Agents Chemother. 2018, 62, e00519-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumura, Y.; Pitout, J.D.D.; Peirano, G.; DeVinney, R.; Noguchi, T.; Yamamoto, M.; Gomi, R.; Matsuda, T.; Nakano, S.; Nagao, M.; et al. Rapid identification of different Escherichia coli sequence type 131 clades. Antimicrob. Agents Chemother. 2017, 61, e00179-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolas-Chanoine, M.H.; Blanco, J.; Leflon-Guibout, V.; Demarty, R.; Alonso, M.P.; Canica, M.M.; Park, Y.-J.; Lavigne, J.-P.; Pitout, J.; Johnson, J.R. Intercontinental emergence of Escherichia coli clone O25:H4-ST131 producing CTX-M-15. J. Antimicrob. Chemother. 2008, 61, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Nicolas-Chanoine, M.H.; Bertrand, X.; Madec, J.Y. Escherichia coli ST131, an intriguing clonal Group. Clin. Microbiol. Rev. 2014, 27, 543–574. [Google Scholar] [CrossRef] [Green Version]
- Price, L.B.; Johnson, J.R.; Aziz, M.; Clabots, C.; Johnston, B.; Tchesnokova, V.; Nordstrom, L.; Billig, M.; Chattopadhyay, S.; Stegger, M.; et al. The epidemic of extended-spectrum-β-lactamase-producing Escherichia coli ST131 is driven by a single highly pathogenic subclone, H30-Rx. MBio 2013, 4, e00377-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belas, A.; Marques, C.; Aboim, C.; Pomba, C. Emergence of Escherichia coli ST131 H30/H30-Rx subclones in companion animals. J. Antimicrob. Chemother. 2018, 74, 266–269. [Google Scholar] [CrossRef] [Green Version]
- Bortolami, A.; Zendri, F.; Maciuca, E.I.; Wattret, A.; Ellis, C.; Schmidt, V.; Pinchbeck, G.; Timofte, D. Diversity, virulence, and clinical significance of extended-spectrum β-lactamase- and pAmpC-producing Escherichia coli from companion animals. Front. Microbiol. 2019, 10, 1260. [Google Scholar] [CrossRef] [Green Version]
- Kawamura, K.; Sugawara, T.; Matsuo, N.; Hayashi, K.; Norizuki, C.; Tamai, K.; Kondo, T.; Arakawa, Y. Spread of CTX-type extended-spectrum β-lactamase-producing Escherichia coli isolates of epidemic clone B2-O25-ST131 among dogs and cats in Japan. Microb. Drug Resist. 2017, 23, 1059–1066. [Google Scholar] [CrossRef]
- Maeyama, Y.; Taniguchi, Y.; Hayashi, W.; Ohsaki, Y.; Osaka, S.; Koide, S.; Tamai, K.; Nagano, Y.; Arakawa, Y.; Nagano, N. Prevalence of ESBL/AmpC genes and specific clones among the third-generation cephalosporin-resistant Enterobacteriaceae from canine and feline clinical specimens in Japan. Vet. Microbiol. 2018, 216, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Melo, L.C.; Haenni, M.; Saras, E.; Duprilot, M.; Nicolas-Chanoine, M.H.; Madec, J.Y. Emergence of the C1-M27 cluster in ST131 Escherichia coli from companion animals in France. J. Antimicrob. Chemother. 2019, 74, 3111–3113. [Google Scholar] [CrossRef]
- Bogaerts, P.; Huang, T.-D.; Bouchahrouf, W.; Bauraing, C.; Berhin, C.; El Garch, F.; Glupczynski, Y. Characterization of ESBL- and AmpC-producing Enterobacteriaceae from diseased companion animals in Europe. Microb. Drug Resist. 2015, 21, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, Z.; Zhang, Y.; Zhang, Z.; Lei, L.; Xia, Z. Increasing prevalence of ESBL-producing multidrug resistance Escherichia coli from diseased pets in Beijing, China from 2012 to 2017. Front. Microbiol. 2019, 10, 2852. [Google Scholar] [CrossRef]
- Falgenhauer, L.; Imirzalioglu, C.; Ghosh, H.; Gwozdzinski, K.; Schmiedel, J.; Gentil, K.; Bauerfeind, R.; Kämpfer, P.; Seifert, H.; Michael, G.B.; et al. Circulation of clonal populations of fluoroquinolone-resistant CTX-M-15-producing Escherichia coli ST410 in humans and animals in Germany. Int. J. Antimicrob. Agents 2016, 47, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Nakai, Y.; Kataoka, Y. Mechanisms of resistance to cephalosporin and emergence of O25b-ST131 clone harboring CTX-M-27 β-lactamase in extraintestinal pathogenic Escherichia coli from dogs and cats in Japan. Microbiol. Immunol. 2012, 56, 480–485. [Google Scholar] [CrossRef]
- Johnson, J.R.; Kuskowski, M.A.; Owens, K.; Clabots, C.; Singer, R.S. Virulence genotypes and phylogenetic background of fluoroquinolone-resistant and susceptible Escherichia coli urine isolates from dogs with urinary tract infection. Vet. Microbiol. 2009, 136, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, H.; Li, Y.; Hao, C. High prevalence of β-lactamase and plasmid-mediated quinolone resistance genes in extended-spectrum cephalosporin-resistant Escherichia coli from dogs in Shaanxi, China. Front. Microbiol. 2016, 7, 1843. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Thungrat, K.; Boothe, D.M. Occurrence of OXA-48 carbapenemase and other β-lactamase genes in ESBL-producing multidrug resistant Escherichia coli from dogs and cats in the United States, 2009–2013. Front. Microbiol. 2016, 7, 1057. [Google Scholar] [CrossRef] [Green Version]
- Marques, C.; Belas, A.; Franco, A.; Aboim, C.; Gama, L.T.; Pomba, C. Increase in antimicrobial resistance and emergence of major international high-risk clonal lineages in dogs and cats with urinary tract infection: 16 year retrospective study. J. Antimicrob. Chemother. 2018, 73, 377–384. [Google Scholar] [CrossRef] [Green Version]
- Pepin-Puget, L.; El Garch, F.; Bertrand, X.; Valot, B.; Hocquet, D. Genome analysis of Enterobacteriaceae with non-wild type susceptibility to third-generation cephalosporins recovered from diseased dogs and cats in Europe. Vet. Microbiol. 2020, 242, 108601. [Google Scholar] [CrossRef]
- Wagner, S.; Gally, D.L.; Argyle, S.A. Multidrug-resistant Escherichia coli from canine urinary tract infections tend to have commensal phylotypes, lower prevalence of virulence determinants and ampC-replicons. Vet. Microbiol. 2014, 169, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.L.C.; Shen, X.; Chalmers, G.; Reid-Smith, R.J.; Slavic, D.; Dick, H.; Boerlin, P. Prevalence and mechanisms of extended-spectrum cephalosporin resistance in clinical and fecal Enterobacteriaceae isolates from dogs in Ontario, Canada. Vet. Microbiol. 2018, 213, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Ukah, U.V.; Glass, M.; Avery, B.; Daignault, D.; Mulvey, M.R.; Reid-smith, R.J.; Parmley, E.J.; Portt, A.; Boerlin, P.; Manges, A.R. Risk factors for acquisition of multidrug-resistant Escherichia coli and development of community-acquired urinary tract infections. Epidemiol. Infect. 2018, 146, 46–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Guinée, P.A.M.; Jansen, W.H.; Wadström, T.; Sellwood, R. Escherichia coli associated with neonatal diarrhoea in piglets and calves. In Laboratory Diagnosis in Neonatal Calf and Pig Diarrhoea; Springer: Dordrecht, The Netherlands, 1981; pp. 126–162. [Google Scholar]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.J.; Ochman, H.; et al. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef] [Green Version]
- Tchesnokova, V.; Billig, M.; Chattopadhyay, S.; Linardopoulou, E.; Aprikian, P.; Roberts, P.L.; Skrivankova, V.; Johnston, B.; Gileva, A.; Igusheva, I.; et al. Predictive diagnostics for Escherichia coli infections based on the clonal association of antimicrobial resistance and clinical outcome. J. Clin. Microbiol. 2013, 51, 2991–2999. [Google Scholar] [CrossRef] [Green Version]
- Roer, L.; Johannesen, T.B.; Hansen, F.; Stegger, M.; Tchesnokova, V.; Sokurenko, E.; Garibay, N.; Allesøe, R.; Thomsen, M.C.F.; Lund, O.; et al. CHTyper, a web tool for subtyping of extraintestinal pathogenic Escherichia coli based on the fumC and fimH alleles. J. Clin. Microbiol. 2018, 56, e00063-18. [Google Scholar] [CrossRef] [Green Version]
- Dahbi, G.; Mora, A.; Mamani, R.; López, C.; Alonso, M.P.; Marzoa, J.; Blanco, M.; Herrera, A.; Viso, S.; García-Garrote, F.; et al. Molecular epidemiology and virulence of Escherichia coli O16:H5-ST131: Comparison with H30 and H30-Rx subclones of O25b:H4-ST131. Int. J. Med. Microbiol. 2014, 304, 1247–1257. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; CLSI supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Blanco, M.; Alonso, M.P.; Nicolas-Chanoine, M.-H.; Dahbi, G.; Mora, A.; Blanco, J.E.; López, C.; Cortés, P.; Llagostera, M.; Leflon-Guibout, V.; et al. Molecular epidemiology of Escherichia coli producing extended-spectrum β-lactamases in Lugo (Spain): Dissemination of clone O25b:H4-ST131 producing CTX-M-15. J. Antimicrob. Chemother. 2009, 63, 1135–1141. [Google Scholar] [CrossRef] [Green Version]
- Leflon-Guibout, V.; Jurand, C.; Bonacorsi, S.; Espinasse, F.; Guelfi, M.C.; Duportail, F.; Heym, B.; Bingen, E.; Nicolas-Chanoine, M.H. Emergence and spread of three clonally related virulent isolates of CTX-M-15-producing Escherichia coli with variable resistance to aminoglycosides and tetracycline in a French geriatric hospital. Antimicrob. Agents Chemother. 2004, 48, 3736–3742. [Google Scholar] [CrossRef] [Green Version]
- Perez-Perez, F.J.; Hanson, N.D. Detection of plasmid-mediated AmpC-lactamase genes in clinical isolates by using multiplex PCR. J. Clin. Microbiol. 2002, 40, 2153–2162. [Google Scholar] [CrossRef] [Green Version]
- Flament-Simon, S.C.; de Toro, M.; Mora, A.; García, V.; García-Meniño, I.; Díaz-Jiménez, D.; Herrera, A.; Blanco, J. Whole genome sequencing and characteristics of mcr-1–harboring plasmids of porcine Escherichia coli isolates belonging to the high-risk clone O25b:H4-ST131 clade B. Front. Microbiol. 2020, 11, 387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanza, V.F.; de Toro, M.; Garcillán-Barcia, M.P.; Mora, A.; Blanco, J.; Coque, T.M.; de la Cruz, F. Plasmid flux in Escherichia coli ST131 sublineages, analyzed by plasmid constellation network (PLACNET), a new method for plasmid reconstruction from whole genome sequences. PLoS Genet. 2014, 10, e1004766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vielva, L.; de Toro, M.; Lanza, V.F.; de la Cruz, F. PLACNETw: A web-based tool for plasmid reconstruction from bacterial genomes. Bioinformatics 2017, 33, 3796–3798. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Alvarado, A.; Garcillán-Barcia, M.P.; de la Cruz, F. A degenerate primer MOB typing (DPMT) method to classify gamma-proteobacterial plasmids in clinical and environmental settings. PLoS ONE 2012, 7, e40438. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Zhou, Z.; Alikhan, N.-F.; Mohamed, K.; Fan, Y.; Achtman, M. The EnteroBase user’s guide, with case studies on Salmonella transmissions, Yersinia pestis phylogeny, and Escherichia core genomic diversity. Genome Res. 2020, 30, 138–152. [Google Scholar] [CrossRef] [Green Version]
- Flament-Simon, S.C.; Duprilot, M.; Mayer, N.; García, V.; Alonso, M.P.; Blanco, J.; Nicolas-Chanoine, M.H. Association between kinetics of early biofilm formation and clonal lineage in Escherichia coli. Front. Microbiol. 2019, 10, 1183. [Google Scholar] [CrossRef] [Green Version]
- Abreu-Salinas, F.; Díaz-Jiménez, D.; García-Meniño, I.; Lumbreras, P.; López-Beceiro, A.M.; Fidalgo, L.E.; Rodicio, M.R.; Mora, A.; Fernández, J. High prevalence and diversity of cephalosporin-resistant Enterobacteriaceae including extraintestinal pathogenic E. coli CC648 lineage in rural and urban dogs in Northwest Spain. Antibiotics 2020, 9, 468. [Google Scholar] [CrossRef] [PubMed]
- Belas, A.; Salazar, A.S.; Gama, L.T.d.; Couto, N.; Pomba, C. Risk factors for faecal colonisation with Escherichia coli producing extended-spectrum and plasmid-mediated AmpC-lactamases in dogs. Vet. Rec. 2014, 175, 202. [Google Scholar] [CrossRef] [PubMed]
- Damborg, P.; Morsing, M.K.; Petersen, T.; Bortolaia, V.; Guardabassi, L. CTX-M-1 and CTX-M-15-producing Escherichia coli in dog faeces from public gardens. Acta Vet. Scand. 2015, 57, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupouy, V.; Abdelli, M.; Moyano, G.; Arpaillange, N.; Bibbal, D.; Cadiergues, M.-C.; Lopez-Pulin, D.; Sayah-Jeanne, S.; de Gunzburg, J.; Saint-Lu, N.; et al. Prevalence of beta-lactam and quinolone/fluoroquinolone resistance in Enterobacteriaceae from dogs in France and Spain—Characterization of ESBL/pAmpC isolates, genes, and conjugative plasmids. Front. Vet. Sci. 2019, 6, 279. [Google Scholar] [CrossRef] [Green Version]
- Haenni, M.; Saras, E.; Métayer, V.; Médaille, C.; Madec, J.-Y. High prevalence of blaCTX-M-1 /IncI1/ST3 and blaCMY-2 /IncI1/ST2 plasmids in healthy urban dogs in France. Antimicrob. Agents Chemother. 2014, 58, 5358–5362. [Google Scholar] [CrossRef] [Green Version]
- Hansen, K.H.; Bortolaia, V.; Nielsen, C.A.; Nielsen, J.B.; Schønning, K.; Agersø, Y.; Guardabassi, L. Host-specific patterns of genetic diversity among IncI1-Iγ and IncK plasmids encoding CMY-2 β-lactamase in Escherichia coli isolates from humans, poultry meat, poultry, and dogs in Denmark. Appl. Environ. Microbiol. 2016, 82, 4705–4714. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.S.; Song, W.; Park, H.-M.; Oh, J.-Y.; Chae, J.-C.; Shin, S.; Jeong, S.H. Clonal spread of extended-spectrum cephalosporin-resistant Enterobacteriaceae between companion animals and humans in South Korea. Front. Microbiol. 2019, 10, 1371. [Google Scholar] [CrossRef] [Green Version]
- Hordijk, J.; Schoormans, A.; Kwakernaak, M.; Duim, B.; Broens, E.; Dierikx, C.; Mevius, D.; Wagenaar, J.A. High prevalence of fecal carriage of extended-spectrum β-lactamase/AmpC-producing Enterobacteriaceae in cats and dogs. Front. Microbiol. 2013, 4, 242. [Google Scholar] [CrossRef] [Green Version]
- Karkaba, A.; Hill, K.; Benschop, J.; Pleydell, E.; Grinberg, A. Carriage and population genetics of extended spectrum β-lactamase-producing Escherichia coli in cats and dogs in New Zealand. Vet. Microbiol. 2019, 233, 61–67. [Google Scholar] [CrossRef]
- Liakopoulos, A.; Betts, J.; La Ragione, R.; van Essen-Zandbergen, A.; Ceccarelli, D.; Petinaki, E.; Koutinas, C.K.; Mevius, D.J. Occurrence and characterization of extended-spectrum cephalosporin-resistant Enterobacteriaceae in healthy household dogs in Greece. J. Med. Microbiol. 2018, 67, 931–935. [Google Scholar] [CrossRef]
- Melo, L.C.; Oresco, C.; Leigue, L.; Netto, H.M.; Melville, P.A.; Benites, N.R.; Saras, E.; Haenni, M.; Lincopan, N.; Madec, J.Y. Prevalence and molecular features of ESBL/pAmpC-producing Enterobacteriaceae in healthy and diseased companion animals in Brazil. Vet. Microbiol. 2018, 221, 59–66. [Google Scholar] [CrossRef]
- Pires, J.; Bernasconi, O.J.; Kasraian, S.; Hilty, M.; Perreten, V.; Endimiani, A. Intestinal colonisation with extended-spectrum cephalosporin-resistant Escherichia coli in Swiss pets: Molecular features, risk factors and transmission with owners. Int. J. Antimicrob. Agents 2016, 48, 759–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha-Gracia, R.C.; Cortés-Cortés, G.; Lozano-Zarain, P.; Bello, F.; Martínez-Laguna, Y.; Torres, C. Faecal Escherichia coli isolates from healthy dogs harbour CTX-M-15 and CMY-2 β-lactamases. Vet. J. 2015, 203, 315–319. [Google Scholar] [CrossRef]
- Schaufler, K.; Bethe, A.; Lübke-Becker, A.; Ewers, C.; Kohn, B.; Wieler, L.H.; Guenther, S. Putative connection between zoonotic multiresistant extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli in dog feces from a veterinary campus and clinical isolates from dogs. Infect. Ecol. Epidemiol. 2015, 5, 25334. [Google Scholar] [CrossRef] [Green Version]
- Seni, J.; Falgenhauer, L.; Simeo, N.; Mirambo, M.M.; Imirzalioglu, C.; Matee, M.; Rweyemamu, M.; Chakraborty, T.; Mshana, S.E. Multiple ESBL-producing Escherichia coli sequence types carrying quinolone and aminoglycoside resistance genes circulating in companion and domestic farm animals in Mwanza, Tanzania, harbor commonly occurring plasmids. Front. Microbiol. 2016, 7, 142. [Google Scholar] [CrossRef]
- Sevilla, E.; Mainar-Jaime, R.C.; Moreno, B.; Martín-Burriel, I.; Morales, M.; Andrés-Lasheras, S.; Chirino-Trejo, M.; Badiola, J.J.; Bolea, R. Antimicrobial resistance among canine enteric Escherichia coli isolates and prevalence of attaching–effacing and extraintestinal pathogenic virulence factors in Spain. Acta Vet. Hung. 2020, 68, 1–7. [Google Scholar] [CrossRef]
- Suay-García, B.; Galán, F.; Rodríguez-Iglesias, M.A.; Pérez-Gracia, M.T. Detection and characterization of extended-spectrum beta-lactamases-producing Escherichia coli in animals. Vector Borne Zoonotic Dis. 2019, 19, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Umeda, K.; Hase, A.; Matsuo, M.; Horimoto, T.; Ogasawara, J. Prevalence and genetic characterization of cephalosporin-resistant Enterobacteriaceae among dogs and cats in an animal shelter. J. Med. Microbiol. 2019, 68, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Wedley, A.L.; Dawson, S.; Maddox, T.W.; Coyne, K.P.; Pinchbeck, G.L.; Clegg, P.; Nuttall, T.; Kirchner, M.; Williams, N.J. Carriage of antimicrobial resistant Escherichia coli in dogs: Prevalence, associated risk factors and molecular characteristics. Vet. Microbiol. 2017, 199, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Adler, A.; Gniadkowski, M.; Baraniak, A.; Izdebski, R.; Fiett, J.; Hryniewicz, W.; Malhotra-Kumar, S.; Goossens, H.; Lammens, C.; Lerman, Y.; et al. Transmission dynamics of ESBL-producing Escherichia coli clones in rehabilitation wards at a tertiary care centre. Clin. Microbiol. Infect. 2012, 18, E497–E505. [Google Scholar] [CrossRef] [Green Version]
- Izdebski, R.; Baraniak, A.; Fiett, J.; Adler, A.; Kazma, M.; Salomon, J.; Lawrence, C.; Rossini, A.; Salvia, A.; Vidal Samso, J.; et al. Clonal structure, extended-spectrum β-lactamases, and acquired AmpC-type cephalosporinases of Escherichia coli populations colonizing patients in rehabilitation centers in four countries. Antimicrob. Agents Chemother. 2013, 57, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Dierikx, C.M.; van Duijkeren, E.; Schoormans, A.H.W.; van Essen-Zandbergen, A.; Veldman, K.; Kant, A.; Huijsdens, X.W.; van der Zwaluw, K.; Wagenaar, J.A.; Mevius, D.J. Occurrence and characteristics of extended-spectrum-β-lactamase- and AmpC-producing clinical isolates derived from companion animals and horses. J. Antimicrob. Chemother. 2012, 67, 1368–1374. [Google Scholar] [CrossRef] [PubMed]
- Ewers, C.; Bethe, A.; Stamm, I.; Grobbel, M.; Kopp, P.A.; Guerra, B.; Stubbe, M.; Doi, Y.; Zong, Z.; Kola, A.; et al. CTX-M-15-D-ST648 Escherichia coli from companion animals and horses: Another pandemic clone combining multiresistance and extraintestinal virulence? J. Antimicrob. Chemother. 2014, 69, 1224–1230. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Liu, H.; Li, Y.; Hao, C. Association between virulence profile and fluoroquinolone resistance in Escherichia coli isolated from dogs and cats in China. J. Infect. Dev. Ctries. 2017, 11, 306–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loncaric, I.; Misic, D.; Szostak, M.P.; Künzel, F.; Schäfer-Somi, S.; Spergser, J. Broad-spectrum cephalosporin-resistant and/or fluoroquinolone-resistant Enterobacterales associated with canine and feline urogenital infections. Antibiotics 2020, 9, 387. [Google Scholar] [CrossRef] [PubMed]
- Kidsley, A.K.; White, R.T.; Beatson, S.A.; Saputra, S.; Schembri, M.A.; Gordon, D.; Johnson, J.R.; O’Dea, M.; Mollinger, J.L.; Abraham, S.; et al. Companion animals are spillover hosts of the multidrug-resistant human extraintestinal Escherichia coli pandemic clones ST131 and ST1193. Front. Microbiol. 2020, 11, 1968. [Google Scholar] [CrossRef]
- Wang, J.; Huang, X.-Y.; Xia, Y.-B.; Guo, Z.-W.; Ma, Z.-B.; Yi, M.-Y.; Lv, L.-C.; Lu, P.-L.; Yan, J.-C.; Huang, J.-W.; et al. Clonal spread of Escherichia coli ST93 carrying mcr-1-harboring IncN1-IncHI2/ST3 plasmid among companion animals, China. Front. Microbiol. 2018, 9, 2989. [Google Scholar] [CrossRef]
- Pulss, S.; Stolle, I.; Stamm, I.; Leidner, U.; Heydel, C.; Semmler, T.; Prenger-Berninghoff, E.; Ewers, C. Multispecies and clonal dissemination of OXA-48 carbapenemase in Enterobacteriaceae from companion animals in Germany, 2009–2016. Front. Microbiol. 2018, 9, 1265. [Google Scholar] [CrossRef]
- Melo, L.C.; Boisson, M.N.G.; Saras, E.; Médaille, C.; Boulouis, H.-J.; Madec, J.-Y.; Haenni, M. OXA-48-producing ST372 Escherichia coli in a French dog. J. Antimicrob. Chemother. 2016, 72, 1256–1258. [Google Scholar] [CrossRef] [Green Version]
- Schubert, S.; Rakin, A.; Karch, H.; Carniel, E.; Heesemann, J. Prevalence of the “High-Pathogenicity Island” of Yersinia species among Escherichia coli strains that are pathogenic to humans. Infect. Immun. 1998, 66, 480–485. [Google Scholar] [CrossRef] [Green Version]
- Dobrindt, U.; Hochhut, B.; Hentschel, U.; Hacker, J. Genomic islands in pathogenic and environmental microorganisms. Nat. Rev. Microbiol. 2004, 2, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Denamur, E.; Clermont, O.; Bonacorsi, S.; Gordon, D. The population genetics of pathogenic Escherichia coli. Nature Rev. Microbiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Desvaux, M.; Dalmasso, G.; Beyrouthy, R.; Barnich, N.; Delmas, J.; Bonnet, R. Pathogenicity factors of genomic islands in intestinal and extraintestinal Escherichia coli. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]


{kind=link}
| VF-Encoding Gene | Number of Isolates (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Total (n = 197) | A (n = 32) | B1 (n = 26) | B2 (n = 84) | C (n = 10) | D (n = 6) | E (n = 14) | F (n = 18) | Clade V (n = 1) | NT 1 (n = 6) | |
| Adhesins | ||||||||||
| fimH | 192 (97.5) | 29 | 24 | 84 | 10 | 6 | 14 | 18 | 1 | 6 |
| fimAvMT78 | 20 (10.2) | 12 | 1 | 1 | 0 | 0 | 2 | 3 | 0 | 1 |
| papAH | 62 (31.5) | 2 | 3 | 54 | 0 | 1 | 1 | 1 | 0 | 0 |
| papC | 62 (31.5) | 2 | 3 | 54 | 0 | 1 | 1 | 1 | 0 | 0 |
| sfa/focDE | 66 (33.5) | 0 | 2 | 63 | 0 | 0 | 0 | 1 | 0 | 0 |
| afa/draBC | 1 (0.5) | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| yfcV | 96 (48.7) | 4 | 0 | 80 | 0 | 0 | 0 | 11 | 0 | 1 |
| Toxins | ||||||||||
| sat | 3 (1.5) | 0 | 0 | 2 | 0 | 0 | 0 | 1 | 0 | 0 |
| cnf1 | 54 (27.4) | 0 | 0 | 53 | 0 | 0 | 0 | 1 | 0 | 0 |
| hlyA | 56 (28.4) | 0 | 2 | 54 | 0 | 0 | 0 | 0 | 0 | 0 |
| cdtB | 8 (4.1) | 0 | 0 | 6 | 0 | 0 | 0 | 2 | 0 | 0 |
| tsh | 21 (10.7) | 2 | 2 | 0 | 8 | 0 | 2 | 6 | 0 | 1 |
| vat | 79 (40.1) | 1 | 0 | 76 | 0 | 0 | 0 | 2 | 0 | 0 |
| Iron uptake | ||||||||||
| iutA | 33 (16.8) | 3 | 3 | 7 | 8 | 1 | 2 | 9 | 0 | 0 |
| iroN | 93 (47.2) | 3 | 6 | 66 | 8 | 0 | 2 | 8 | 0 | 0 |
| fyuA | 106 (53.8) | 7 | 2 | 77 | 8 | 5 | 1 | 5 | 0 | 1 |
| chuA | 122 (61.9) | 0 | 0 | 84 | 0 | 6 | 14 | 18 | 0 | 0 |
| Capsule | ||||||||||
| kpsM II | 64 (32.5) | 1 | 0 | 41 | 0 | 4 | 4 | 10 | 1 | 3 |
| neuC-K1 | 12 (6.1) | 0 | 0 | 10 | 0 | 0 | 0 | 2 | 0 | 0 |
| kpsM II-K2 | 7 (3.6) | 0 | 0 | 2 | 0 | 3 | 0 | 1 | 0 | 1 |
| kpsM II-K5 | 45 (22.8) | 1 | 0 | 29 | 0 | 1 | 4 | 7 | 1 | 2 |
| kpsM III | 6 (3.0) | 1 | 0 | 2 | 0 | 2 | 1 | 0 | 0 | 0 |
| Miscellaneous | ||||||||||
| cvaC | 21 (10.7) | 1 | 4 | 2 | 8 | 0 | 0 | 6 | 0 | 0 |
| iss | 31 (15.7) | 2 | 5 | 6 | 8 | 0 | 2 | 8 | 0 | 0 |
| traT | 59 (29.9) | 2 | 12 | 17 | 8 | 2 | 7 | 10 | 0 | 1 |
| ibeA | 54 (27.4) | 0 | 0 | 49 | 0 | 0 | 0 | 3 | 0 | 2 |
| malX | 93 (47.2) | 0 | 5 | 79 | 0 | 0 | 0 | 7 | 0 | 2 |
| usp | 86 (43.7) | 1 | 3 | 77 | 0 | 1 | 1 | 2 | 0 | 1 |
| ExPEC status | 74 (37.6) | 2 | 2 | 61 | 0 | 1 | 1 | 7 | 0 | 0 |
| UPEC status | 82 (41.6) | 1 | 0 | 77 | 0 | 0 | 0 | 4 | 0 | 0 |
| Range of VFs | 0 to 18 | 0 to 10 | 1 to 10 | 2 to 17 | 1 to 8 | 5 to 7 | 2 to 7 | 3 to 18 | 3 | 1 to 9 |
| Mean of VFs | 7.87 | 2.31 | 2.96 | 12.79 | 6.60 | 5.67 | 4.14 | 7.94 | 3.00 | 3.67 |
| VF-Encoding Gene | Number of Isolates | ||||||
|---|---|---|---|---|---|---|---|
| B2-ST12 (n = 9) | D-ST38 (n = 4) | B2-ST73 (n = 4) | B2-ST127 (n = 8) | B2-ST141 (n = 5) | B2-ST372 (n = 29) | F-ST648 (n = 6) | |
| Adhesins | |||||||
| fimH | 9 | 4 | 4 | 8 | 5 | 29 | 6 |
| fimAvMT78 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| papAH | 9 | 0 | 4 | 7 | 1 | 21 | 0 |
| papC | 9 | 0 | 4 | 7 | 1 | 21 | 0 |
| sfa/focDE | 9 | 0 | 3 | 8 | 4 | 26 | 1 |
| afa/draBC | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| yfcV | 9 | 0 | 4 | 8 | 5 | 29 | 6 |
| Toxins | |||||||
| sat | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| cnf1 | 8 | 0 | 4 | 7 | 0 | 23 | 0 |
| hlyA | 9 | 0 | 4 | 7 | 1 | 23 | 0 |
| cdtB | 0 | 0 | 2 | 0 | 0 | 0 | 1 |
| tsh | 0 | 0 | 0 | 0 | 0 | 0 | 5 |
| vat | 8 | 0 | 4 | 8 | 5 | 29 | 2 |
| Iron uptake | |||||||
| iutA | 1 | 0 | 0 | 1 | 0 | 0 | 6 |
| iroN | 9 | 0 | 2 | 7 | 4 | 26 | 5 |
| fyuA | 8 | 4 | 4 | 8 | 5 | 29 | 3 |
| chuA | 9 | 4 | 4 | 8 | 5 | 29 | 6 |
| Capsule | |||||||
| kpsM II | 6 | 4 | 4 | 7 | 5 | 2 | 6 |
| neuC-K1 | 0 | 0 | 0 | 0 | 5 | 0 | 1 |
| kpsM II-K2 | 0 | 3 | 0 | 0 | 0 | 0 | 1 |
| kpsM II-K5 | 6 | 1 | 4 | 7 | 0 | 2 | 4 |
| kpsM III | 2 | 0 | 0 | 0 | 0 | 0 | 0 |
| Miscellaneous | |||||||
| cvaC | 0 | 0 | 0 | 0 | 0 | 0 | 5 |
| iss | 0 | 0 | 0 | 1 | 0 | 0 | 5 |
| traT | 1 | 1 | 2 | 1 | 0 | 4 | 6 |
| ibeA | 0 | 0 | 0 | 0 | 4 | 29 | 1 |
| malX | 9 | 0 | 4 | 8 | 5 | 29 | 2 |
| usp | 9 | 1 | 4 | 7 | 5 | 29 | 1 |
| ExPEC status | 9 | 0 | 3 | 8 | 4 | 20 | 6 |
| UPEC status | 8 | 0 | 3 | 8 | 5 | 29 | 3 |
| Range of VFs | 12 to 16 | 5 to 7 | 14 to 16 | 12 to 17 | 10 to 14 | 8 to 16 | 9 to 18 |
| Mean of VFs | 14.40 | 5.50 | 15.30 | 14.40 | 12.00 | 13.10 | 12.17 |
| Clone of Canine ExPEC and/or UPEC Isolates | Clone-Related Serotype of Canine ExPEC and/or UPEC Isolates (Number of Isolates) | Number of Human ExPEC and/or UPEC Isolates with Same Clone of Canine Isolates (49 of 261) | Number of Human ExPEC and/or UPEC Isolates with Same Clone and Serotype of Canine Isolates (31 of 261) |
|---|---|---|---|
| A-CH11-NEG-ST93 | O5:H4 (1) | 3 | 0 |
| A-CH11-27-ST new 1 | O4:H27 (1) | 0 | 0 |
| B1-CH4-27-ST58 | O8:H25 (1), O9:H25 (1) | 0 | 0 |
| B2-CH13-5-ST12 | O4:HNM (1), O18:H5 (1) | 0 | 0 |
| B2-CH13-7-ST12 | O4:H1 (1), O4:HNM (1) | 0 | 0 |
| B2-CH13-130-ST12 | O18:H5 (1) | 0 | 0 |
| B2-CH13-223-ST12 | O18:H5 (2) | 1 | 0 |
| B2-CH13-430-ST12 | O4:H5 (1) | 0 | 0 |
| B2-CH13-431-ST12 | O4:H5 (1) | 0 | 0 |
| B2-CH24-9-ST73 | O120:H31 (1) | 0 | 0 |
| B2-CH24-27-ST73 | O6:H1 (1) | 1 | 1 |
| B2-CH24-30-ST73 | O6:H1 (1) | 4 | 3 |
| B2-CH24-103-ST73 | O6:H1 (1) | 6 | 4 |
| B2-CH24-1-ST80 | O75:H7 (1) | 0 | 0 |
| B2-CH38-30-ST95 | O1:H7 (1) | 1 | 0 |
| B2-CH14-2-ST127 | O6:HNM (2) | 4 | 3 |
| B2-CH14-180-ST127 | O6:HNM (3), O6:H11 (1) | 1 | 1 |
| B2-CH14-fimHTRnew 1-ST127 | O6:H31 (1), O6:HNM (1) | 0 | 0 |
| B2-CH40-NEG-ST131 | O25:H4 (1) | 0 | 0 |
| B2-CH52-5-ST141 | O2:H6 (1) | 13 | 9 |
| B2-CH52-14-ST141 | O2:H6 (4) | 2 | 2 |
| B2-CH103-9-ST372 | O4:H31 (9), O15:H31 (1), O21:H31 (3), O25:H31 (4), O83:H31 (7), O117:H28 (1) | 2 | 2 |
| B2-CH103-10-ST372 | O15:H31 (1) | 0 | 0 |
| B2-CH103-17-ST372 | O15:H31 (1) | 0 | 0 |
| B2-CH103-240-ST372 | O83:H31 (1) | 0 | 0 |
| B2-CH103-706-ST372 | O83:H31 (1) | 0 | 0 |
| B2-CH96-433-ST646 | O22:HNM (1) | 0 | 0 |
| B2-CH43-fimHTRnew 2-ST929 | O138:H14 (1) | 0 | 0 |
| B2-CH13-175-ST961 | O4:HNM (1) | 0 | 0 |
| B2-CH52-428-ST998 | O2:H6 (1) | 0 | 0 |
| B2-CH14-64-ST1193 | O75:HNM (2) | 4 | 3 |
| B2-CH363-75-ST2622 | O83:H6 (1) | 0 | 0 |
| B2-CH195-2-ST5644 | O175:H5 (1) | 0 | 0 |
| B2-CH13-fimHTRnew 3-ST new 2 | O18:HNM (1) | 0 | 0 |
| B2-CH13-429-ST new 3 | O4:H5 (1) | 0 | 0 |
| B2-CH103-9-ST new 4 | O4:H31 (1) | 0 | 0 |
| B2-CH103-12-ST new 5 | O6:HNM (1) | 0 | 0 |
| B2-CH11-34-ST new 6 | O5:H11 (1) | 0 | 0 |
| B2-CH40-20-ST new 7 | O1:H4 (1) | 0 | 0 |
| B2-CH363-75-ST new 8 | O83:H4 (1) | 0 | 0 |
| B2-CH24-2-ST new 9 | ONT:H1 (1) | 0 | 0 |
| B2-CH24-1473-ST new 10 | O120:H5 (1) | 0 | 0 |
| B2-CH23-31-ST new 11 | O103:H4 (1) | 0 | 0 |
| B2-CH40-20-ST new 12 | O1:H4 (1) | 0 | 0 |
| B2-CH40-21-ST new 13 | O13:H4 (1) | 0 | 0 |
| D-CH35-27-ST new 14 | O77:H18 (1) | 0 | 0 |
| E-CH132-65-ST501 | ONT:H1 (1) | 0 | 0 |
| F-CH32-41-ST59 | O1:H7 (1) | 4 | 3 |
| F-CH4-27-ST648 | O4:H6 (1) | 1 | 0 |
| F-CH4-58-ST648 | O153:H42 (5) | 2 | 0 |
| Clone of Canine MDR Isolates | Clone-Related Serotype of Canine MDR Isolates (Number of Isolates) | Type of ESBL and pAmpC Enzymes Produced by Canine MDR Isolates | Number of Human E. coli Isolates with Same Clone of Canine MDR Isolates (Number and Type ESBL Produced by Human Isolates) (35 of 394) | Number of Human E. coli Isolates with Same Clone and Serotype of Canine MDR Isolates (1 of 394) |
|---|---|---|---|---|
| A-CH11-54-ST10 | O128:HNM (1) | SHV12 | 10 (3 SHV12) | 0 |
| A-CH11-NEG-ST93 | O5:H4 (1) | none | 4 (1 CTX-M-14) | 0 |
| A-CH11-54-ST8953 | O101:HNM (1) | CMY-2 | 0 | 0 |
| A-CH11-27-ST new 1 | O4:H27 (1) | none | 0 | 0 |
| B1-CH4-27-ST58 | O8:H25 (1), O9:H25 (1) | none | 4 (1 CTX-M-14 and 1 CTX-M-32) | 1 |
| B1-CH4-121-ST155 | O5:H11 (1) | none | 0 | 0 |
| B1-CH4-366-ST155 | O9:H10 (1) | CMY-2 | 1 (1 CTX-M-1) | 0 |
| B1-CH4-425-ST new 15 | O123:H11 (1) | CTX-M-1 | 0 | 0 |
| B1-CH4-31-ST new 16 | O8:H7 (1) | CTX-M-1 | 0 | 0 |
| B1-CH29-38-ST new 17 | O8:H49 (1) | CTX-M-1 | 0 | 0 |
| B1-CH30-38-ST new 18 | O12:H8 (1) | CMY-2 | 0 | 0 |
| B2-CH13-223-ST12 | O18:H5 (1) | CMY-2 | 1 | 0 |
| B2-CH13-429-ST new 3 | O4:H5 (1) | CMY-2 | 0 | 0 |
| C-CH4-39-ST88 | O45:HNM (1) | CTX-M-1 | 11 (1 CTX-M-14) | 0 |
| D-CH26-5-ST38 | O86:H18 (3) | CTX-M-14 | 0 | 0 |
| D-CH26-65-ST38 | O1:H34 (1) | CMY-2 | 2 (1 CTX-M-15) | 0 |
| D-CH35-27-ST new 14 | O77:H18 (1) | none | 0 | 0 |
| E-CH31-54-ST57 | O27:H40 (1) | CMY-2 | 1 | 0 |
| E-CH11-167-ST695 | O99:H38 (1) | none | 0 | 0 |
| E-CH4-31-ST1011 | O166:H45 (1) | CTX-M-55 | 0 | 0 |
| E-CH23-221-ST1140 | O44:H39 (1) | none | 0 | 0 |
| E-CH485-426-ST3774 | O9:H31 (1) | CMY-2 | 0 | 0 |
| F-CH88-145-ST457 | O11:H25 (2) | CMY-2 | 0 | 0 |
| F-CH4-27-ST648 | O4:H6 (1) | CTX-M-14 | 1 (1 CTX-M-15) | 0 |
| Cluster | Number of Strains (%) | p-Value 1 | |
|---|---|---|---|
| Canine (n = 151) | Human (n = 46) | ||
| 1 | 138 (91.4) | 9 (19.6) | <0.00001 |
| 2 | 0 | 28 (60.9) | <0.00001 |
| 3 | 0 | 3 (6.5) | 0.01209 |
| 4 | 9 (6.0) | 0 | |
| 5 | 0 | 3 (6.5) | 0.01209 |
| 6 | 2 (1.3) | 1 (2.2) | |
| Undefined | 2 (1.3) | 2 (4.3) | |
| Cluster (Number of Strains) | Number of Strains (%) | p-Value 1 Europe vs. North America | Countries (Number of Strains) | |
|---|---|---|---|---|
| Europe (n = 46) | North America (n = 143) | |||
| 1 (n = 147) | 30 (65.2) | 117 (81.8) | 0.02476 | USA (109), Spain (20), Canada (6), UK (4), Sweden (3), France (2), Germany (1), North America (2) |
| 2 (n = 28) | 11 (23.9) | 13 (9.1) | 0.01233 | USA (13), UK (8), Spain (1), France (1), The Netherlands (1), Australia (2), unknown (2) |
| 3 (n = 3) | 0 | 0 | Japan (1), Kenya (1), unknown (1) | |
| 4 (n = 8) | 1 (2.2) | 7 (4.9) | USA (7), Spain (1), unknown (1) | |
| 5 (n = 3) | 3 (6.5) | 0 | 0.01371 | Denmark (2), France (1) |
| 6 (n = 3) | 1 (2.2) | 2 (1.4) | USA (2), UK (1) | |
| Undefined (n = 5) | 0 | 4 (2.8) | USA (4), Australia (1) | |
| VF-Encoding Gene | Number of Strains (%) | p-Value 1 Canine vs. Human | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Canine (n =151) | Human (n = 46) | Cluster 1 (n = 147) | Cluster 2 (n = 28) | Cluster 3 (n = 3) | Cluster 4 (n = 9) | Cluster 5 (n = 3) | Cluster 6 (n = 3) | Undefined (n = 4) | ||
| Adhesins | ||||||||||
| fimH | 151 (100) | 46 (100) | 147 | 28 | 3 | 9 | 3 | 3 | 4 | |
| papAH | 88 (58) | 5 (11) | 89 | 0 | 0 | 0 | 0 | 3 | 1 | <0.00001 |
| papC | 113 (75) | 9 (20) | 113 | 0 | 0 | 0 | 3 | 3 | 3 | <0.00001 |
| papEF | 112 (74) | 9 (20) | 113 | 0 | 0 | 0 | 3 | 2 | 3 | <0.00001 |
| sfaDE | 3 (2) | 0 (0) | 1 | 0 | 0 | 0 | 0 | 2 | 0 | |
| sfaS | 6 (4) | 0 (0) | 6 | 0 | 0 | 0 | 0 | 0 | 0 | |
| focCD | 136 (90) | 12 (26) | 144 | 0 | 0 | 0 | 1 | 1 | 2 | <0.00001 |
| focG | 138 (91) | 12 (26) | 144 | 0 | 0 | 0 | 1 | 3 | 2 | <0.00001 |
| afaBCD/draP | 0 | 0 (0) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| yfcV | 150 (99) | 46 (100) | 146 | 28 | 3 | 9 | 3 | 3 | 4 | |
| Toxins | ||||||||||
| sat | 0 | 0 (0) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| cnf1 | 117 (77) | 11 (24) | 120 | 0 | 0 | 0 | 3 | 3 | 2 | <0.00001 |
| hlyA | 117 (77) | 11 (24) | 119 | 0 | 0 | 0 | 3 | 3 | 3 | <0.00001 |
| hlyF | 1 (1) | 3 (7) | 1 | 0 | 0 | 0 | 3 | 0 | 0 | 0.04034 |
| cdtB | 0 | 0 (0) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| vat | 148 (98) | 45 (98) | 144 | 27 | 3 | 9 | 3 | 3 | 4 | |
| Iron uptake | ||||||||||
| iutA | 1 (1) | 3 (7) | 1 | 0 | 0 | 0 | 3 | 0 | 0 | 0.04034 |
| iroN | 139 (92) | 14 (30) | 144 | 0 | 0 | 0 | 3 | 3 | 3 | <0.00001 |
| fyuA | 151 (100) | 46 (100) | 147 | 28 | 3 | 9 | 3 | 3 | 4 | |
| chuA | 151 (100) | 46 (100) | 147 | 28 | 3 | 9 | 3 | 3 | 4 | |
| ireA | 5 (3) | 1 (2) | 5 | 0 | 0 | 0 | 0 | 0 | 1 | |
| Capsule | ||||||||||
| kpsM II | 16 (11) | 36 (78) | 5 | 28 | 3 | 9 | 3 | 0 | 4 | <0.00001 |
| kpsM II-K1 | 0 (0) | 0 (0) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| kpsM II-K2 | 0 (0) | 0 (0) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| kpsM II-K5 | 16 (11) | 36 (78) | 5 | 28 | 3 | 9 | 3 | 0 | 4 | <0.00001 |
| Miscellaneous | ||||||||||
| iss1 | 1 (1) | 3 (7) | 1 | 0 | 0 | 0 | 3 | 0 | 0 | 0.04034 |
| iss2 | 140 (93) | 45 (98) | 136 | 27 | 3 | 9 | 3 | 3 | 4 | |
| traT | 34 (23) | 10 (22) | 25 | 5 | 0 | 9 | 3 | 1 | 1 | |
| ibeA | 149 (99) | 46 (100) | 146 | 28 | 3 | 9 | 3 | 3 | 3 | |
| malX-PAI | 150 (99) | 46 (100) | 146 | 28 | 3 | 9 | 3 | 3 | 4 | |
| usp | 151 (100) | 46 (100) | 147 | 28 | 3 | 9 | 3 | 3 | 4 | |
| ompT | 151 (100) | 46 (100) | 147 | 28 | 3 | 9 | 3 | 3 | 4 | |
| ExPEC status | 113 (75) | 10 (22) | 113 | 0 | 1 | 0 | 3 | 3 | 3 | <0.00001 |
| UPEC status | 151 (100) | 46 (100) | 147 | 28 | 3 | 9 | 3 | 3 | 4 | |
| Mean of VFs | 16.79 | 13.76 | 16.93 | 12.11 | 12.00 | 13.00 | 21.67 | 18.00 | 17.00 | |
| Serotype In Silico | Number of Strains (%) | p-Value 1 Canine vs. Human | Number of Isolates Belonging to Cluster | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Canine (n = 151) | Human (n = 46) | |||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | Undefined | ||||
| O2:H6 | 1 (0.7) | 0 | 1 | |||||||
| O4:H31 | 32 (21.2) | 3 (6.5) | 0.02631 | 35 | ||||||
| O6:H31 | 2 (1.3) | 1 (2.2) | 3 | |||||||
| O15:H31 | 30 (19.9) | 0 | 0.00062 | 30 | ||||||
| O18:H31 | 0 | 20 (43.5) | <0.00001 | 20 | ||||||
| O21:H14 | 5 (3.3) | 0 | 5 | |||||||
| O21:H31 | 4 (2.6) | 1 (2.2) | 5 | |||||||
| O25:H31 | 4 (2.6) | 0 | 4 | |||||||
| O45:H31 | 0 | 6 (13.0) | 0.00012 | 6 | ||||||
| O75:H31 | 5 (3.3) | 0 | 5 | |||||||
| O83:H31 | 58 (38.4) | 13 (28.3) | 53 | 3 | 9 | 3 | 3 | |||
| O117:H28 | 6 (4.0) | 0 | 6 | |||||||
| O-unknown:H31 | 3 (2.0) | 2 (4.3) | 3 | 2 | ||||||
| O-unknown:H28 | 1 (0.7) | 0 | 1 | |||||||
| Serotype in Silico | Number of Strains (%) | p-Value 1 Europe vs. North America | Countries (Number of Strains) | |
|---|---|---|---|---|
| Europe (n = 46) | North America (n = 143) | |||
| O2:H6 | 0 | 1 (0.7) | USA (1) | |
| O4:H31 | 11 (23.9) | 24 (16.8) | USA (23), Spain (7), UK (2), France (1), Sweden (1), Canada (1) | |
| O6:H31 | 1 (2.2) | 2 (1.4) | USA (2), UK (1) | |
| O15:H31 | 4 (8.7) | 26 (18.2) | USA (25), Spain (3), Canada (1), France (1) | |
| O18:H31 | 9 (19.6) | 10 (7.0) | 0.02203 | USA (10), UK (7), Spain (1), France (1) |
| O21:H14 | 1 (2.2) | 4 (2.8) | USA (4), Sweden (1) | |
| O21:H31 | 3 (6.5) | 2 (1.4) | Spain (2), UK (1), USA (1), Canada (1) | |
| O25:H31 | 4 (8.7) | 0 | 0.00317 | Spain (4) |
| O45:H31 | 1 (2.2) | 2 (1.4) | USA (2), Australia (2), Netherlands (1) | |
| O75:H31 | 0 | 5 (3.5) | USA (5) | |
| O83:H31 | 10 (21.7) | 57 (39.9) | 0.03291 | USA (52), Spain (4), Canada (2), Denmark (2), UK (1), France (1), Sweden (1), Germany (1) |
| O117:H28 | 1 (2.2) | 5 (3.5) | USA (5), Spain (1) | |
| O-unknown:H31 | 1 (2.2) | 4 (2.8) | USA (3), Canada (1), UK (1) | |
| O-unknown:H28 | 0 | 1 (0.7) | USA (1) | |
| Genome of Strain | Serotype of Strain | Origin | Cluster | MGEs | MOB/Inc Typing; Size (kb) | ||
|---|---|---|---|---|---|---|---|
| ICEs (kb of Contig) | Number of Plasmids | Plasmid_1 | Plasmid_2 | ||||
| LREC_341 | O18:H31 | Human | 2 | MOBQ (308) | 2 | MOBP1/nd 1; (164) | nd/p0111; (92) |
| LREC_342 | O18:H31 | Human | 2 | MOBQ (90) | 2 | MOBP1/nd; (720) | MOBQu/ColRNAI; (4) |
| LREC_343 | O4:H31 | Canine | 1 | MOBQ (474) | 1 | MOBF12/IncFII [F-:A-:B-], pCD1; (66) | |
| LREC_344 | O4:H31 | Canine | 1 | MOBQ (1208) | 0 | ||
| LREC_345 | O4:H31 | Canine | 1 | MOBQ (158) | 1 | MOBF12/IncFII [F2:A-:B-], pCD1; (75) | |
| LREC_346 | O4:H31 | Canine | 1 | MOBQ (653) | 1 | MOBP3/IncX1; (47) | |
| LREC_347 | O117:H28 | Canine | 1 | 1 | MOBP3/IncX1; (38) | ||
| LREC_348 | O4:H31 | Canine | 1 | MOBQ (472) | 0 | ||
| LREC_349 | O83:H31 | Canine | 1 | MOBQ (707) | 0 | ||
| LREC_350 | O83:H31 | Canine | 1 | MOBQ (707) | 0 | ||
| LREC_351 | O4:H31 | Canine | 1 | MOBQ (653) | 0 | ||
| LREC_352 | O25:H31 | Canine | 1 | MOBQ (658) | 0 | ||
| LREC_353 | O25:H31 | Canine | 1 | MOBQ (658) | 0 | ||
| LREC_354 | O25:H31 | Canine | 1 | MOBQ (658) | 0 | ||
| LREC_355 | O4:H31 | Canine | 1 | MOBQ (157) | 0 | ||
| LREC_356 | O83:H31 | Canine | 4 | MOBQ (2541) | 2 | MOBF12/IncFIB, IncFII [F-:A-:B52]; (162) | MOBP3/IncX1; (36) |
| LREC_357 | O83:H31 | Canine | 1 | MOBQ (1678) | 0 | ||
| LREC_358 | O25:H31 | Canine | 1 | MOBQ (660) | 0 | ||
| LREC_359 | O21:H31 | Canine | 1 | MOBQ (1317) | 0 | ||
| LREC_360 | O15:H31 | Canine | 1 | MOBQ (86) | 0 | ||
| LREC_361 | O21:H31 | Canine | 1 | MOBQ (1317) | 0 | ||
| LREC_362 | O15:H31 | Canine | 1 | MOBQ (641) | 0 | ||
| LREC_363 | O15:H31 | Canine | 1 | MOBQ (172) | 1 | MOBH11/IncHI2; (204) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flament-Simon, S.-C.; de Toro, M.; García, V.; Blanco, J.E.; Blanco, M.; Alonso, M.P.; Goicoa, A.; Díaz-González, J.; Nicolas-Chanoine, M.-H.; Blanco, J. Molecular Characteristics of Extraintestinal Pathogenic E. coli (ExPEC), Uropathogenic E. coli (UPEC), and Multidrug Resistant E. coli Isolated from Healthy Dogs in Spain. Whole Genome Sequencing of Canine ST372 Isolates and Comparison with Human Isolates Causing Extraintestinal Infections. Microorganisms 2020, 8, 1712. https://doi.org/10.3390/microorganisms8111712
Flament-Simon S-C, de Toro M, García V, Blanco JE, Blanco M, Alonso MP, Goicoa A, Díaz-González J, Nicolas-Chanoine M-H, Blanco J. Molecular Characteristics of Extraintestinal Pathogenic E. coli (ExPEC), Uropathogenic E. coli (UPEC), and Multidrug Resistant E. coli Isolated from Healthy Dogs in Spain. Whole Genome Sequencing of Canine ST372 Isolates and Comparison with Human Isolates Causing Extraintestinal Infections. Microorganisms. 2020; 8(11):1712. https://doi.org/10.3390/microorganisms8111712
Chicago/Turabian StyleFlament-Simon, Saskia-Camille, María de Toro, Vanesa García, Jesús E. Blanco, Miguel Blanco, María Pilar Alonso, Ana Goicoa, Juan Díaz-González, Marie-Hélène Nicolas-Chanoine, and Jorge Blanco. 2020. "Molecular Characteristics of Extraintestinal Pathogenic E. coli (ExPEC), Uropathogenic E. coli (UPEC), and Multidrug Resistant E. coli Isolated from Healthy Dogs in Spain. Whole Genome Sequencing of Canine ST372 Isolates and Comparison with Human Isolates Causing Extraintestinal Infections" Microorganisms 8, no. 11: 1712. https://doi.org/10.3390/microorganisms8111712
APA StyleFlament-Simon, S. -C., de Toro, M., García, V., Blanco, J. E., Blanco, M., Alonso, M. P., Goicoa, A., Díaz-González, J., Nicolas-Chanoine, M. -H., & Blanco, J. (2020). Molecular Characteristics of Extraintestinal Pathogenic E. coli (ExPEC), Uropathogenic E. coli (UPEC), and Multidrug Resistant E. coli Isolated from Healthy Dogs in Spain. Whole Genome Sequencing of Canine ST372 Isolates and Comparison with Human Isolates Causing Extraintestinal Infections. Microorganisms, 8(11), 1712. https://doi.org/10.3390/microorganisms8111712