Maternal Microbiome and Infections in Pregnancy


 , ,
, , 
Abstract
:1. Introduction
2. Microbial Changes in the Maternal Microbiome during Pregnancy
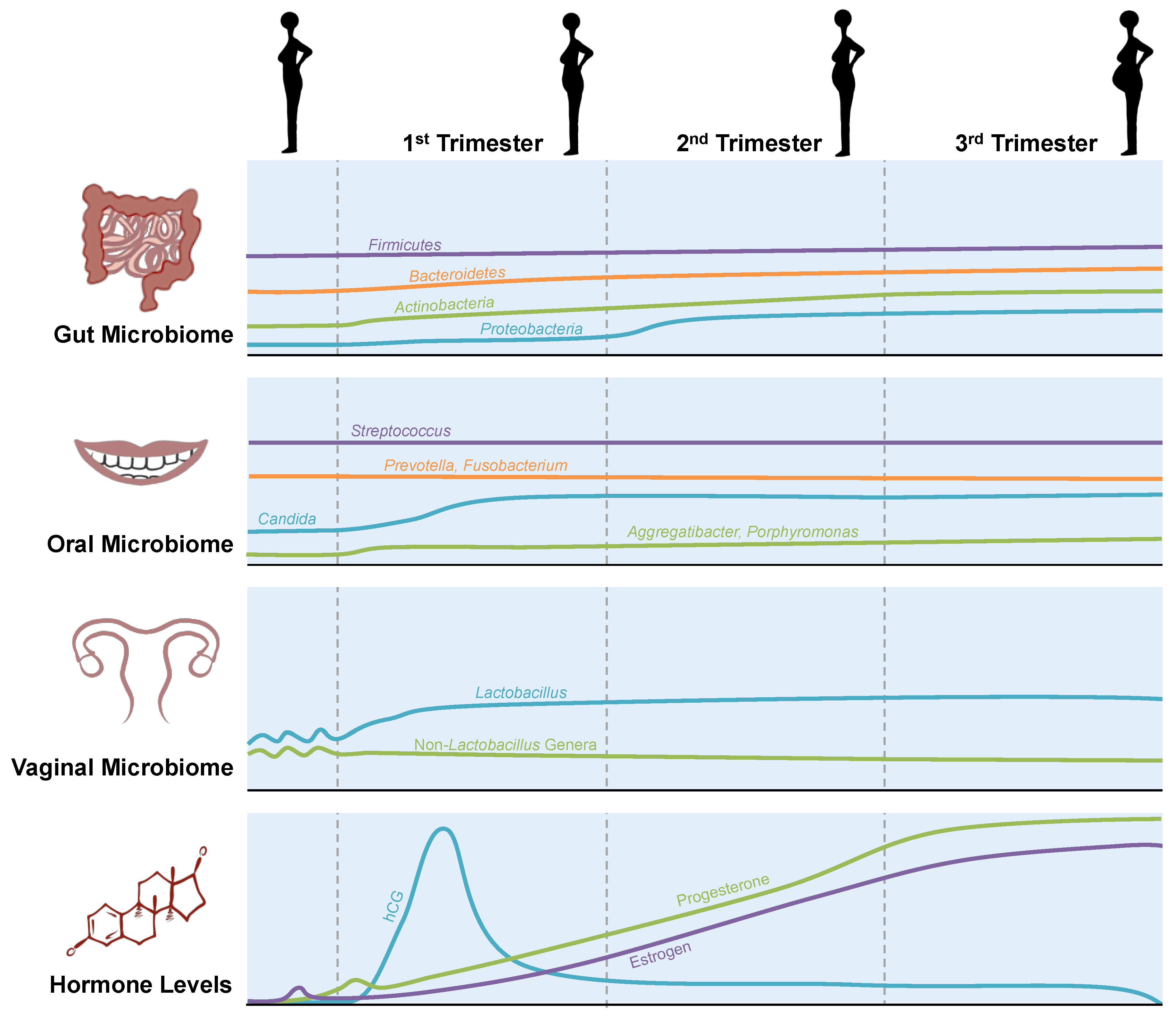
2.1. Gut Microbiome
2.2. Oral Microbiome
2.3. Vaginal Microbiome
2.4. Effects of Maternal Dysbiosis during Pregnancy on the Offspring
2.5. Metabolic Changes in Pregnancy and the Gut Microbiome
2.6. Changes in Gut Permeability during Pregnancy
3. Clinical Implications of the Gut Microbiome in Pregnancy
3.1. Gestational Diabetes
3.2. Preeclampsia
3.3. Fetal Growth Restriction
3.4. Infections during Pregnancy, Clinical Outcomes, and Therapeutics
3.5. Bacterial Infections
3.6. Viral Infections
3.7. Parasitic Infections
3.8. Treatments and Therapeutics
4. Limitations in Animal Models of Pregnancy
5. Conclusions
Funding
Conflicts of Interest
References
- Kumar, P.; Magon, N. Hormones in pregnancy. Niger. Med. J. 2012, 53, 179–183. [Google Scholar] [PubMed]
- Mulak, A.; Tache, Y.; Larauche, M. Sex hormones in the modulation of irritable bowel syndrome. World J. Gastroenterol. 2014, 20, 2433–2448. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Gomez, E.; Gonzalez-Pedrajo, B.; Camacho-Arroyo, I. Role of sex steroid hormones in bacterial-host interactions. BioMed Res. Int. 2013, 2013, 928290. [Google Scholar] [CrossRef] [PubMed]
- Nuriel-Ohayon, M.; Neuman, H.; Ziv, O.; Belogolovski, A.; Barsheshet, Y.; Bloch, N.; Uzan, A.; Lahav, R.; Peretz, A.; Frishman, S.; et al. Progesterone increases bifidobacterium relative abundance during late pregnancy. Cell Rep. 2019, 27, 730–736 e733. [Google Scholar] [CrossRef] [Green Version]
- Koren, O.; Goodrich, J.K.; Cullender, T.C.; Spor, A.; Laitinen, K.; Backhed, H.K.; Gonzalez, A.; Werner, J.J.; Angenent, L.T.; Knight, R.; et al. Host remodeling of the gut microbiome and metabolic changes during pregnancy. Cell 2012, 150, 470–480. [Google Scholar] [CrossRef] [Green Version]
- Gohir, W.; Whelan, F.J.; Surette, M.G.; Moore, C.; Schertzer, J.D.; Sloboda, D.M. Pregnancy-related changes in the maternal gut microbiota are dependent upon the mother’s periconceptional diet. Gut Microbes 2015, 6, 310–320. [Google Scholar] [CrossRef] [Green Version]
- DiGiulio, D.B.; Callahan, B.J.; McMurdie, P.J.; Costello, E.K.; Lyell, D.J.; Robaczewska, A.; Sun, C.L.; Goltsman, D.S.; Wong, R.J.; Shaw, G.; et al. Temporal and spatial variation of the human microbiota during pregnancy. Proc. Natl. Acad Sci. USA 2015, 112, 11060–11065. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.; Azhar, E.I.; Abbas, A.T.; Kumosani, T.; Barbour, E.K.; Raoult, D.; Yasir, M. Metagenomic analysis of antibiotic-induced changes in gut microbiota in a pregnant rat model. Front. Pharmacol. 2016, 7, 104. [Google Scholar] [CrossRef] [Green Version]
- Deo, P.N.; Deshmukh, R. Oral microbiome: Unveiling the fundamentals. J. Oral Maxillofac Pathol. 2019, 23, 122–128. [Google Scholar]
- Perera, M.; Al-Hebshi, N.N.; Speicher, D.J.; Perera, I.; Johnson, N.W. Emerging role of bacteria in oral carcinogenesis: A review with special reference to perio-pathogenic bacteria. J. Oral Microbiol. 2016, 8, 32762. [Google Scholar] [CrossRef] [Green Version]
- Borgo, P.V.; Rodrigues, V.A.; Feitosa, A.C.; Xavier, K.C.; Avila-Campos, M.J. Association between periodontal condition and subgingival microbiota in women during pregnancy: A longitudinal study. J. Appl. Oral Sci. 2014, 22, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, N.; Tsuruda, K.; Iwamoto, Y.; Kato, F.; Odaki, T.; Yamane, N.; Hori, Y.; Harashima, Y.; Sakoda, A.; Tagaya, A.; et al. Significant increase of oral bacteria in the early pregnancy period in japanese women. J. Investig. Clin. Dent. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Hill, G.B. Preterm birth: Associations with genital and possibly oral microflora. Ann. Periodontol. 1998, 3, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, N.; Ekuni, D.; Irie, K.; Furuta, M.; Tomofuji, T.; Morita, M.; Watanabe, T. Relationship between periodontal inflammation and fetal growth in pregnant women: A cross-sectional study. Arch. Gynecol. Obstet. 2013, 287, 951–957. [Google Scholar] [CrossRef]
- Cobb, C.M.; Kelly, P.J.; Williams, K.B.; Babbar, S.; Angolkar, M.; Derman, R.J. The oral microbiome and adverse pregnancy outcomes. Int. J. Womens Health 2017, 9, 551–559. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.W.; Fardini, Y.; Chen, C.; Iacampo, K.G.; Peraino, V.A.; Shamonki, J.M.; Redline, R.W. Term stillbirth caused by oral fusobacterium nucleatum. Obstet. Gynecol. 2010, 115, 442–445. [Google Scholar] [CrossRef] [Green Version]
- Coppenhagen-Glazer, S.; Sol, A.; Abed, J.; Naor, R.; Zhang, X.; Han, Y.W.; Bachrach, G. Fap2 of fusobacterium nucleatum is a galactose-inhibitable adhesin involved in coaggregation, cell adhesion, and preterm birth. Infect. Immun. 2015, 83, 1104–1113. [Google Scholar] [CrossRef] [Green Version]
- Figuero, E.; Han, Y.W.; Furuichi, Y. Periodontal diseases and adverse pregnancy outcomes: Mechanisms. Periodontol. 2000 2020, 83, 175–188. [Google Scholar] [CrossRef]
- Cao, B.; Stout, M.J.; Lee, I.; Mysorekar, I.U. Placental microbiome and its role in preterm birth. Neoreviews 2014, 15, e537–e545. [Google Scholar] [CrossRef] [Green Version]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4680–4687. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Hansmann, M.A.; Davis, C.C.; Suzuki, H.; Brown, C.J.; Schutte, U.; Pierson, J.D.; Forney, L.J. The vaginal bacterial communities of japanese women resemble those of women in other racial groups. FEMS Immunol. Med. Microbiol. 2010, 58, 169–181. [Google Scholar] [CrossRef]
- Zhou, X.; Brown, C.J.; Abdo, Z.; Davis, C.C.; Hansmann, M.A.; Joyce, P.; Foster, J.A.; Forney, L.J. Differences in the composition of vaginal microbial communities found in healthy caucasian and black women. ISME J. 2007, 1, 121–133. [Google Scholar] [CrossRef]
- Verstraelen, H.; Verhelst, R.; Claeys, G.; Temmerman, M.; Vaneechoutte, M. Culture-independent analysis of vaginal microflora: The unrecognized association of atopobium vaginae with bacterial vaginosis. Am. J. Obstet. Gynecol. 2004, 191, 1130–1132. [Google Scholar] [CrossRef] [PubMed]
- Verhelst, R.; Verstraelen, H.; Claeys, G.; Verschraegen, G.; Delanghe, J.; Van Simaey, L.; De Ganck, C.; Temmerman, M.; Vaneechoutte, M. Cloning of 16s rrna genes amplified from normal and disturbed vaginal microflora suggests a strong association between atopobium vaginae, gardnerella vaginalis and bacterial vaginosis. BMC Microbiol. 2004, 4, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredricks, D.N.; Fiedler, T.L.; Marrazzo, J.M. Molecular identification of bacteria associated with bacterial vaginosis. N. Engl. J. Med. 2005, 353, 1899–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aagaard, K.; Riehle, K.; Ma, J.; Segata, N.; Mistretta, T.A.; Coarfa, C.; Raza, S.; Rosenbaum, S.; Van den Veyver, I.; Milosavljevic, A.; et al. A metagenomic approach to characterization of the vaginal microbiome signature in pregnancy. PLoS ONE 2012, 7, e36466. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.; Hassan, S.S.; Gajer, P.; Tarca, A.L.; Fadrosh, D.W.; Nikita, L.; Galuppi, M.; Lamont, R.F.; Chaemsaithong, P.; Miranda, J.; et al. The composition and stability of the vaginal microbiota of normal pregnant women is different from that of non-pregnant women. Microbiome 2014, 2, 4. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; DiGiulio, D.B.; Goltsman, D.S.A.; Sun, C.L.; Costello, E.K.; Jeganathan, P.; Biggio, J.R.; Wong, R.J.; Druzin, M.L.; Shaw, G.M.; et al. Replication and refinement of a vaginal microbial signature of preterm birth in two racially distinct cohorts of us women. Proc. Natl. Acad. Sci. USA 2017, 114, 9966–9971. [Google Scholar] [CrossRef] [Green Version]
- Fettweis, J.M.; Serrano, M.G.; Brooks, J.P.; Edwards, D.J.; Girerd, P.H.; Parikh, H.I.; Huang, B.; Arodz, T.J.; Edupuganti, L.; Glascock, A.L.; et al. The vaginal microbiome and preterm birth. Nat. Med. 2019, 25, 1012–1021. [Google Scholar] [CrossRef] [Green Version]
- Stout, M.J.; Zhou, Y.; Wylie, K.M.; Tarr, P.I.; Macones, G.A.; Tuuli, M.G. Early pregnancy vaginal microbiome trends and preterm birth. Am. J. Obstet. Gynecol. 2017, 217, e351–e356. [Google Scholar] [CrossRef]
- MacIntyre, D.A.; Chandiramani, M.; Lee, Y.S.; Kindinger, L.; Smith, A.; Angelopoulos, N.; Lehne, B.; Arulkumaran, S.; Brown, R.; Teoh, T.G.; et al. The vaginal microbiome during pregnancy and the postpartum period in a european population. Sci. Rep. 2015, 5, 8988. [Google Scholar] [CrossRef] [Green Version]
- Nanthakumar, N.; Meng, D.; Goldstein, A.M.; Zhu, W.; Lu, L.; Uauy, R.; Llanos, A.; Claud, E.C.; Walker, W.A. The mechanism of excessive intestinal inflammation in necrotizing enterocolitis: An immature innate immune response. PLoS ONE 2011, 6, e17776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuppala, V.S.; Meinzen-Derr, J.; Morrow, A.L.; Schibler, K.R. Prolonged initial empirical antibiotic treatment is associated with adverse outcomes in premature infants. J. Pediatr. 2011, 159, 720–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niebyl, J.R. Antibiotics and other anti-infective agents in pregnancy and lactation. Am. J. Perinatol. 2003, 20, 405–414. [Google Scholar] [PubMed]
- Zhang, D.; Huang, Y.; Ye, D. Intestinal dysbiosis: An emerging cause of pregnancy complications? Med. Hypotheses 2015, 84, 223–226. [Google Scholar] [CrossRef]
- Wang, J.; Gu, X.; Yang, J.; Wei, Y.; Zhao, Y. Gut microbiota dysbiosis and increased plasma lps and tmao levels in patients with preeclampsia. Front. Cell. Infect. Microbiol. 2019, 9, 409. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zheng, J.; Shi, W.; Du, N.; Xu, X.; Zhang, Y.; Ji, P.; Zhang, F.; Jia, Z.; Wang, Y.; et al. Dysbiosis of maternal and neonatal microbiota associated with gestational diabetes mellitus. Gut 2018, 67, 1614–1625. [Google Scholar] [CrossRef]
- Crusell, M.K.W.; Hansen, T.H.; Nielsen, T.; Allin, K.H.; Rühlemann, M.C.; Damm, P.; Vestergaard, H.; Rørbye, C.; Jørgensen, N.R.; Christiansen, O.B.; et al. Gestational diabetes is associated with change in the gut microbiota composition in third trimester of pregnancy and postpartum. Microbiome 2018, 6, 89. [Google Scholar] [CrossRef]
- Gomez-Arango, L.F.; Barrett, H.L.; McIntyre, H.D.; Callaway, L.K.; Morrison, M.; Dekker Nitert, M. Increased systolic and diastolic blood pressure is associated with altered gut microbiota composition and butyrate production in early pregnancy. Hypertension 2016, 68, 974–981. [Google Scholar] [CrossRef]
- Chen, X.; Li, P.; Liu, M.; Zheng, H.; He, Y.; Chen, M.X.; Tang, W.; Yue, X.; Huang, Y.; Zhuang, L.; et al. Gut dysbiosis induces the development of pre-eclampsia through bacterial translocation. Gut 2020, 69, 513–522. [Google Scholar] [CrossRef]
- Lv, L.J.; Li, S.H.; Li, S.C.; Zhong, Z.C.; Duan, H.L.; Tian, C.; Li, H.; He, W.; Chen, M.C.; He, T.W.; et al. Early-onset preeclampsia is associated with gut microbial alterations in antepartum and postpartum women. Front. Cell. Infect. Microbiol. 2019, 9, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, C.; Eichelberger, K. Maternal microbiome and pregnancy outcomes. Fertil. Steril. 2015, 104, 1358–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldassarre, M.E.; Di Mauro, A.; Capozza, M.; Rizzo, V.; Schettini, F.; Panza, R.; Laforgia, N. Dysbiosis and prematurity: Is there a role for probiotics? Nutrients 2019, 11, 1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhasan, M.M.; Cait, A.M.; Heimesaat, M.M.; Blaut, M.; Klopfleisch, R.; Wedel, A.; Conlon, T.M.; Yildirim, A.Ö.; Sodemann, E.B.; Mohn, W.W.; et al. Antibiotic use during pregnancy increases offspring asthma severity in a dose-dependent manner. Allergy 2020, 75, 1979–1990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaevska, E.; Popovic, M.; Pizzi, C.; Fiano, V.; Rusconi, F.; Merletti, F.; Richiardi, L.; Maule, M. Maternal antibiotic use and vaginal infections in the third trimester of pregnancy and the risk of obesity in preschool children. Pediatr. Obes. 2020, 15, e12632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamont, R.F.; Møller Luef, B.; Stener Jørgensen, J. Childhood inflammatory and metabolic disease following exposure to antibiotics in pregnancy, antenatally, intrapartum and neonatally. F1000Research 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Perez, G.; Hicks, A.L.; Tekieli, T.M.; Radens, C.M.; Williams, B.L.; Lamouse-Smith, E.S. Maternal antibiotic treatment impacts development of the neonatal intestinal microbiome and antiviral immunity. J. Immunol. 2016, 196, 3768–3779. [Google Scholar] [CrossRef] [Green Version]
- Paul, H.A.; Bomhof, M.R.; Vogel, H.J.; Reimer, R.A. Diet-induced changes in maternal gut microbiota and metabolomic profiles influence programming of offspring obesity risk in rats. Sci. Rep. 2016, 6, 20683. [Google Scholar] [CrossRef]
- Rautava, S.; Collado, M.C.; Salminen, S.; Isolauri, E. Probiotics modulate host-microbe interaction in the placenta and fetal gut: A randomized, double-blind, placebo-controlled trial. Neonatology 2012, 102, 178–184. [Google Scholar] [CrossRef]
- Abrahamsson, T.R.; Wu, R.Y.; Jenmalm, M.C. Gut microbiota and allergy: The importance of the pregnancy period. Pediatr. Res. 2015, 77, 214–219. [Google Scholar] [CrossRef] [Green Version]
- Levan, S.R.; Stamnes, K.A.; Lin, D.L.; Panzer, A.R.; Fukui, E.; McCauley, K.; Fujimura, K.E.; McKean, M.; Ownby, D.R.; Zoratti, E.M.; et al. Elevated faecal 12,13-dihome concentration in neonates at high risk for asthma is produced by gut bacteria and impedes immune tolerance. Nat. Microbiol. 2019, 4, 1851–1861. [Google Scholar] [CrossRef] [PubMed]
- Sharon, G.; Cruz, N.J.; Kang, D.W.; Gandal, M.J.; Wang, B.; Kim, Y.M.; Zink, E.M.; Casey, C.P.; Taylor, B.C.; Lane, C.J.; et al. Human gut microbiota from autism spectrum disorder promote behavioral symptoms in mice. Cell 2019, 177, 1600–1618 e1617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Chow, J.; Lee, S.M.; Shen, Y.; Khosravi, A.; Mazmanian, S.K. Host-bacterial symbiosis in health and disease. Adv. Immunol. 2010, 107, 243–274. [Google Scholar]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial degradation of complex carbohydrates in the gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef] [Green Version]
- Zimmet, P.; Alberti, K.G.; Shaw, J. Global and societal implications of the diabetes epidemic. Nature 2001, 414, 782–787. [Google Scholar] [CrossRef]
- Ponzo, V.; Fedele, D.; Goitre, I.; Leone, F.; Lezo, A.; Monzeglio, C.; Finocchiaro, C.; Ghigo, E.; Bo, S. Diet-gut microbiota interactions and gestational diabetes mellitus (gdm). Nutrients 2019, 11, 330. [Google Scholar] [CrossRef] [Green Version]
- Collado, M.C.; Isolauri, E.; Laitinen, K.; Salminen, S. Distinct composition of gut microbiota during pregnancy in overweight and normal-weight women. Am. J. Clin. Nutr. 2008, 88, 894–899. [Google Scholar] [CrossRef]
- Choi, Y.; Kwon, Y.; Kim, D.K.; Jeon, J.; Jang, S.C.; Wang, T.; Ban, M.; Kim, M.H.; Jeon, S.G.; Kim, M.S.; et al. Gut microbe-derived extracellular vesicles induce insulin resistance, thereby impairing glucose metabolism in skeletal muscle. Sci. Rep. 2015, 5, 15878. [Google Scholar] [CrossRef] [Green Version]
- Wallace, J.G.; Bellissimo, C.J.; Yeo, E.; Fei Xia, Y.; Petrik, J.J.; Surette, M.G.; Bowdish, D.M.E.; Sloboda, D.M. Obesity during pregnancy results in maternal intestinal inflammation, placental hypoxia, and alters fetal glucose metabolism at mid-gestation. Sci. Rep. 2019, 9, 17621. [Google Scholar] [CrossRef]
- Gohir, W.; Kennedy, K.M.; Wallace, J.G.; Saoi, M.; Bellissimo, C.J.; Britz-McKibbin, P.; Petrik, J.J.; Surette, M.G.; Sloboda, D.M. High-fat diet intake modulates maternal intestinal adaptations to pregnancy and results in placental hypoxia, as well as altered fetal gut barrier proteins and immune markers. J. Physiol. 2019, 597, 3029–3051. [Google Scholar] [CrossRef]
- Cani, P.D.; Possemiers, S.; Van de Wiele, T.; Guiot, Y.; Everard, A.; Rottier, O.; Geurts, L.; Naslain, D.; Neyrinck, A.; Lambert, D.M.; et al. Changes in gut microbiota control inflammation in obese mice through a mechanism involving glp-2-driven improvement of gut permeability. Gut 2009, 58, 1091–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeyabalan, A. Epidemiology of preeclampsia: Impact of obesity. Nutr. Rev. 2013, 71 (Suppl. 1), S18–S25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grill, S.; Rusterholz, C.; Zanetti-Dallenbach, R.; Tercanli, S.; Holzgreve, W.; Hahn, S.; Lapaire, O. Potential markers of preeclampsia—A review. Reprod. Biol. Endocrinol. 2009, 7, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aagaard, K.; Ma, J.; Antony, K.M.; Ganu, R.; Petrosino, J.; Versalovic, J. The placenta harbors a unique microbiome. Sci. Transl. Med. 2014, 6, 237–265. [Google Scholar] [CrossRef] [Green Version]
- Beckers, K.F.; Sones, J.L. Maternal microbiome and the hypertensive disorder of pregnancy, preeclampsia. Am. J. Physiol. Heart Circ. Physiol. 2020, 318, H1–H10. [Google Scholar] [CrossRef]
- Amarasekara, R.; Jayasekara, R.W.; Senanayake, H.; Dissanayake, V.H. Microbiome of the placenta in pre-eclampsia supports the role of bacteria in the multifactorial cause of pre-eclampsia. J. Obstet. Gynaecol. Res. 2015, 41, 662–669. [Google Scholar] [CrossRef]
- Barak, S.; Oettinger-Barak, O.; Machtei, E.E.; Sprecher, H.; Ohel, G. Evidence of periopathogenic microorganisms in placentas of women with preeclampsia. J. Periodontol. 2007, 78, 670–676. [Google Scholar] [CrossRef]
- Romo, A.; Carceller, R.; Tobajas, J. Intrauterine growth retardation (iugr): Epidemiology and etiology. Pediatr. Endocrinol. Rev. 2009, 6 (Suppl. 3), 332–336. [Google Scholar]
- Committee on Practice Bulletins--Gynecology, A.C.o.O.; Gynecologists, W.D.C.U.S.A. Intrauterine growth restriction. Clinical management guidelines for obstetrician-gynecologists. American college of obstetricians and gynecologists. Int. J. Gynaecol. Obstet. 2001, 72, 85–96. [Google Scholar]
- Sharma, D.; Shastri, S.; Sharma, P. Intrauterine growth restriction: Antenatal and postnatal aspects. Clin. Med. Insights Pediatr. 2016, 10, 67–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Den Hollander, W.J.; Schalekamp-Timmermans, S.; Holster, I.L.; Jaddoe, V.W.; Hofman, A.; Moll, H.A.; Perez-Perez, G.I.; Blaser, M.J.; Steegers, E.A.; Kuipers, E.J. Helicobacter pylori colonization and pregnancies complicated by preeclampsia, spontaneous prematurity, and small for gestational age birth. Helicobacter 2017, 22, e12364. [Google Scholar] [CrossRef] [PubMed]
- Vedmedovska, N.; Rezeberga, D.; Donder, G.G.G. Is abnormal vaginal microflora a risk factor for intrauterine fetal growth restriction? Asian Pac. J. Reprod. 2015, 4, 313–316. [Google Scholar] [CrossRef]
- Zhang, W.; Ma, C.; Xie, P.; Zhu, Q.; Wang, X.; Yin, Y.; Kong, X. Gut microbiota of newborn piglets with intrauterine growth restriction have lower diversity and different taxonomic abundances. J. Appl. Microbiol. 2019, 127, 354–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kourtis, A.P.; Read, J.S.; Jamieson, D.J. Pregnancy and infection. N. Engl. J. Med. 2014, 370, 2211–2218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, C.J.; Barrett, F.F.; Gordon, R.C.; Yow, M.D. Suppurative meningitis due to streptococci of lancefield group b: A study of 33 infants. J. Pediatr. 1973, 82, 724–729. [Google Scholar] [CrossRef]
- Moller, M.; Thomsen, A.C.; Borch, K.; Dinesen, K.; Zdravkovic, M. Rupture of fetal membranes and premature delivery associated with group b streptococci in urine of pregnant women. Lancet 1984, 2, 69–70. [Google Scholar] [CrossRef]
- Pylipow, M.; Gaddis, M.; Kinney, J.S. Selective intrapartum prophylaxis for group b streptococcus colonization: Management and outcome of newborns. Pediatrics 1994, 93, 631–635. [Google Scholar] [CrossRef]
- Kim, E.J.; Oh, K.Y.; Kim, M.Y.; Seo, Y.S.; Shin, J.H.; Song, Y.R.; Yang, J.H.; Foxman, B.; Ki, M. Risk factors for group b streptococcus colonization among pregnant women in korea. Epidemiol. Health 2011, 33, e2011010. [Google Scholar] [CrossRef] [Green Version]
- Tam, T.; Bilinski, E.; Lombard, E. Recolonization of group b streptococcus (gbs) in women with prior gbs genital colonization in pregnancy. J. Matern. Fetal Neonatal Med. 2012, 25, 1987–1989. [Google Scholar] [CrossRef]
- Nan, C.; Dangor, Z.; Cutland, C.L.; Edwards, M.S.; Madhi, S.A.; Cunnington, M.C. Maternal group b streptococcus-related stillbirth: A systematic review. BJOG 2015, 122, 1437–1445. [Google Scholar] [CrossRef] [PubMed]
- Vornhagen, J.; Adams Waldorf, K.M.; Rajagopal, L. Perinatal group b streptococcal infections: Virulence factors, immunity, and prevention strategies. Trends Microbiol. 2017, 25, 919–931. [Google Scholar] [CrossRef] [PubMed]
- Jackson, K.A.; Iwamoto, M.; Swerdlow, D. Pregnancy-associated listeriosis. Epidemiol. Infect. 2010, 138, 1503–1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mylonakis, E.; Paliou, M.; Hohmann, E.L.; Calderwood, S.B.; Wing, E.J. Listeriosis during pregnancy: A case series and review of 222 cases. Medicine 2002, 81, 260–269. [Google Scholar] [CrossRef]
- Temple, M.E.; Nahata, M.C. Treatment of listeriosis. Ann. Pharmacother. 2000, 34, 656–661. [Google Scholar] [CrossRef]
- Haider, G.; Zehra, N.; Munir, A.A.; Haider, A. Risk factors of urinary tract infection in pregnancy. J. Pak. Med. Assoc. 2010, 60, 213–216. [Google Scholar]
- Reeves, D.S. Treatment of bacteriuria in pregnancy with single dose fosfomycin trometamol: A review. Infection 1992, 20 (Suppl. 4), S313–S316. [Google Scholar] [CrossRef]
- Sharami, S.H.; Afrakhteh, M.; Shakiba, M. Urinary tract infections in pregnant women with bacterial vaginosis. J. Obstet. Gynaecol. 2007, 27, 252–254. [Google Scholar] [CrossRef]
- Brocklehurst, P.; Gordon, A.; Heatley, E.; Milan, S.J. Antibiotics for treating bacterial vaginosis in pregnancy. Cochrane Database Syst. Rev. 2013, CD000262. [Google Scholar] [CrossRef] [Green Version]
- Georgijevic, A.; Cjukic-Ivancevic, S.; Bujko, M. Bacterial vaginosis. Epidemiology and risk factors. Srp. Arh. Celok. Lek. 2000, 128, 29–33. [Google Scholar]
- Hay, P.E.; Lamont, R.F.; Taylor-Robinson, D.; Morgan, D.J.; Ison, C.; Pearson, J. Abnormal bacterial colonisation of the genital tract and subsequent preterm delivery and late miscarriage. BMJ 1994, 308, 295–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, B.; Brunham, R.C.; Laga, M.; Piot, P.; Ndinya-Achola, J.O.; Maitha, G.; Cheang, M.; Plummer, F.A. Maternal gonococcal infection as a preventable risk factor for low birth weight. J. Infect. Dis. 1990, 161, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Force, U.S.P.S.T.; Curry, S.J.; Krist, A.H.; Owens, D.K.; Barry, M.J.; Caughey, A.B.; Davidson, K.W.; Doubeni, C.A.; Epling, J.W., Jr.; Kemper, A.R.; et al. Ocular prophylaxis for gonococcal ophthalmia neonatorum: Us preventive services task force reaffirmation recommendation statement. JAMA 2019, 321, 394–398. [Google Scholar]
- Kreisel, K.; Weston, E.; Braxton, J.; Llata, E.; Torrone, E. Keeping an eye on chlamydia and gonorrhea conjunctivitis in infants in the united states, 2010–2015. Sex. Transm. Dis. 2017, 44, 356–358. [Google Scholar] [CrossRef] [PubMed]
- Rours, G.I.; Duijts, L.; Moll, H.A.; Arends, L.R.; de Groot, R.; Jaddoe, V.W.; Hofman, A.; Steegers, E.A.; Mackenbach, J.P.; Ott, A.; et al. Chlamydia trachomatis infection during pregnancy associated with preterm delivery: A population-based prospective cohort study. Eur. J. Epidemiol. 2011, 26, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.M.; Sheffield, J.S.; Sanchez, P.J.; Mayfield, J.; Wendel, G.D., Jr. Efficacy of treatment for syphilis in pregnancy. Obstet. Gynecol. 1999, 93, 5–8. [Google Scholar] [PubMed]
- Gomez, G.B.; Kamb, M.L.; Newman, L.M.; Mark, J.; Broutet, N.; Hawkes, S.J. Untreated maternal syphilis and adverse outcomes of pregnancy: A systematic review and meta-analysis. Bull. World Health Organ. 2013, 91, 217–226. [Google Scholar] [CrossRef]
- Zhou, H.; Chen, X.S.; Hong, F.C.; Pan, P.; Yang, F.; Cai, Y.M.; Yin, Y.P.; Peeling, R.W.; Mabey, D. Risk factors for syphilis infection among pregnant women: Results of a case-control study in shenzhen, china. Sex. Transm. Infect. 2007, 83, 476–480. [Google Scholar] [CrossRef] [Green Version]
- Boppana, S.B.; Pass, R.F.; Britt, W.J.; Stagno, S.; Alford, C.A. Symptomatic congenital cytomegalovirus infection: Neonatal morbidity and mortality. Pediatr. Infect. Dis. J. 1992, 11, 93–99. [Google Scholar] [CrossRef]
- Fowler, K.B.; Boppana, S.B. Congenital cytomegalovirus (cmv) infection and hearing deficit. J. Clin. Virol. 2006, 35, 226–231. [Google Scholar] [CrossRef]
- Malm, G.; Engman, M.L. Congenital cytomegalovirus infections. Semin. Fetal Neonatal Med. 2007, 12, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Rawlinson, W.D.; Hamilton, S.T.; van Zuylen, W.J. Update on treatment of cytomegalovirus infection in pregnancy and of the newborn with congenital cytomegalovirus. Curr. Opin. Infect. Dis. 2016, 29, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Stagno, S.; Pass, R.F.; Cloud, G.; Britt, W.J.; Henderson, R.E.; Walton, P.D.; Veren, D.A.; Page, F.; Alford, C.A. Primary cytomegalovirus infection in pregnancy. Incidence, transmission to fetus, and clinical outcome. JAMA 1986, 256, 1904–1908. [Google Scholar] [CrossRef] [PubMed]
- Colvin, H.M.; Mitchell, A.E. Hepatitis and Liver Cancer: A National Strategy for Prevention and Control of Hepatitis b and c; Colvin, H.M., Mitchell, A.E., Eds.; National Academies Press: Washington, DC, USA, 2010. [Google Scholar]
- Perz, J.F.; Armstrong, G.L.; Farrington, L.A.; Hutin, Y.J.; Bell, B.P. The contributions of hepatitis b virus and hepatitis c virus infections to cirrhosis and primary liver cancer worldwide. J. Hepatol. 2006, 45, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, R.L.; Culhane, J.F.; Johnson, D.C. Maternal infection and adverse fetal and neonatal outcomes. Clin. Perinatol. 2005, 32, 523–559. [Google Scholar] [CrossRef] [PubMed]
- Lambert, N.; Strebel, P.; Orenstein, W.; Icenogle, J.; Poland, G.A. Rubella. Lancet 2015, 385, 2297–2307. [Google Scholar] [CrossRef] [Green Version]
- Brasil, P.; Pereira, J.P., Jr.; Moreira, M.E.; Ribeiro Nogueira, R.M.; Damasceno, L.; Wakimoto, M.; Rabello, R.S.; Valderramos, S.G.; Halai, U.A.; Salles, T.S.; et al. Zika virus infection in pregnant women in rio de janeiro. N. Engl. J. Med. 2016, 375, 2321–2334. [Google Scholar] [CrossRef]
- Brown, Z.A.; Selke, S.; Zeh, J.; Kopelman, J.; Maslow, A.; Ashley, R.L.; Watts, D.H.; Berry, S.; Herd, M.; Corey, L. The acquisition of herpes simplex virus during pregnancy. N. Engl. J. Med. 1997, 337, 509–515. [Google Scholar] [CrossRef]
- Triunfol, M. Microcephaly in brazil: Confidence builds in zika connection. Lancet Infect. Dis. 2016, 16, 527–528. [Google Scholar] [CrossRef]
- Braig, S.; Luton, D.; Sibony, O.; Edlinger, C.; Boissinot, C.; Blot, P.; Oury, J.F. Acyclovir prophylaxis in late pregnancy prevents recurrent genital herpes and viral shedding. Eur. J. Obstet. Gynecol. Reprod. Biol. 2001, 96, 55–58. [Google Scholar] [CrossRef]
- Pinninti, S.G.; Kimberlin, D.W. Neonatal herpes simplex virus infections. Semin. Perinatol. 2018, 42, 168–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Straface, G.; Selmin, A.; Zanardo, V.; De Santis, M.; Ercoli, A.; Scambia, G. Herpes simplex virus infection in pregnancy. Infect. Dis. Obstet. Gynecol. 2012, 2012, 385697. [Google Scholar] [CrossRef] [PubMed]
- Tideman, R.L.; Taylor, J.; Marks, C.; Seifert, C.; Berry, G.; Trudinger, B.; Cunningham, A.; Mindel, A. Sexual and demographic risk factors for herpes simplex type 1 and 2 in women attending an antenatal clinic. Sex. Transm. Infect. 2001, 77, 413–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, E.R.; Charurat, M.; Mofenson, L.; Hanson, I.C.; Pitt, J.; Diaz, C.; Hayani, K.; Handelsman, E.; Smeriglio, V.; Hoff, R.; et al. Combination antiretroviral strategies for the treatment of pregnant hiv-1-infected women and prevention of perinatal hiv-1 transmission. J. Acquir. Immune Defic. Syndr. 2002, 29, 484–494. [Google Scholar] [CrossRef]
- Maartens, G.; Celum, C.; Lewin, S.R. Hiv infection: Epidemiology, pathogenesis, treatment, and prevention. Lancet 2014, 384, 258–271. [Google Scholar] [CrossRef]
- Smith, N.H.; Hwang, L.Y. Risk factors for hiv in pregnant women. Int. J. STD AIDS 1996, 7, 388–395. [Google Scholar] [CrossRef]
- Townsend, C.L.; Cortina-Borja, M.; Peckham, C.S.; de Ruiter, A.; Lyall, H.; Tookey, P.A. Low rates of mother-to-child transmission of hiv following effective pregnancy interventions in the united kingdom and ireland, 2000–2006. AIDS 2008, 22, 973–981. [Google Scholar] [CrossRef]
- Havelaar, A.H.; Kemmeren, J.M.; Kortbeek, L.M. Disease burden of congenital toxoplasmosis. Clin. Infect. Dis. 2007, 44, 1467–1474. [Google Scholar] [CrossRef]
- Pappas, G.; Roussos, N.; Falagas, M.E. Toxoplasmosis snapshots: Global status of toxoplasma gondii seroprevalence and implications for pregnancy and congenital toxoplasmosis. Int. J. Parasitol. 2009, 39, 1385–1394. [Google Scholar] [CrossRef]
- Paquet, C.; Yudin, M.H.; Allen, V.M.; Bouchard, C.; Boucher, M.; Caddy, S.; Castillo, E.; Money, D.M.; Murphy, K.E.; Ogilvie, G.; et al. Toxoplasmosis in pregnancy: Prevention, screening, and treatment. J. Obstet. Gynaecol. Can. 2013, 35, 78–81. [Google Scholar] [CrossRef]
- Doran, K.S.; Nizet, V. Molecular pathogenesis of neonatal group b streptococcal infection: No longer in its infancy. Mol. Microbiol. 2004, 54, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Ulett, G.C.; Webb, R.I.; Ulett, K.B.; Cui, X.; Benjamin, W.H.; Crowley, M.; Schembri, M.A. Group b streptococcus (gbs) urinary tract infection involves binding of gbs to bladder uroepithelium and potent but gbs-specific induction of interleukin 1alpha. J. Infect. Dis. 2010, 201, 866–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pass, M.A.; Gray, B.M.; Khare, S.; Dillon, H.C., Jr. Prospective studies of group b streptococcal infections in infants. J. Pediatr. 1979, 95, 437–443. [Google Scholar] [CrossRef]
- Zangwill, K.M.; Schuchat, A.; Wenger, J.D. Group b streptococcal disease in the united states, 1990: Report from a multistate active surveillance system. MMWR CDC Surveill. Summ. 1992, 41, 25–32. [Google Scholar] [PubMed]
- Morgan, J.; Zafar, N.; Cooper, D. Group b Streptococcus and Pregnancy. Available online: https://www.ncbi.nlm.nih.gov/books/NBK482443/ (accessed on 25 October 2020).
- Silver, H.M. Listeriosis during pregnancy. Obstet. Gynecol. Surv. 1998, 53, 737–740. [Google Scholar] [CrossRef] [PubMed]
- Berman, S.M.; Harrison, H.R.; Boyce, W.T.; Haffner, W.J.; Lewis, M.; Arthur, J.B. Low birth weight, prematurity, and postpartum endometritis. Association with prenatal cervical mycoplasma hominis and chlamydia trachomatis infections. JAMA 1987, 257, 1189–1194. [Google Scholar] [CrossRef] [PubMed]
- Baud, D.; Regan, L.; Greub, G. Emerging role of chlamydia and chlamydia-like organisms in adverse pregnancy outcomes. Curr. Opin. Infect. Dis. 2008, 21, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Brunham, R.C.; Binns, B.; McDowell, J.; Paraskevas, M. Chlamydia trachomatis infection in women with ectopic pregnancy. Obstet. Gynecol. 1986, 67, 722–726. [Google Scholar] [CrossRef]
- Livengood, C.H. Bacterial vaginosis: An overview for 2009. Rev. Obstet Gynecol 2009, 2, 28–37. [Google Scholar]
- Baldwin, E.A.; Walther-Antonio, M.; MacLean, A.M.; Gohl, D.M.; Beckman, K.B.; Chen, J.; White, B.; Creedon, D.J.; Chia, N. Persistent microbial dysbiosis in preterm premature rupture of membranes from onset until delivery. PeerJ 2015, 3, e1398. [Google Scholar] [CrossRef]
- Delzell, J.E., Jr.; Lefevre, M.L. Urinary tract infections during pregnancy. Am. Fam. Physician 2000, 61, 713–721. [Google Scholar] [PubMed]
- Goldenberg, R.L.; Hauth, J.C.; Andrews, W.W. Intrauterine infection and preterm delivery. N. Engl. J. Med. 2000, 342, 1500–1507. [Google Scholar] [CrossRef]
- Virgin, H.W. The virome in mammalian physiology and disease. Cell 2014, 157, 142–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins-McMillen, D.; Buehler, J.; Peppenelli, M.; Goodrum, F. Molecular determinants and the regulation of human cytomegalovirus latency and reactivation. Viruses 2018, 10, 444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maidji, E.; McDonagh, S.; Genbacev, O.; Tabata, T.; Pereira, L. Maternal antibodies enhance or prevent cytomegalovirus infection in the placenta by neonatal fc receptor-mediated transcytosis. Am. J. Pathol. 2006, 168, 1210–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhat, P.; Anderson, D.A. Hepatitis b virus translocates across a trophoblastic barrier. J. Virol. 2007, 81, 7200–7207. [Google Scholar] [CrossRef] [Green Version]
- Delorme-Axford, E.; Sadovsky, Y.; Coyne, C.B. The placenta as a barrier to viral infections. Annu. Rev. Virol. 2014, 1, 133–146. [Google Scholar] [CrossRef]
- Prasad, M.R.; Honegger, J.R. Hepatitis c virus in pregnancy. Am. J. Perinatol. 2013, 30, 149–159. [Google Scholar]
- Tondury, G.; Smith, D.W. Fetal rubella pathology. J. Pediatr. 1966, 68, 867–879. [Google Scholar] [CrossRef]
- Toizumi, M.; Do, C.G.T.; Motomura, H.; Do, T.N.; Fukunaga, H.; Iijima, M.; Le, N.N.; Nguyen, H.T.; Moriuchi, H.; Yoshida, L.M. Characteristics of patent ductus arteriosus in congenital rubella syndrome. Sci. Rep. 2019, 9, 17105. [Google Scholar] [CrossRef]
- Breastfeeding and HIV International Transmission Study Group; Coutsoudis, A.; Dabis, F.; Fawzi, W.; Gaillard, P.; Haverkamp, G.; Harris, D.R.; Jackson, J.B.; Leroy, V.; Meda, N.; et al. Late postnatal transmission of hiv-1 in breast-fed children: An individual patient data meta-analysis. J. Infect. Dis. 2004, 189, 2154–2166. [Google Scholar] [PubMed] [Green Version]
- Reynolds, M.R.; Jones, A.M.; Petersen, E.E.; Lee, E.H.; Rice, M.E.; Bingham, A.; Ellington, S.R.; Evert, N.; Reagan-Steiner, S.; Oduyebo, T.; et al. Vital signs: Update on zika virus-associated birth defects and evaluation of all u.S. Infants with congenital zika virus exposure—U.S. Zika pregnancy registry, 2016. MMWR Morb. Mortal. Wkly. Rep. 2017, 66, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Cauchemez, S.; Besnard, M.; Bompard, P.; Dub, T.; Guillemette-Artur, P.; Eyrolle-Guignot, D.; Salje, H.; Van Kerkhove, M.D.; Abadie, V.; Garel, C.; et al. Association between zika virus and microcephaly in french polynesia, 2013–2015: A retrospective study. Lancet 2016, 387, 2125–2132. [Google Scholar] [CrossRef] [Green Version]
- Driggers, R.W.; Ho, C.Y.; Korhonen, E.M.; Kuivanen, S.; Jaaskelainen, A.J.; Smura, T.; Rosenberg, A.; Hill, D.A.; DeBiasi, R.L.; Vezina, G.; et al. Zika virus infection with prolonged maternal viremia and fetal brain abnormalities. N. Engl. J. Med. 2016, 374, 2142–2151. [Google Scholar] [CrossRef]
- Mlakar, J.; Korva, M.; Tul, N.; Popovic, M.; Poljsak-Prijatelj, M.; Mraz, J.; Kolenc, M.; Resman Rus, K.; Vesnaver Vipotnik, T.; Fabjan Vodusek, V.; et al. Zika virus associated with microcephaly. N. Engl. J. Med. 2016, 374, 951–958. [Google Scholar] [CrossRef]
- Cotch, M.F.; Pastorek, J.G., 2nd; Nugent, R.P.; Hillier, S.L.; Gibbs, R.S.; Martin, D.H.; Eschenbach, D.A.; Edelman, R.; Carey, J.C.; Regan, J.A.; et al. Trichomonas vaginalis associated with low birth weight and preterm delivery. The vaginal infections and prematurity study group. Sex. Transm. Dis. 1997, 24, 353–360. [Google Scholar] [CrossRef]
- Lockwood, C.J.; Magriples, U. Prenatal care: Initial assessment. In Uptodate; Berghella, V., Ed.; Wolters Kluwer: Waltham, MA, USA, 2020. [Google Scholar]
- Lockwood, C.J.; Magriples, U. Prenatal care: Second and third trimesters. In Uptodate; Wolters Kluwer: Waltham, MA, USA, 2020. [Google Scholar]
- Santos, F.; Oraichi, D.; Berard, A. Prevalence and predictors of anti-infective use during pregnancy. Pharmacoepidemiol. Drug Saf. 2010, 19, 418–427. [Google Scholar] [CrossRef]
- Bookstaver, P.B.; Bland, C.M.; Griffin, B.; Stover, K.R.; Eiland, L.S.; McLaughlin, M. A review of antibiotic use in pregnancy. Pharmacotherapy 2015, 35, 1052–1062. [Google Scholar] [CrossRef]
- Matok, I.; Gorodischer, R.; Koren, G.; Landau, D.; Wiznitzer, A.; Levy, A. Exposure to folic acid antagonists during the first trimester of pregnancy and the risk of major malformations. Br. J. Clin. Pharmacol. 2009, 68, 956–962. [Google Scholar] [CrossRef] [Green Version]
- Schrag, S.J.; Zywicki, S.; Farley, M.M.; Reingold, A.L.; Harrison, L.H.; Lefkowitz, L.B.; Hadler, J.L.; Danila, R.; Cieslak, P.R.; Schuchat, A. Group b streptococcal disease in the era of intrapartum antibiotic prophylaxis. N. Engl. J. Med. 2000, 342, 15–20. [Google Scholar] [CrossRef]
- Watson, J.C.; Hadler, S.C.; Dykewicz, C.A.; Reef, S.; Phillips, L. Measles, mumps, and rubella—Vaccine use and strategies for elimination of measles, rubella, and congenital rubella syndrome and control of mumps: Recommendations of the advisory committee on immunization practices (acip). MMWR Recomm. Rep. 1998, 47, 1–57. [Google Scholar]
- Biron, K.K. Antiviral drugs for cytomegalovirus diseases. Antivir. Res. 2006, 71, 154–163. [Google Scholar] [CrossRef]
- Brown, Z.A.; Gardella, C.; Wald, A.; Morrow, R.A.; Corey, L. Genital herpes complicating pregnancy. Obstet. Gynecol. 2005, 106, 845–856. [Google Scholar] [CrossRef]
- Nuccitelli, A.; Cozzi, R.; Gourlay, L.J.; Donnarumma, D.; Necchi, F.; Norais, N.; Telford, J.L.; Rappuoli, R.; Bolognesi, M.; Maione, D.; et al. Structure-based approach to rationally design a chimeric protein for an effective vaccine against group b streptococcus infections. Proc. Natl. Acad. Sci. USA 2011, 108, 10278–10283. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Schrag, S.J.; Alderson, M.R.; Madhi, S.A.; Baker, C.J.; Sobanjo-Ter Meulen, A.; Kaslow, D.C.; Smith, P.G.; Moorthy, V.S.; Vekemans, J. Who consultation on group b streptococcus vaccine development: Report from a meeting held on 27–28 april 2016. Vaccine 2019, 37, 7307–7314. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.S.; Jenks, J.A.; Pardi, N.; Goodwin, M.; Roark, H.; Edwards, W.; McLellan, J.S.; Pollara, J.; Weissman, D.; Permar, S.R. Human cytomegalovirus glycoprotein b nucleoside-modified mrna vaccine elicits antibody responses with greater durability and breadth than mf59-adjuvanted gb protein immunization. J. Virol. 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- Fowler, K.B.; Stagno, S.; Pass, R.F. Maternal immunity and prevention of congenital cytomegalovirus infection. JAMA 2003, 289, 1008–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miner, J.J.; Cao, B.; Govero, J.; Smith, A.M.; Fernandez, E.; Cabrera, O.H.; Garber, C.; Noll, M.; Klein, R.S.; Noguchi, K.K.; et al. Zika virus infection during pregnancy in mice causes placental damage and fetal demise. Cell 2016, 165, 1081–1091. [Google Scholar] [CrossRef] [Green Version]
- Shan, C.; Muruato, A.E.; Nunes, B.T.D.; Luo, H.; Xie, X.; Medeiros, D.B.A.; Wakamiya, M.; Tesh, R.B.; Barrett, A.D.; Wang, T.; et al. A live-attenuated zika virus vaccine candidate induces sterilizing immunity in mouse models. Nat. Med. 2017, 23, 763–767. [Google Scholar] [CrossRef]
- Larocca, R.A.; Abbink, P.; Peron, J.P.; Zanotto, P.M.; Iampietro, M.J.; Badamchi-Zadeh, A.; Boyd, M.; Ng’ang’a, D.; Kirilova, M.; Nityanandam, R.; et al. Vaccine protection against zika virus from brazil. Nature 2016, 536, 474–478. [Google Scholar] [CrossRef]
- Bonney, E.A. Demystifying animal models of adverse pregnancy outcomes: Touching bench and bedside. Am. J. Reprod. Immunol. 2013, 69, 567–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clancy, B.; Finlay, B.L.; Darlington, R.B.; Anand, K.J. Extrapolating brain development from experimental species to humans. Neurotoxicology 2007, 28, 931–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, R.; Martin-Fairey, C.; Sojka, D.K.; Herzog, E.D.; Jungheim, E.S.; Stout, M.J.; Fay, J.C.; Mahendroo, M.; Reese, J.; Herington, J.L.; et al. Mouse models of preterm birth: Suggested assessment and reporting guidelines. Biol. Reprod. 2018, 99, 922–937. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.; Morales-Prieto, D.M.; Pastuschek, J.; Fröhlich, K. Only humans have human placentas: Molecular differences between mice and humans. J. Reprod. Immunol. 2015, 108, 65–71. [Google Scholar] [CrossRef]
- Yamane, T. Mouse yolk sac hematopoiesis. Front. Cell Dev. Biol. 2018, 6, 80. [Google Scholar] [CrossRef] [PubMed]
- Marshall, S.A.; Hannan, N.J.; Jelinic, M.; Nguyen, T.P.H.; Girling, J.E.; Parry, L.J. Animal models of preeclampsia: Translational failings and why. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 314, R499–R508. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.A. The use and misuse of animal analog models of human pregnancy disorders. J. Reprod. Immunol. 2014, 103, 1–8. [Google Scholar] [CrossRef]


{kind=link}
| Infection | Causative Agent(s) | Risk Factors | Effects on Offspring | Treatment | References |
|---|---|---|---|---|---|
| Group B Strep | Streptococcus agalactiae | History of GBS colonization in a previous pregnancy, nulliparity | Preterm birth, stillbirth, bacteremia, EOS or LOS, meningitis, death | Oral antibiotics (penicillin G), IV antibiotics during labor Newborns: supportive therapy, antibiotics | [76,77,78,79,80,81,82] |
| Listeriosis | Listeria monocytogenes | Ingestion of high-risk foods during pregnancy, including unpasteurized dairy, prepared deli meats and salads, hot dogs, and raw vegetables | Preterm birth, stillbirth, miscarriages, meningitis, sepsis, and newborn death | Penicillin, ampicillin, or amoxicillin | [83,84,85] |
| UTI | Escherichia coli, Klebsiella pneumoniae, other gram negative rods | Sexual activity, history of UTIs, bacterial vaginosis | If untreated: increased risk of preterm birth, low birth weight, and perinatal death | Nitrofurantoin, sulfisoxazole (not for use near term), cephalexin, or fosfomycin | [86,87,88] |
| Bacterial Vaginosis | Increased Gardnerella vaginalis and Mycoplasma hominis, decreased Lactobacillus | Multiple sex partners, douching, use of scented soap | Preterm labor, low birth weight, risk of miscarriage from chorioamnionitis | Metronidazole, clindamycin, or erythromycin | [89,90,91] |
| Chlamydia & Gonorrhea | Chlamydia: Chlamydia trachomatis Gonorrhea: Neisseria gonorrhoeae | Multiple sex partners, previous STI, inconsistent use of barrier protection | Preterm birth, low birth weight, conjunctivitis that may lead to blindness | Newborns: topical ocular prophylaxis with erythromycin | [92,93,94,95] |
| Syphilis | Treponema pallidum | Multiple sex partners, previous STI | Preterm birth, stillbirth, low birth weight | Penicillin G | [96,97,98] |
| CMV | Human cytomegalovirus | Primary infection during pregnancy | Hearing and/or vision loss, intellectual disabilities, microcephaly, hepatosplenomegaly, seizures, perinatal death | Supportive therapy: antiviral therapies, such as ganciclovir or valaciclovir, may be contraindicated during pregnancy but may be used to treat newborns | [99,100,101,102,103] |
| HBV | Hepatitis B virus | Contact with infected bodily fluids | Chronic liver disease or liver cancer | Newborns: HBV vaccine and Hep B immunoglobulin | [104,105] |
| RV | Rubella virus | Close contact with an infected individual | Ocular defects, auditory problems, cardiovascular defects, microcephaly, meningitis, encephalitis, neurodevelopmental delays; highest risk during first trimester infection | Supportive therapy; pregnancy counseling | [106,107] |
| ZIKV | Zika virus | Mosquito bites in endemic area | Miscarriage, stillbirth, microcephaly, lissencephaly, brain calcifications, enlarged ventricles, collapsing brain, and asymmetrical brain | Supportive therapy; close monitoring for fetal abnormalities | [108,109,110] |
| HSV | Herpes simplex virus | Genital herpes, early age of coitarche, more than one lifetime partner, previous genital chlamydia | Little effect from latent infection; risk of herpes simplex encephalitis, seizures, perinatal death from primary infection acquired close to delivery | Caesarian delivery, IV acyclovir prophylaxis Newborns: acyclovir if third-trimester primary infection and Caesarian delivery not performed | [111,112,113,114] |
| HIV | Human immunodeficiency virus | Inconsistent use of barrier protection, multiple sex partners, IV drug use, limited prenatal care | Risk of developing AIDS, cardiovascular and liver disease | Cesarean delivery, bottle feeding Newborns: Prophylactic antiretroviral therapy (zidovudine) | [115,116,117,118] |
| Toxoplasmosis | Toxoplasma gondii | Ingestion of undercooked meat, contact with feces of felines | Increased risk of severe disease if acquired during first trimester; Miscarriage, stillbirth, visual impairment, intellectual disability | Spiramycin for fetal prophylaxis; pyrimethamine, sulfadiazine, or folic acid for confirmed fetal infection | [119,120,121] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amir, M.; Brown, J.A.; Rager, S.L.; Sanidad, K.Z.; Ananthanarayanan, A.; Zeng, M.Y. Maternal Microbiome and Infections in Pregnancy. Microorganisms 2020, 8, 1996. https://doi.org/10.3390/microorganisms8121996
Amir M, Brown JA, Rager SL, Sanidad KZ, Ananthanarayanan A, Zeng MY. Maternal Microbiome and Infections in Pregnancy. Microorganisms. 2020; 8(12):1996. https://doi.org/10.3390/microorganisms8121996
Chicago/Turabian StyleAmir, Mohammed, Julia A. Brown, Stephanie L. Rager, Katherine Z. Sanidad, Aparna Ananthanarayanan, and Melody Y. Zeng. 2020. "Maternal Microbiome and Infections in Pregnancy" Microorganisms 8, no. 12: 1996. https://doi.org/10.3390/microorganisms8121996
APA StyleAmir, M., Brown, J. A., Rager, S. L., Sanidad, K. Z., Ananthanarayanan, A., & Zeng, M. Y. (2020). Maternal Microbiome and Infections in Pregnancy. Microorganisms, 8(12), 1996. https://doi.org/10.3390/microorganisms8121996