Importance of the Farm Environment and Wildlife for Transmission of Campylobacter jejuni in A Pasture-Based Dairy Herd



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Sampling Collection
2.3. Cultivation and Identification of C. jejuni
2.4. Genotyping of C. jejuni Single-Colony Isolates by Enterobacterial Repetitive Intergenic Consensus (ERIC)-PCR and Multi Locus Sequence Typing (MLST)
2.5. Statistical Analysis
3. Results
3.1. Prevalence of C. jejuni in the Cows and the Farm Environment
3.2. Characterization of C. jejuni Isolates
3.2.1. Ruminant Isolates
3.2.2. Wildlife Isolates
3.2.3. Farm Environment Isolates
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Blaser, M.J.; Engberg, J. Clinical Aspects of Campylobacter Jejuni and Campylobacter Coli Infections, Campylobacter, 3rd ed.; Nachamkin, I., Szymanski, C.M., Blaser, M.J., Eds.; ASM Press: Washington, DC, USA, 2008; pp. 99–121. [Google Scholar]
- CDC. Foodborne Diseases Active Surveillance Network (FoodNet): FoodNet 2015 Surveillance Report (Final Data); Department of Health and Human Services, CDC: Atlanta, GA, USA, 2017; p. 25. [Google Scholar]
- EFSA (European Food Safety Authority) and ECDC (European Centre for Disease Prevention and Control). The European Union One Health 2018 zoonoses report. EFSA J. 2019, 17, 5926. [Google Scholar]
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Mitchell, H.M.; Man, S.M. Global epidemiology of Campylobacter infection. Clin. Microbiol. Rev. 2015, 28, 681–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, D.J.; Gabriel, E.; Leatherbarrow, A.J.H.; Cheesbrough, J.; Gee, S.; Bolton, E.; Fox, A.; Fearnhead, P.; Hart, C.A.; Diggle, P.J. Tracing the source of campylobacteriosis. PLoS Genet. 2008, 4, e1000203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lake, R.; Ashmore, E.; Cressey, P.; Horn, B.; Pirikahu, S.; Sherwood, J. Source Assigned Campylobacteriosis in New Zealand Study; MPI Report: Wellington, New Zealand, 2020. [Google Scholar]
- Taylor, E.V.; Herman, K.M.; Ailes, E.C.; Fitzgerald, C.; Yoder, J.S.; Mahon, B.E.; Tauxe, R.V. Common source outbreaks of Campylobacter infection in the USA, 1997–2008. Epidemiol. Infect. 2013, 141, 987–996. [Google Scholar] [CrossRef] [Green Version]
- Kwan, P.S.; Xavier, C.; Santovenia, M.; Pruckler, J.; Stroika, S.; Joyce, K.; Gardner, T.; Fields, P.I.; McLaughlin, J.; Tauxe, R.V.; et al. Multilocus sequence typing confirms wild birds as the source of a Campylobacter outbreak associated with the consumption of raw peas. Appl. Environ. Microbiol. 2014, 80, 4540–4546. [Google Scholar] [CrossRef] [Green Version]
- Verhoeff-Bakkenes, L.; Jansen, H.A.P.M.; In’t Veld, P.H.; Beumer, R.R.; Zwietering, M.H.; van Leusden, F.M. Consumption of raw vegetables and fruits: A risk factor for campylobacter infections. Int. J. Food Microbiol. 2011, 144, 406–412. [Google Scholar] [CrossRef]
- Gilpin, B.J.; Scholes, P.; Robson, B.; Savill, M.G. The transmission of thermotolerant Campylobacter spp. to people living or working on dairy farms in New Zealand. Zoonoses Public Health 2008, 55, 352–360. [Google Scholar] [CrossRef]
- Gilpin, B.J.; Walker, T.; Paine, S.; Sherwood, J.; Mackereth, G.; Wood, T.; Hambling, T.; Hewison, C.; Brounts, A.; Wilson, M.; et al. A large scale waterborne campylobacteriosis outbreak, Havelock North, New Zealand. J. Infect. 2020, 81, 390–395. [Google Scholar] [CrossRef]
- Fitzenberger, J.; Uphoff, H.; Gawrich, S.; Hauri, A.M. Urban–rural differences of age- and species-specific campylobacteriosis incidence, Hesse, Germany, July 2005–June 2006. Eurosurveillance 2010, 15, 19693. [Google Scholar] [CrossRef]
- Ceuppens, S.; Johannessen, G.S.; Allende, A.; Tondo, E.C.; El-Tahan, F.; Sampers, I.; Jacxsens, L.; Uyttendaele, M. Risk factors for Salmonella, Shiga Toxin-Producing Escherichia coli and Campylobacter occurrence in primary production of leafy greens and strawberries. Int. J. Environ. Res. Public Health 2015, 12, 9809–9831. [Google Scholar] [CrossRef] [Green Version]
- Sanderson, R.A.; Maas, J.A.; Blain, A.P.; Gorton, R.; Ward, J.; O’Brien, S.J.; Hunter, P.R.; Rushton, S.P. Spatio-temporal models to determine association between Campylobacter cases and environment. Int. J. Epidemiol. 2018, 47, 202–216. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization, Food and Agriculture Organization of the United Nations & World Organisation for Animal Health. The Global View of Campylobacteriosis: Report of an Expert Consultation, Utrecht, Netherlands, 9–11 July 2012; World Health Organization: Geneva, Switzerland, 2013; 57p, Available online: https://apps.who.int/iris/handle/10665/80751 (accessed on 2 May 2013).
- Wagenaar, J.A.; French, N.P.; Havelaar, A.H. Preventing Campylobacter at the source: Why is it so difficult? Clin. Infect. Dis. 2013, 57, 1600–1606. [Google Scholar] [CrossRef] [Green Version]
- Hänninen, M.L.; Niskanen, M.; Korhonen, L. Water as reservoir for Campylobacter jejuni infection in cows studied by serotyping and pulsed-field gel electrophoresis (PFGE). J. Vet. Med. B 1998, 45, 37–42. [Google Scholar]
- LeJeune, J.T.; Besser, T.E.; Merrill, N.L.; Rice, D.H.; Hancock, D.D. Livestock Drinking Water Microbiology and the Factors Influencing the Quality of Drinking Water Offered to Cattle. J. Dairy Sci. 2001, 84, 1856–1862. [Google Scholar] [CrossRef]
- Jaakkonen, A.; Castro, H.; Hallanvuo, S.; Ranta, J.; Rossi, M.; Isidro, J.; Lindström, M.; Hakkinen, M. Longitudinal study of Shiga toxin-producing Escherichia coli and Campylobacter jejuni on Finnish dairy farms and in raw milk. Appl. Environ. Microbiol. 2019, 85, e02910-18. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, B.; Connolly, J.H.; Madie, P.; Davies, P.R. Prevalence and clonal diversity of Campylobacter jejuni from dairy farms and urban sources. N. Z. Vet. J. 2004, 52, 378–383. [Google Scholar] [CrossRef]
- Devane, M.L.; Nicol, C.; Ball, A.; Klena, J.D.; Scholes, P.; Hudson, J.A.; Baker, M.G.; Gilpin, B.J.; Garrett, N.; Savill, M.G. The occurrence of Campylobacter subtypes in environmental reservoirs and potential transmission routes. J. Appl. Microbiol. 2005, 98, 980–990. [Google Scholar] [CrossRef]
- Donnison, A. Isolation of Thermotolerant Campylobacter—Review and Methods for New Zealand Laboratories; Ministry of Health: Wellington, New Zealand, 2003; p. 79. [Google Scholar]
- Vandamme, P.; Van Doorn, L.J.; Al Rashid, S.T.; Quint, W.G.V.; Van Der Plas, J.; Chan, V.L.; On, S.L.W. Campylobacter hyoilei Alderton et al. 1995 and Campylobacter coli Véron and Chatelain 1973 are subjective synonyms. Int. J. Syst. Bacteriol. 1997, 47, 1055–1060. [Google Scholar] [CrossRef] [Green Version]
- Rapp, D.; Ross, C.M.; Cave, V.; Muirhead, R.W. Prevalence, concentration and genotypes of Campylobacter jejuni in feces from dairy herds managed in farm systems with or without housing. J. Appl. Microbiol. 2014, 116, 1035–1043. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package; R Package Version 2.5-6; 2019; Available online: https://CRAN.R-project.org/package=vegan (accessed on 29 October 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 29 October 2020).
- Kwan, P.L.S.; Birtles, A.; Bolton, F.J.; French, N.P.; Robinson, S.E.; Newbold, L.S.; Upton, M.; Fox, A.J. Longitudinal study of the molecular epidemiology of Campylobacter jejuni in cattle on dairy farms. Appl. Environ. Microbiol. 2008, 74, 3626–3633. [Google Scholar] [CrossRef] [Green Version]
- Robinson, D.A. Campylobacter Infection in Milking Herds, Campylobacter: Epidemiology, Pathogenesis, and Biochemistry; Newell, D.G., Ed.; MTP Press: Lancaster, UK, 1982; p. 274. [Google Scholar]
- Ross, C.M.; Donnison, A.M.; Clark, D.A. Effect of using a stand-off pad on Campylobacter jejuni strain diversity in a herd of dairy cows. Lett. Appl. Microbiol. 2008, 47, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, E.M. Occurrence and strain diversity of thermophilic campylobacters in cattle of different age groups in dairy herds. Lett. Appl. Microbiol. 2002, 35, 85–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheppard, S.K.; Cheng, L.; Méric, G.; de Haan, C.P.A.; Llarena, A.-K.; Marttinen, P.; Vidal, A.; Ridley, A.; Clifton-Hadley, F.; Connor, T.R.; et al. Cryptic ecology among host generalist Campylobacter jejuni in domestic animals. Mol. Ecol. 2014, 23, 2442–2451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullner, P.; Shabolt, T.; Collins-Emerson, J.M.; Midwinter, A.C.; Spencer, S.E.F.; Marshall, J.; Carter, P.E.; Campbell, D.M.; Wilson, D.J.; Hathaway, S.; et al. Molecular and spatial epidemiology of human campylobacteriosis: Source association and genotype-related risk factors. Epidemiol. Infect. 2010, 138, 1372–1383. [Google Scholar] [CrossRef] [Green Version]
- Besser, T.E.; Lejeune, J.T.; Rice, D.H.; Berg, J.; Stilborn, R.P.; Kaya, K.; Bae, W.; Hancock, D.D. Increasing prevalence of Campylobacter jejuni in feedlot cattle through the feeding period. Appl. Environ. Microbiol. 2005, 71, 5752–5758. [Google Scholar] [CrossRef] [Green Version]
- Minihan, D.; Whyte, P.; O’Mahony, M.; Fanning, S.; McGill, K.; Collins, J.D. Campylobacter spp. in Irish feedlot cattle: A longitudinal study involving pre-harvest and harvest phases of the food chain. J. Vet. Med. B Infect. Dis Vet. Public Health 2004, 51, 28–33. [Google Scholar] [CrossRef]
- Jones, K. Campylobacters in water, sewage and the environment. J. Appl. Microbiol. 2001, 90, 68S–70S. [Google Scholar] [CrossRef]
- Fremaux, B.; Prigent-Combaret, C.; Beutin, L.; Gleizal, A.; Trevisan, D.; Quetin, P.; Jocteur-Monrozier, L.; Rozand, C. Survival and spread of Shiga toxin-producing Escherichia coli in alpine pasture grasslands. J. Appl. Microbiol. 2010, 108, 1332–1343. [Google Scholar] [CrossRef]
- Scheile, T.; Isselstein, J.; Tonn, B. Herbage biomass and uptake under low-input grazing as affected by cattle and sheep excrement patches. Nutr. Cycl. Agroecosyst. 2018, 112, 277–289. [Google Scholar] [CrossRef]
- Ahlstrom, C.; Muellner, P.; Lammers, G.; Jones, M.; Octavia, S.; Lan, R.; Heller, J. Shiga Toxin-Producing Escherichia coli O157 shedding dynamics in an Australian beef herd. Front. Vet. Sci. 2017, 27, 200. [Google Scholar] [CrossRef] [Green Version]
- Jones, B.A.; Grace, D.; Kock, R.; Alonso, S.; Rushton, J.; Said, M.Y.; McKeever, D.; Mutua, F.; Young, J.; McDermott, J.; et al. Zoonosis emergence linked to agricultural intensification and environmental change. Proc. Nat. Acad. Sci. USA 2013, 110, 8399–8404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanad, Y.M.; Closs, G., Jr.; Kumar, A.; LeJeune, J.T.; Rajashekara, G. Molecular epidemiology and public health relevance of Campylobacter isolated from dairy cattle and European starlings in Ohio, USA. Foodborne Pathog. Dis. 2013, 10, 229–236. [Google Scholar] [CrossRef]
- Mohan, V.; Stevensen, M.; Marshall, J.; Fearnhead, P.; Holland, B.R.; Hotter, G.; French, N.P. Campylobacter jejuni colonization and population structure in urban populations of ducks and starlings in New Zealand. Microbiol. Open 2013, 2, 659–673. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Meinersmann, R.J.; Sahin, O.; Wu, Z.; Dai, L.; Carlson, J.; Plumblee Lawrence, J.; Genzlinger, L.; LeJeune, J.T.; Zhang, Q. Wide but variable distribution of a hypervirulent Campylobacter jejuni clone in beef and dairy cattle in the United States. Appl. Environ. Microbiol. 2017, 83, e01425-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hald, B.; Skov, M.N.; Nielsen, E.M.; Rahbek, C.; Madsen, J.J.; Wainø, M.; Chriél, M.; Nordentoft, S.; Baggesen, D.L.; Madsen, M. Campylobacter jejuni and Campylobacter coli in wild birds on Danish livestock farms. Acta Vet. Scand. 2016, 58, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, L.A.; Bennett, M.; Coffey, P.; Elliott, J.; Jones, T.R.; Jones, R.C.; Lahuerta-Marin, A.; Leatherbarrow, A.H.; McNiffe, K.; Norman, D.; et al. Molecular epidemiology and characterisation of Campylobacter spp. isolated from wild bird populations in northern England. Appl. Environ. Microbiol. 2009, 75, 3007–3015. [Google Scholar] [CrossRef] [Green Version]
- Colles, F.M.; McCarthy, N.D.; Howe, J.C.; Devereux, C.L.; Gosler, A.G.; Maiden, M.C.J. Dynamics of Campylobacter colonization of a natural host, Sturnus vulgaris (European Starling). Environ. Microbiol. 2009, 11, 258–267. [Google Scholar] [CrossRef] [Green Version]
- Smith, O.M.; Snyder, W.E.; Owen, J.P. Are we overestimating risk of enteric pathogen spillover from wild birds to humans? Biol. Rev. 2020, 95, 652–679. [Google Scholar] [CrossRef] [Green Version]
- Mourkas, E.; Taylor, A.J.A.; Méric, G.; Bayliss, S.C.S.; Pascoe, B.; Mageiros, L.; Calland, J.K.; Hitchings, M.D.; Ridley, A.; Vidal, A.; et al. Agricultural intensification and the evolution of host specialism in the enteric pathogen Campylobacter jejuni. Proc. Natl. Acad. Sci. USA 2020, 117, 1118–11028. [Google Scholar] [CrossRef]
- Rapp, D.; Ross, C.M.; Cave, V.M. Excretion patterns of Campylobacter jejuni by dairy cows. N. Z. J. Agric. Res. 2019, 62, 83–95. [Google Scholar] [CrossRef]
- De Haan, C.P.; Llarena, A.K.; Revez, J.; Hanninen, M.L. Association of Campylobacter jejuni metabolic traits with multilocus sequence types. Appl. Environ. Microbiol. 2012, 78, 5550–5554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gripp, E.; Hlahla, D.; Didelot, X.; Kops, F.; Maurischat, S.; Tedin, K.; Alter, T.; Ellerbroek, L.; Schreiber, K.; Schomburg, D.; et al. Closely related Campylobacter jejuni strains from different sources reveal a generalist rather than a specialist lifestyle. BMC Genom. 2011, 12, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]


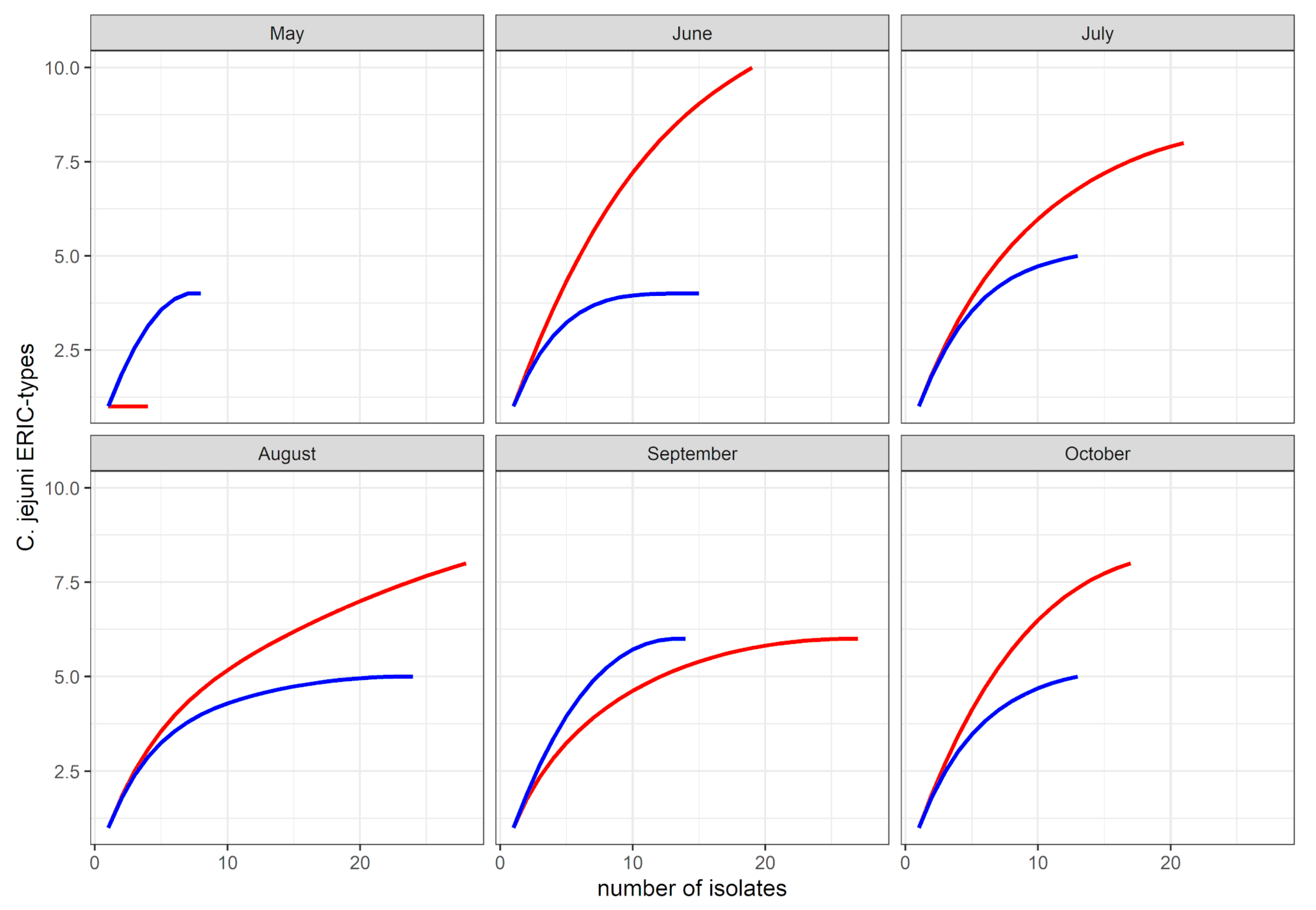{kind=link}
| Sample Type | Number C. jejuni-Positive Sample/Total Samples (%) | ||||||
|---|---|---|---|---|---|---|---|
| May | June | July | Aug | Sept | Oct | Total | |
| Cows faeces | 4/15 (27) | 8/15 (53) | 8/15 (53) | 12/15 (80) | 10/15 (67) | 7/15 (47) | 49/90 (54) |
| Pasture | 1/2 (50) | 1/3 (33) | 2/3 (67) | 3/4 (75) | 4/4 (100) | 2/3 (67) | 13/19 (68) |
| Soil | 0/2 (0) | 1/3 (33) | 1/3 (33) | 2/4 (50) | 1/4 (25) | 0/3 (0) | 5/19 (26) |
| Water | 0/3 (0) | 0/3 (0) | 1/4 (25) | 0/5 (0) | 1/5 (20) | 2/3 (67) | 4/23 (17) |
| Bird droppings | 2/12 (17) | 13/35 (37) | 12/41 (29) | 15/45 (33) | 25/42 (60) | 10/36 (28) | 77/211 (36) |
| Rat faeces | 0/0 (0) | 0/0 (0) | 0/0 (0) | 0/1 (0) | 6/6 (100) | 0/0 (0) | 6/7 (86) |
| Total | 154/369 (42) | ||||||
| ERIC Genotype | Total Number of Isolates | % Isolates within Datasets | MLST | |||
|---|---|---|---|---|---|---|
| Cow (n = 87) | Birds (n = 116) | Environment (Pasture, Soil, Water) (n = 40) | Sequence Type | Clonal Complex | ||
| 1 | 40 | 23 | 5 | 35 | ST-61 | CC-61 |
| 5 | 3 | 2 | 1 | 0 | ST-61 | |
| 6 | 35 | 20 | 9 | 20 | ST-42 | CC-42 |
| 7 | 4 | 4 | 0 | 0 | ST-42 | |
| 29 | 3 | 3 | 0 | 0 | ST-42 | |
| 4 | 1 | 0 | 1 | 0 | ST-42 | |
| 12 | 17 | 2 | 13 | 0 | ST-45 | CC-45 |
| 30 | 4 | 2 | 2 | 0 | ST-45 | |
| 10 | 6 | 0 | 5 | 0 | ST-45 | |
| 27 | 7 | 0 | 6 | 0 | ST-45 | |
| 25 | 3 | 0 | 3 | 0 | ST-45 | |
| 11 | 1 | 1 | 0 | 0 | ST-45 | |
| 20 | 1 | 0 | 0 | 3 | ST-45 | |
| 24 | 1 | 0 | 1 | 0 | ST-583 | |
| 13 | 16 | 10 | 4 | 5 | ST-38 | CC-38 |
| 21 | 5 | 0 | 1 | 10 | ST-50 | CC-21 |
| 3 | 5 | 2 | 2 | 3 | ST-50 | |
| 14 | 5 | 1 | 3 | 0 | ST-4337 | |
| 22 | 7 | 0 | 3 | 10 | ST-2345 | CC-206 |
| 19 | 2 | 2 | 0 | 0 | ST-2345 | |
| 9 | 4 | 0 | 3 | 0 | ST-2026 | CC-403 |
| 16 | 3 | 0 | 3 | 0 | ST-2343 | CC-48 |
| 26 | 2 | 0 | 2 | 0 | ST-508 | CC-508 |
| 28 | 2 | 0 | 2 | 0 | ST-508 | |
| 15 | 1 | 0 | 1 | 0 | ST-682 | CC-682 |
| 17 | 22 | 15 | 8 | 0 | ST-10821 | |
| 23 | 6 | 0 | 5 | 0 | Not identified | |
| 8 | 1 | 0 | 1 | 0 | Not identified | |
| 2 | 34 | 10 | 17 | 13 | Not identified | |
| 18 | 2 | 0 | 1 | 3 | Not identified | |
| Sample Type | C. jejuni ERIC Genotypes | |||||
|---|---|---|---|---|---|---|
| May | June | July | Aug | Sept | Oct | |
| Cows faeces | 1, 6, 12, 17 | 1, 6, 13, 17 | 7, 11, 13, 17, 29 | 1, 2, 3, 5, 6 | 1, 2, 6, 13, 17, 30 | 1, 6, 14, 17, 19 |
| Pasture | 1, 13 | 22 | 2, 6, 20 | 2, 3, 6, | 1, 2, 13 | 1 |
| Soil | 22 | 6 | 1, 6 | |||
| Water | 1 | 18 | 21 | |||
| Birds droppings | 10 | 4, 6, 8, 9, 12, 18, 22, 25, 26, 27 | 1, 2, 5, 6, 12, 14, 17, 23 | 2, 3, 6, 12, 14, 17, 24, 27 | 1, 2, 6, 13, 27, 30 | 9, 10, 12, 15, 16, 21, 27, 28 |
| Rat faeces | 12, 30 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rapp, D.; Ross, C.; Hea, S.-Y.; Brightwell, G. Importance of the Farm Environment and Wildlife for Transmission of Campylobacter jejuni in A Pasture-Based Dairy Herd. Microorganisms 2020, 8, 1877. https://doi.org/10.3390/microorganisms8121877
Rapp D, Ross C, Hea S-Y, Brightwell G. Importance of the Farm Environment and Wildlife for Transmission of Campylobacter jejuni in A Pasture-Based Dairy Herd. Microorganisms. 2020; 8(12):1877. https://doi.org/10.3390/microorganisms8121877
Chicago/Turabian StyleRapp, Delphine, Colleen Ross, Shen-Yan Hea, and Gale Brightwell. 2020. "Importance of the Farm Environment and Wildlife for Transmission of Campylobacter jejuni in A Pasture-Based Dairy Herd" Microorganisms 8, no. 12: 1877. https://doi.org/10.3390/microorganisms8121877
APA StyleRapp, D., Ross, C., Hea, S. -Y., & Brightwell, G. (2020). Importance of the Farm Environment and Wildlife for Transmission of Campylobacter jejuni in A Pasture-Based Dairy Herd. Microorganisms, 8(12), 1877. https://doi.org/10.3390/microorganisms8121877