Apophysomyces jiangsuensis sp. nov., a Salt Tolerant and Phosphate-Solubilizing Fungus from the Tidelands of Jiangsu Province of China



Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strain
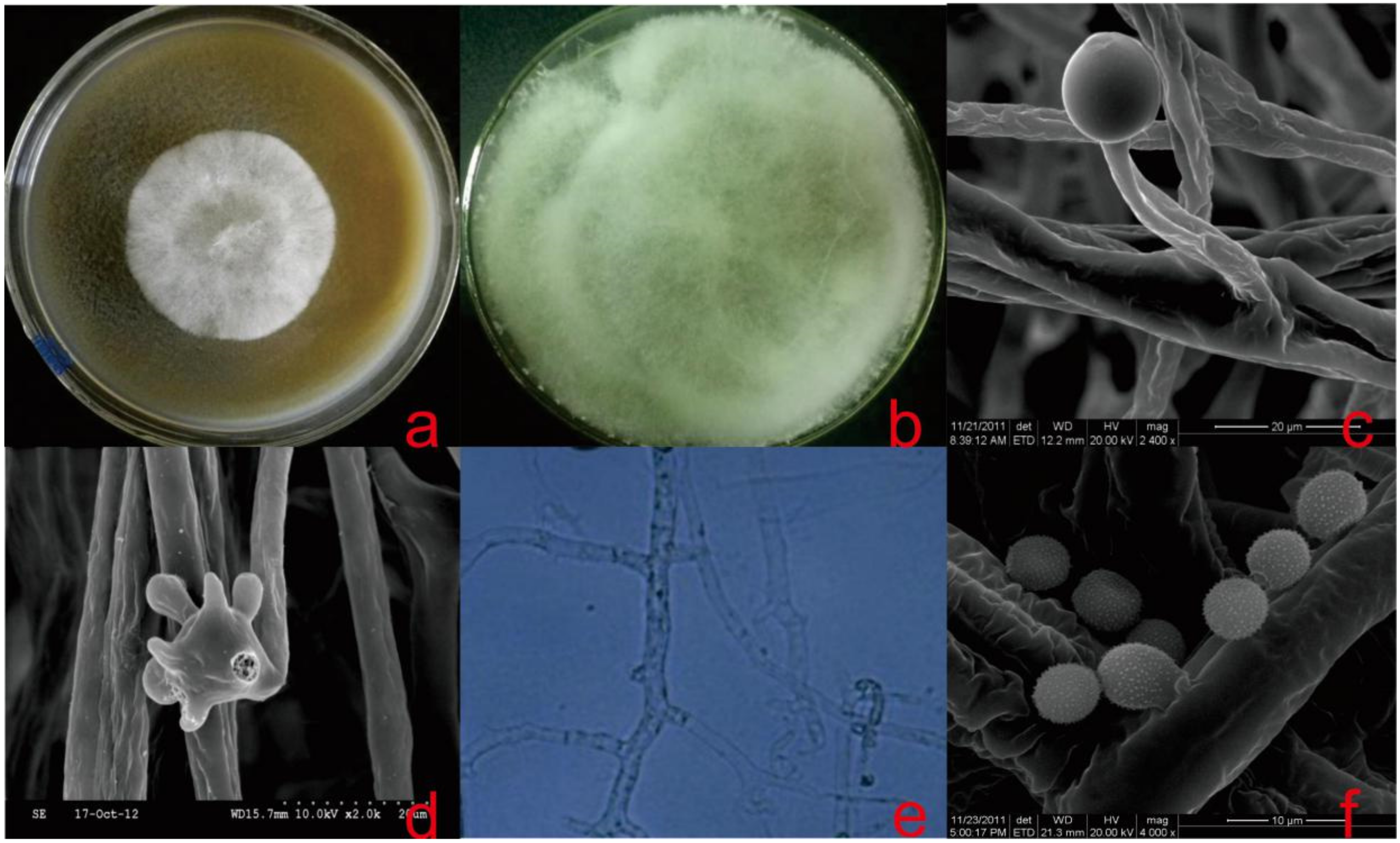
2.2. Morphological Studies and Growth Observation
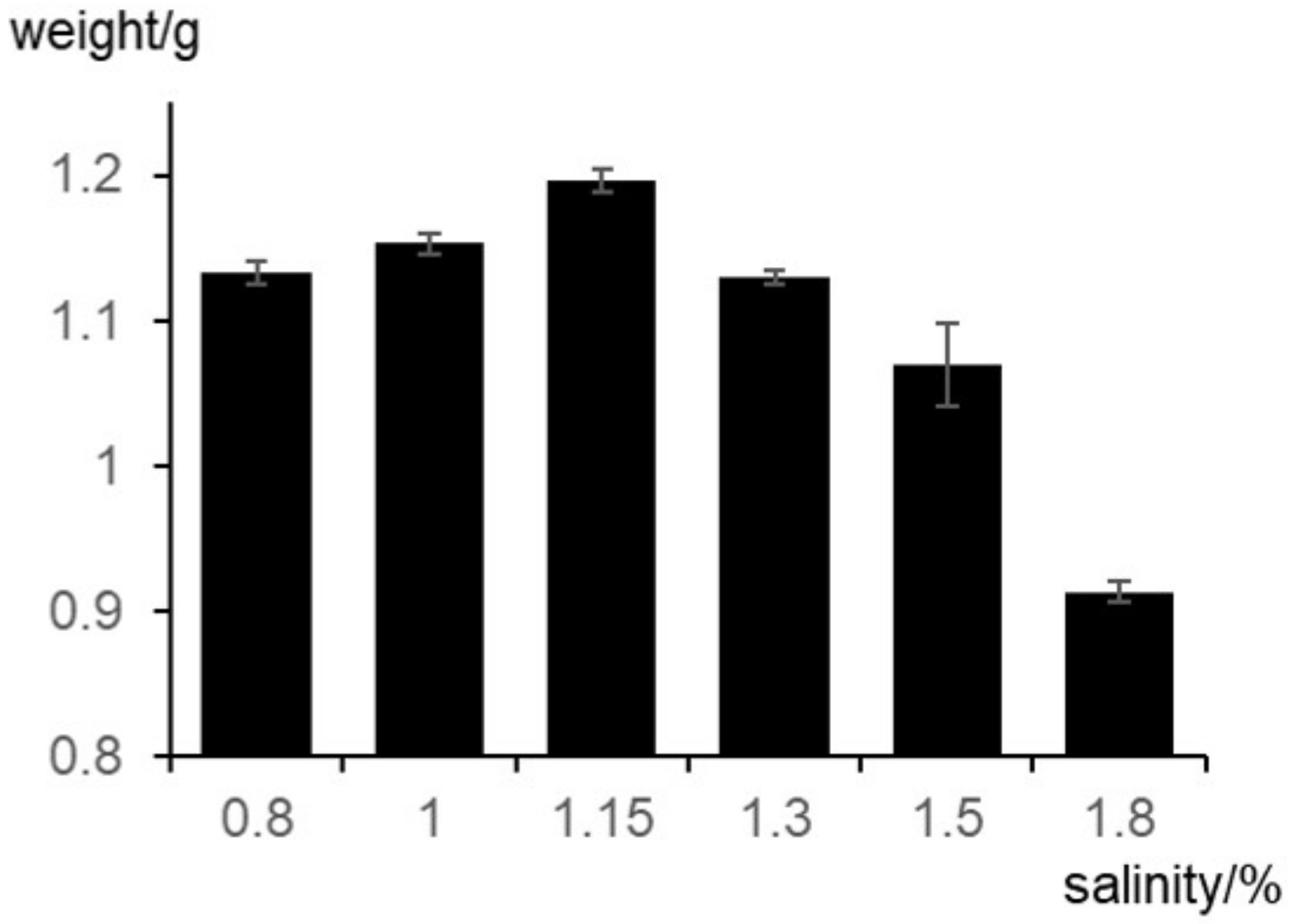
2.3. Optimum Growth Temperature and Salt Concentration
2.4. The Specific Method of Inducing Sporulation
2.5. DNA Extraction, Amplification, and Sequencing
2.6. Phylogenetic Analysis
2.7. Dissolved Phosphorus Ability
2.8. Low Molecular Weight (LMW) Organic Acids Derived from the Culture Liquor
3. Results
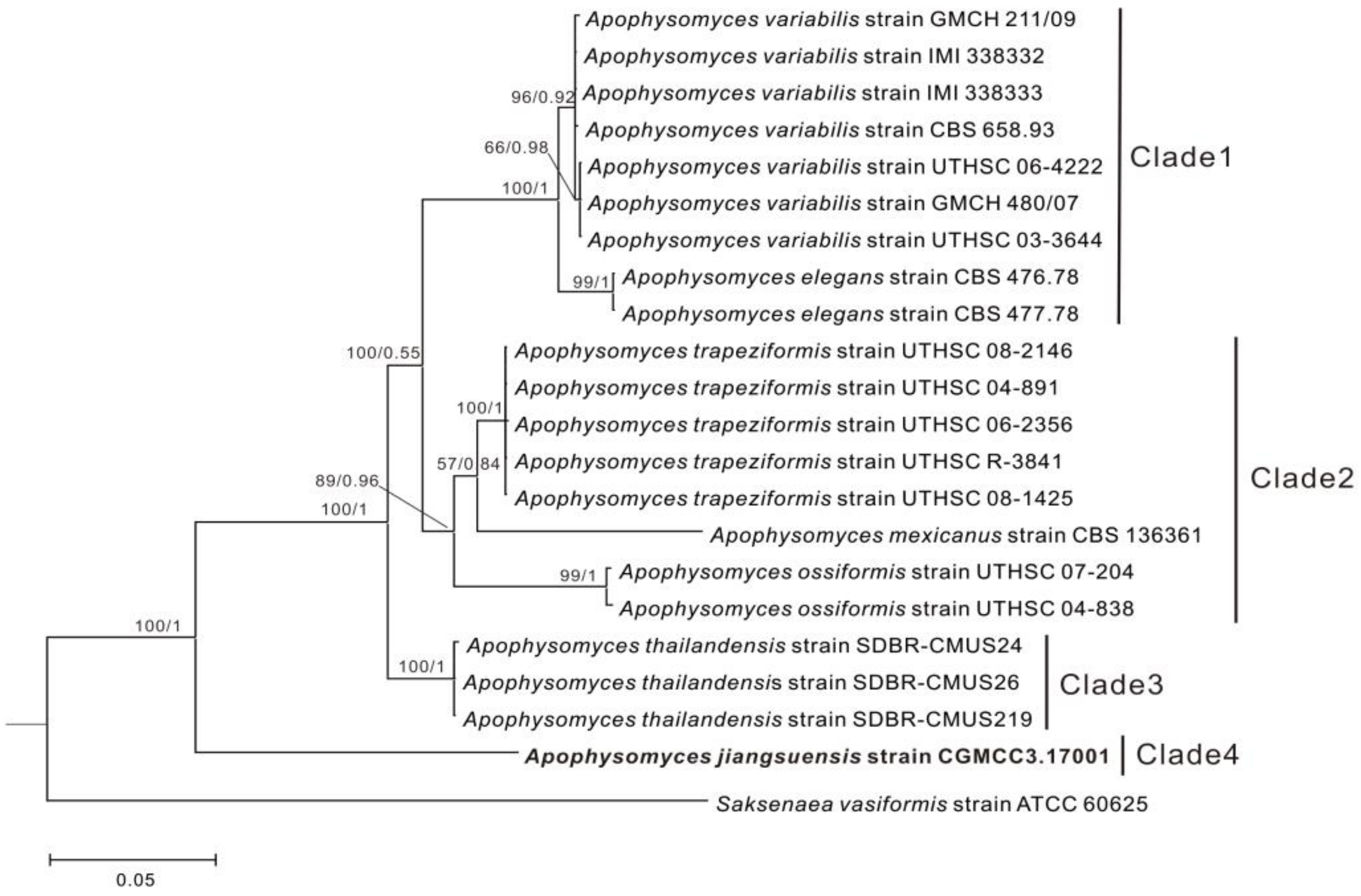
3.1. Phylogenetic Analyses
3.2. Taxonomy
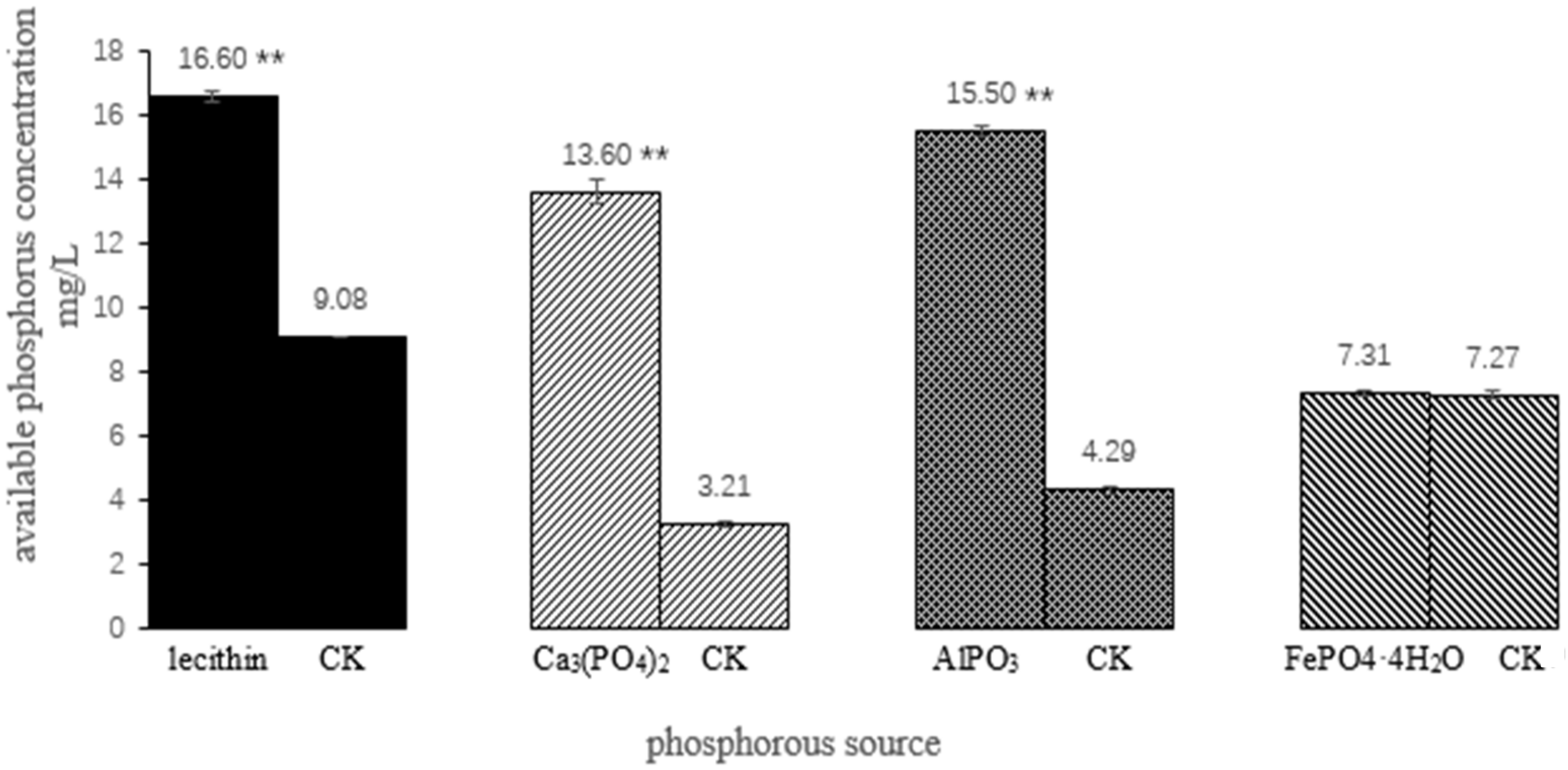
3.3. The Phosphate-Solubilizing Ability
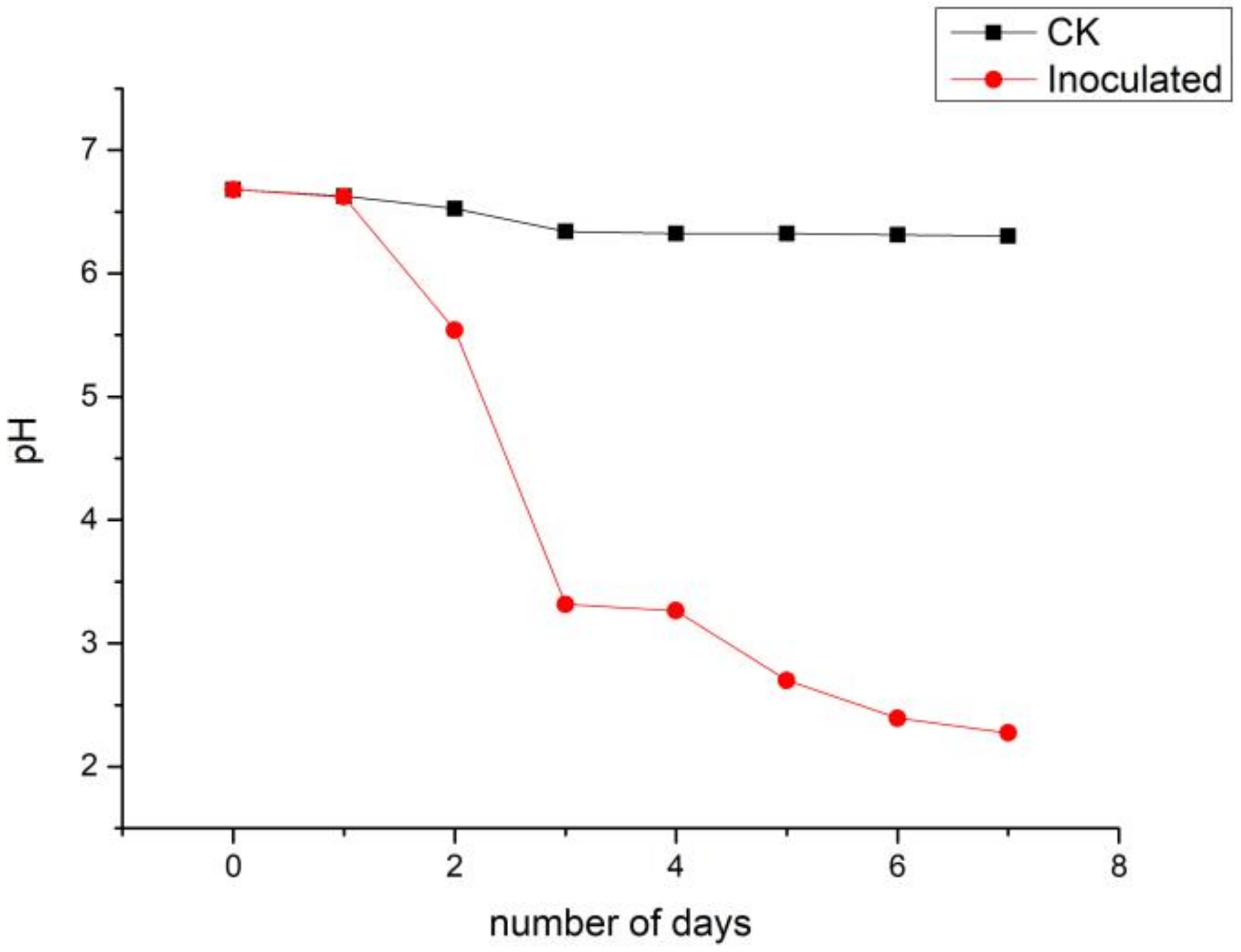
3.4. pH Dynamics of the Culture Liquor
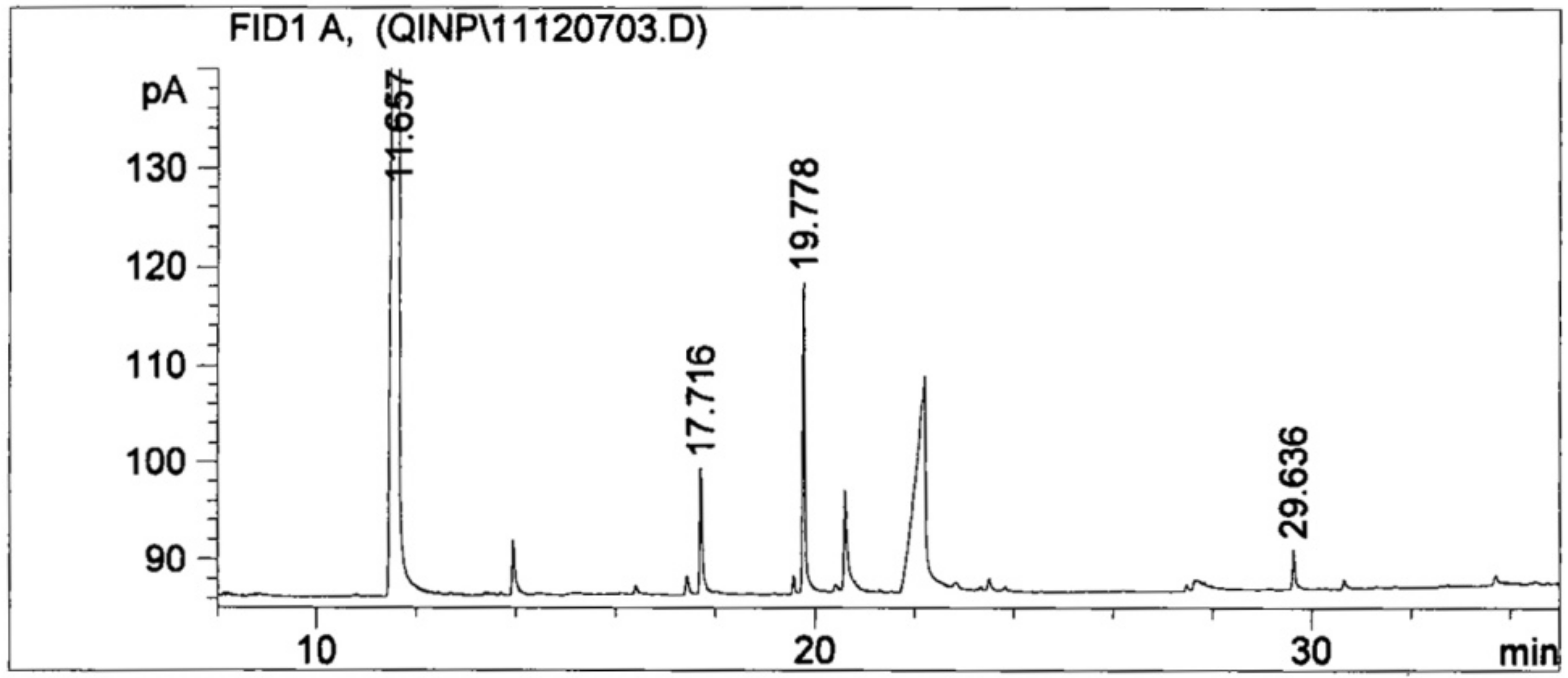
3.5. LMW Organic Acids Derived from the Culture Liquor
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Misra, P.C.; Srivastava, K.J.; Lata, K. Apophysomyces, a new genus of the Mucorales. Mycotaxon 1979, 8, 377–382. [Google Scholar]
- Benny, G.L.; Humber, R.A.; Morton, J.B. Zygomycota: Zygomycetes. In The Mycota; McLaughlin, D.J., McLaughlin, E.G., Lemke, P.A., Eds.; Systematics and Evolution: 113–146; Springer: Berlin, Germany, 2007; Volume VIIA. [Google Scholar]
- Hoffmann, K.; Pawłowska, J.; Walther, G.; Wrzosek, M.; de Hoog, G.S.; Benny, G.L.; Kirk, P.M.; Voigt, K. The family structure of the Mucorales: A synoptic revision based on comprehensive multigene-genealogies. Persoonia 2013, 30, 57–76. [Google Scholar] [CrossRef] [Green Version]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Cooter, R.D.; Lim, I.S.; Ellis, D.H.; Leitch, I.O.W. Burn wound zygomycosis caused by Apophysomyces elegans. J. Clin. Microbiol. 1990, 28, 2151–2153. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, A.; Ghosh, A.; Prasad, G.S.; David, J.K.; Gupta, S. Apophysomyces elegans: An emerging zygomycete in India. J. Clin. Microbiol. 2003, 41, 783–788. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M.; Smith, M.B.; McGinnis, M.R. Zygomycosis due to Apophysomyces elegans: Report of 2 cases and review of the literature. Arch. Pathol. Lab. Med. 1999, 123, 386–390. [Google Scholar]
- Reddy, I.S.; Rao, N.R.; Reddy, V.M.; Rao, R. Primary cutaneous mucormycosis (Zygomycosis) caused by Apophysomyces elegans. Indian J. Dermatol. Venereol. Leprol. 2008, 74, 367–370. [Google Scholar] [CrossRef]
- Alvarez, E.; Stchigel, A.M.; Cano, J.; Sutton, D.A.; Fothergill, A.W.; Chander, J.; Salas, V.; Rinaldi, M.G.; Guarro, J. Molecular phylogenetic diversity of the emerging mucoralean fungus Apophysomyces: Proposal of three new species. Rev. Iberoam. Micol. 2010, 27, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Bonifaz, A.; Stchigel, A.M.; Guarro, J.; Guevara, E.; Pintos, L.; Sanchis, M.; Cano-Lira, J.F. Primary cutaneous mucormycosis produced by the new species Apophysomyces mexicanus. J. Clin. Microbiol. 2014, 52, 4428–4431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khuna, S.; Suwannarach, N.; Kumla, J.; Jomkhwan, M.; Nuangmek, W.; Kiatsiriroat, T. Apophysomyces thailandensis (Mucorales, Mucoromycota), a new species isolated from soil in northern Thailand and its solubilization of non-soluble minerals. MycoKeys 2019, 45, 75–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumas, M.; Frossard, E.; Scholz, R.W. Modeling biogeochemical processes of phosphorus for global food supply. Chemosphere 2011, 84, 798–805. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Kiran, S.; Gulati, A.; Singh, B.; Tewari, R. Isolation and identification of phosphate solubilizing bacteria able to enhance the growth and aloin-A biosynthesis of Aloe barbadensis Miller. Microbiol. Res. 2012, 167, 358–363. [Google Scholar] [CrossRef]
- Mehta, P.; Walia, A.; Chauhan, A.; Shirkot, C.K. Plant growth promoting traits of phosphate-solubilizing rhizobacteria isolated from apple trees in trans Himalayan region of Himachal Pradesh. Arch. Microbiol. 2013, 195, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Sindhu, S.S.; Phour, M.; Choudhary, S.R.; Chaudhary, D. Phosphorus cycling: Prospects of using rhizosphere microorganisms for improving phosphorus nutrition of plants. Geomicrobiol. Biogeochem. 2014, 39, 199–237. [Google Scholar]
- Dorcas, Z.S.; Julio, C.R.L.; Jeffrey, J.C.; Ayixon, S.R.; Susana, V.D.; Mario, S.; Luz, E.D.B.; Jorge, L.F.M. Soil type affects organic acid production and phosphorus solubilization efficiency mediated by several native fungal strains from Mexico. Microorganisms 2020, 8, 1337. [Google Scholar] [CrossRef]
- Zhang, H.S.; Wu, X.H.; Li, G.; Qin, P. Interactions between arbuscular mycorrhizal fungi and phosphate-solubilizing fungus (Mortierella sp.) and their effects on Kostelelzkya virginica growth and enzyme activities of rhizosphere and bulk soils at different salinities. Biol. Fertil. Soils 2011, 47, 543–554. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.S.; Qin, F.F.; Qin, P.; Pan, S.M. Evidence that arbuscular mycorrhizal and phosphate-solubilizing fungi alleviate NaCl stress in the halophyte Kosteletzkya virginica: Nutrient uptake and ion distribution within root tissues. Mycorrhiza 2014, 24, 383–395. [Google Scholar] [CrossRef]
- Ellis, J.J.; Ajello, L. An unusual source of Apophysomyces elegans and a method for stimulating sporulation of Saksenaea vasiformis. Mycologia 1982, 74, 144–145. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Nicholas, K.; Nicholas, H. GeneDoc: A tool for editing and annotating multiple sequence alignments. Distributed by the authors. Embnew News 1997, 4, 14. [Google Scholar]
- Ronquist, F.; Teslenko, M.; Mark, P.V.D.; Ayres, D.L. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ–TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, D.T.; Chernomor, O.; Von, H.A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2017, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boltz, D.F. Book reviews: Colorimetric determination of nonmetals. Science 1958, 128, 709–710. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Kristiansson, E.; Ryberg, M.; Hallenberg, N.; Larsson, K.H. Intraspecific ITS variability in the kingdom Fungi as expressed in the international sequence databases and its implications for molecular species identification. Evol. Bioinform. 2008, 4, 193–201. [Google Scholar] [CrossRef]
- Padhye, A.A.; Ajello, L. Simple method of inducing sporulation by Apophysomyces elegans and Saksenaea vasiformis. J. Clin. Microbiol. 1988, 26, 1862–1863. [Google Scholar] [CrossRef] [Green Version]
- Sundareshwar, P.V.; Morris, J.T.; Koepfler, E.K. Phosphorus Limitation of Coastal Ecosystem Processes. Science 2003, 229, 563–565. [Google Scholar] [CrossRef] [Green Version]
- Mikanova, O.; Novakova, J. Evaluation of the P-solubilizing activity of soil microorganisms and its sensitivity to soluble phosphate. Rostl. Vyrob. 2002, 48, 397–400. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Isolate | Source | GenBank Accession No. | ||
|---|---|---|---|---|---|
| ITS | 28S | Reference | |||
| A. variabilis | UTHSC 06-4222 | Dolphin, Bahamas | FN556428 | FN554255 | [9] |
| A. variabilis | UTHSC 03-3644 | Dolphin, FL, USA | FN556431 | FN554259 | [9] |
| A. variabilis | IMI 338333 | Daly river, Australia | FN556439 | FN554256 | [9] |
| A. variabilis | IMI 338332 | Ankle aspirate, Australia | FN556438 | FN554257 | [9] |
| A. variabilis | GMCH 480/07 | Cutaneous infection, India | FN556442 | FN554253 | [9] |
| A. variabilis | GMCH 211/09 | Cutaneous infection, India | FN556443 | FN554254 | [9] |
| A. variabilis | CBS 658.93 | Osteomyelitis, Netherlands Antilles | FN556436 | FN554258 | [9] |
| A. trapeziformis | UTHSC R-3841 | Necrotic face tissue, GA, USA | FN556434 | FN554263 | [9] |
| A. trapeziformis | UTHSC 08-2146 | Skin biopsy, CO, USA | FN556430 | FN554260 | [9] |
| A. trapeziformis | UTHSC 08-1425 | Abdominal tissue, PHA, USA | FN556429 | FN554261 | [9] |
| A. trapeziformis | UTHSC 06-2356 | Dolphin, TX, USA | FN556427 | FN554262 | [9] |
| A. trapeziformis | UTHSC 04-891 | Sinus, MN, USA | FN556433 | FN554264 | [9] |
| A. thailandensis | SDBR-CMUS26 | Soil, Thailand | MH733251 | MH733254 | [11] |
| A. thailandensis | SDBR-CMUS24 | Soil, Thailand | MH733250 | MH733253 | [11] |
| A. thailandensis | SDBR-CMUS219 | Soil, Thailand | MH733252 | MH733255 | [11] |
| A. ossiformis | UTHSC 07-204 | Facial cellulitis, AZ, USA | FN556435 | FN554251 | [9] |
| A. ossiformis | UTHSC 04-838 | Cellulitis wound leg, MN, USA | FN556432 | FN554252 | [9] |
| A. mexicanus | CBS 136361 | Human, Mexican | HG974255 | HG974256 | [10] |
| A. elegans | CBS 477.78 | Soil, Gorakhpur, India | FN556437 | FN554250 | [9] |
| A. elegans | CBS 476.78 | Soil, Deoria, India | FN556440 | FN554249 | [9] |
| A. jiangsuensis | CGMCC3.17001 | Soil, JiangSu, China | KF048102 | KF055451 | This study |
| Medium | Isolate Diameter (mm/day) | Isolate Color | Height (mm) | Odor | ||||
|---|---|---|---|---|---|---|---|---|
| 1 | 3 | 5 | 7 | Front | Reverse | |||
| MAM | 15.4 ± 0.07 | 80.6 ± 0.07 | 90 ± 0.00 | 90 ± 0.00 | white | brown | 77.3 ± 0.07 | / |
| PDA | 12.1 ± 0.03 | 70.4 ± 0.05 | 90 ± 0.00 | 90 ± 0.00 | white | brown | 70.1 ± 0.53 | / |
| SDA | 1.7 ± 0.00 | 35.3 ± 0.07 | 72.8 ± 0.09 | 90 ± 0.00 | white | white | 35.3 ± 0.10 | / |
| MEA | 5.3 ± 0.05 | 50.7 ± 0.12 | 84.4 ± 0.09 | 90 ± 0.00 | white turn yellow | brown | 120.4 ± 0.12 | mushroom aroma |
| OMA | 2.1 ± 0.05 | 20.4 ± 0.07 | 31.2 ± 0.07 | 33.3 ± 0.05 | white turn yellow | brown | 20 ± 0.47 | / |
| CZA | – | – | – | – | – | – | – | / |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Han, R.; Zhang, H.; Song, Y.; Zhao, F.; Qin, P. Apophysomyces jiangsuensis sp. nov., a Salt Tolerant and Phosphate-Solubilizing Fungus from the Tidelands of Jiangsu Province of China. Microorganisms 2020, 8, 1868. https://doi.org/10.3390/microorganisms8121868
Li S, Han R, Zhang H, Song Y, Zhao F, Qin P. Apophysomyces jiangsuensis sp. nov., a Salt Tolerant and Phosphate-Solubilizing Fungus from the Tidelands of Jiangsu Province of China. Microorganisms. 2020; 8(12):1868. https://doi.org/10.3390/microorganisms8121868
Chicago/Turabian StyleLi, Siyu, Ruiming Han, Huanshi Zhang, Yongchun Song, Fugeng Zhao, and Pei Qin. 2020. "Apophysomyces jiangsuensis sp. nov., a Salt Tolerant and Phosphate-Solubilizing Fungus from the Tidelands of Jiangsu Province of China" Microorganisms 8, no. 12: 1868. https://doi.org/10.3390/microorganisms8121868
APA StyleLi, S., Han, R., Zhang, H., Song, Y., Zhao, F., & Qin, P. (2020). Apophysomyces jiangsuensis sp. nov., a Salt Tolerant and Phosphate-Solubilizing Fungus from the Tidelands of Jiangsu Province of China. Microorganisms, 8(12), 1868. https://doi.org/10.3390/microorganisms8121868