Improvement of Saccharification and Delignification Efficiency of Trichoderma reesei Rut-C30 by Genetic Bioengineering

 ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Microbial Strains and Growth Conditions
2.2. Construction of Cloning Vector
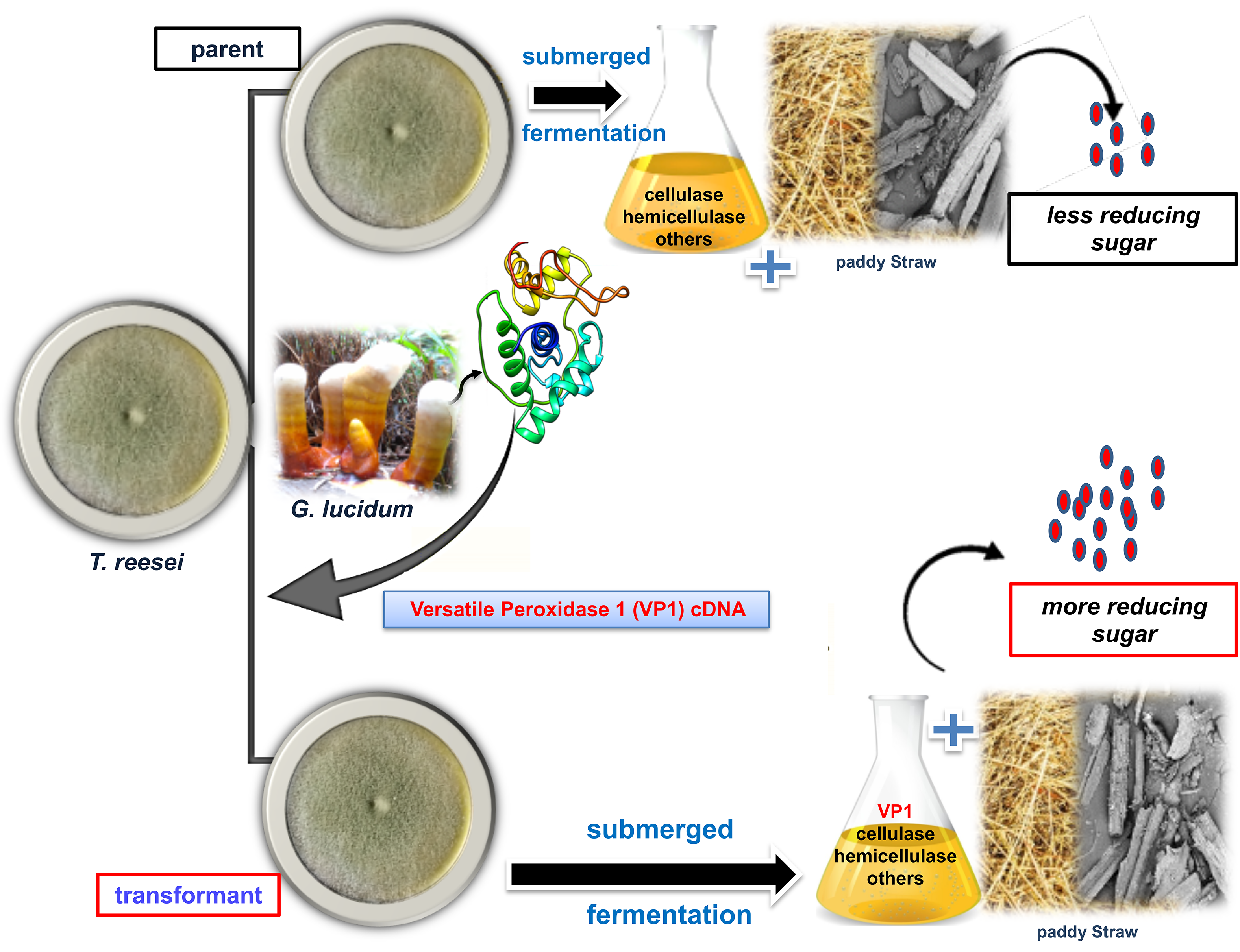
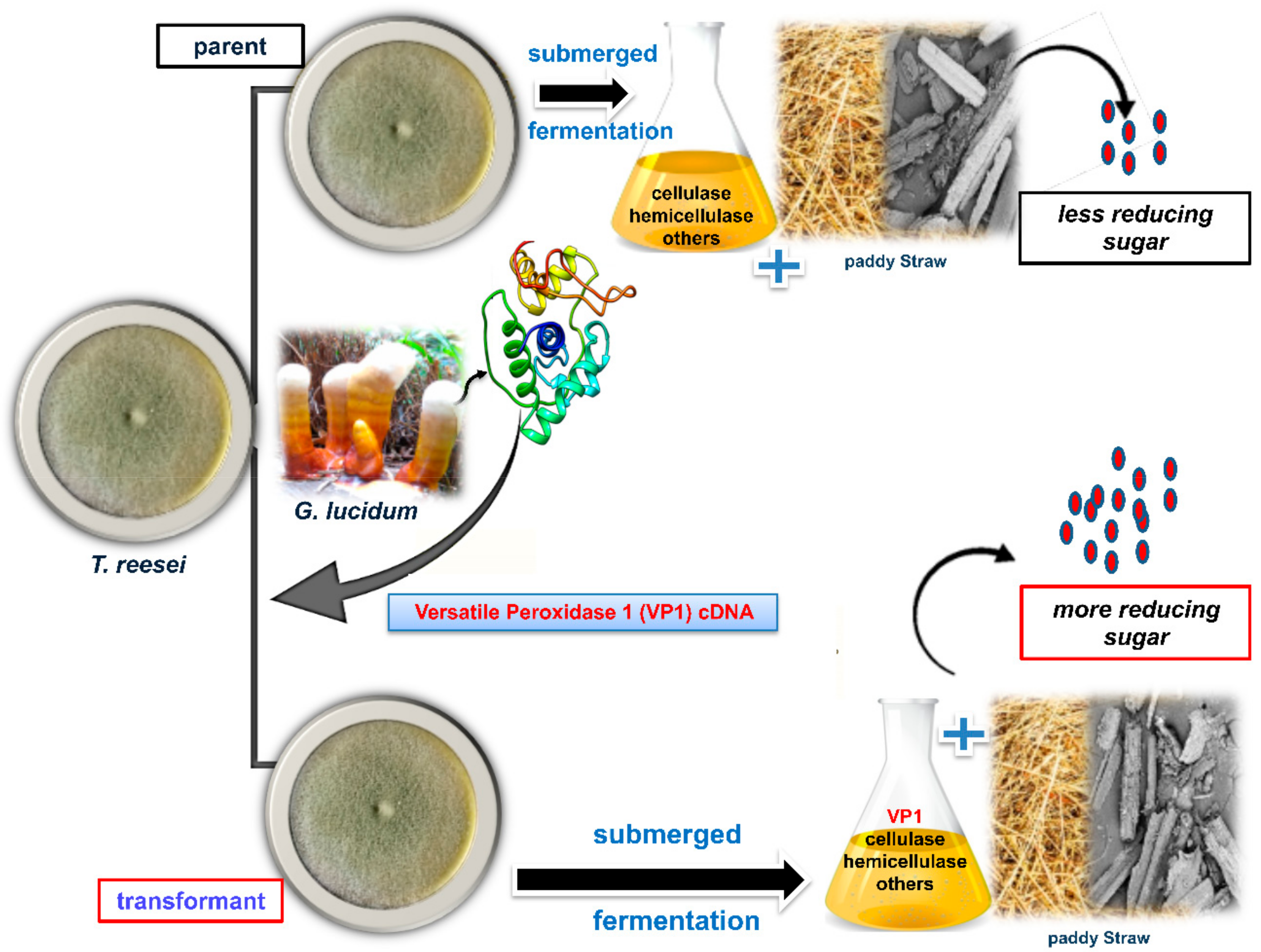
2.3. Transformation of vp1 into T. reesei Rut-C30
2.4. Enzyme Activity of rVP1 and Protein Assay
2.5. Delignification and Saccharification of Paddy Straw by Transformants and Parent T. reesei Rut-C30
2.6. Scanning Electron Microscopy
2.7. Statistical Analysis
3. Results
3.1. Construction and Transformation of Expression Vector Containing vp1 into T. reesei Rut-C30
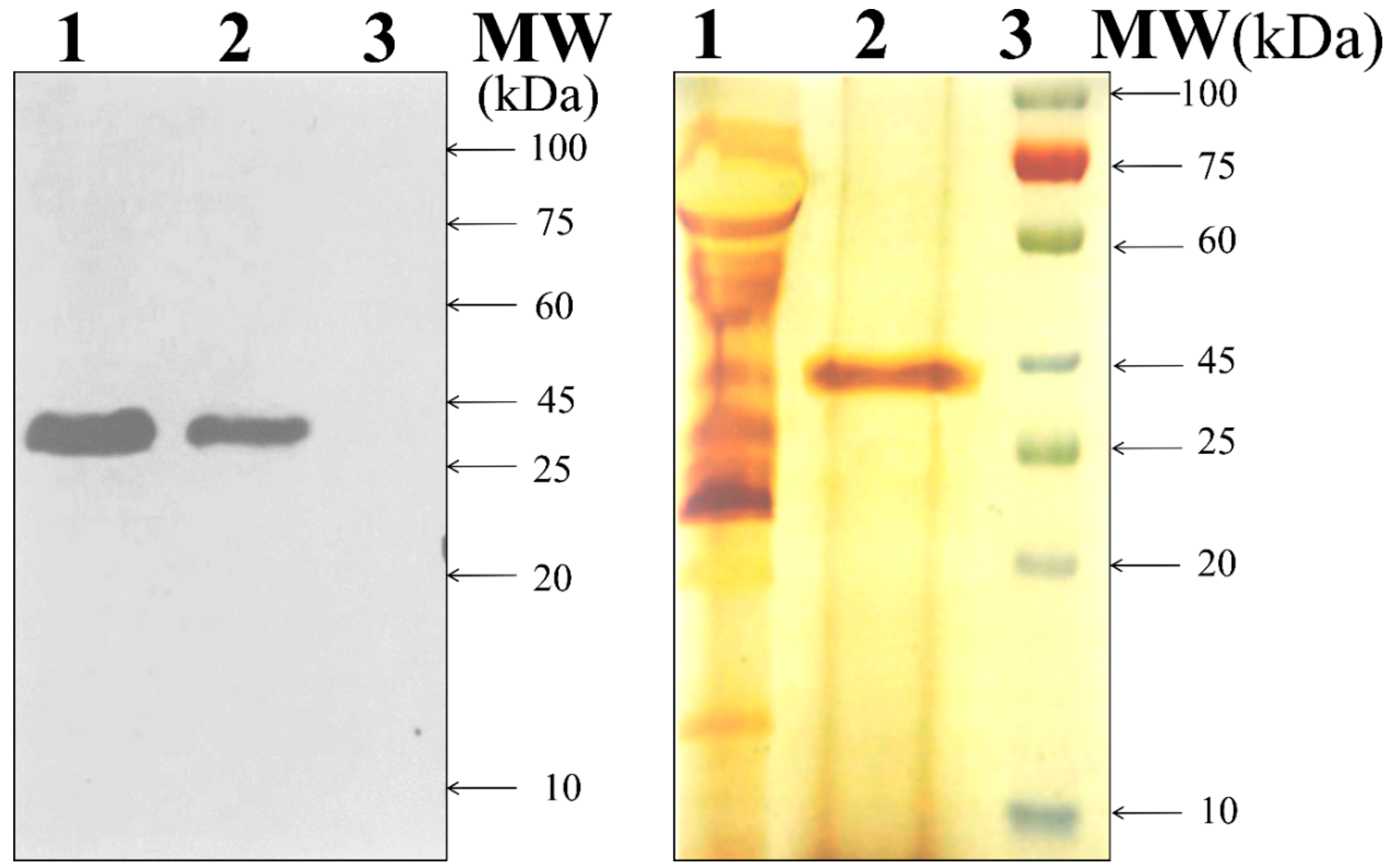
3.2. Enzyme and Protein Production by Transformants and Parent Strain T. reesei Rut-C30
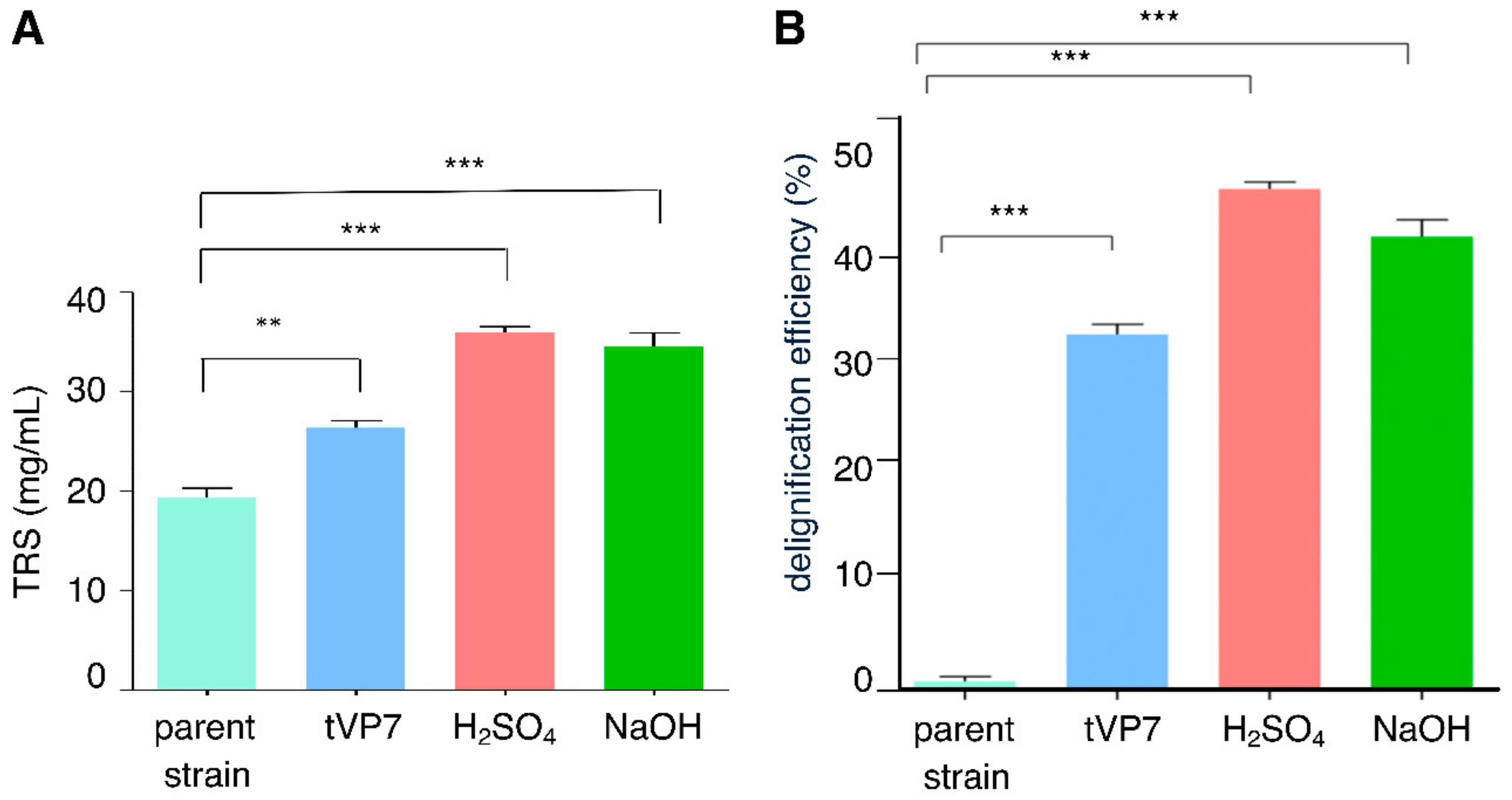
3.3. Delignification and Saccharification Efficiency of Transformed T. reesei Rut-C30
3.4. Scanning Electron Microscopic Observation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gupta, A.; Verma, J.P. Sustainable bio-ethanol production from agro-residues: A review. Renew. Sust. Energ. Rev. 2015, 41, 550–567. [Google Scholar] [CrossRef]
- Siddique, A.; Gul, A.; Irfan, M.; Nadeem, M.; Syed, Q. Comparison of different pretreatment methods for efficient conversion of bagasse into ethanol. Biofuels 2017, 8, 135–141. [Google Scholar] [CrossRef]
- Saini, J.K.; Saini, R.; Tewari, L. Lignocellulosic agriculture wastes as biomass feedstocks for second-generation bioethanol production: Concepts and recent developments. 3 Biotech. 2015, 5, 337–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D. Physico-chemical conversion of lignocellulose: Inhibitor effects and detoxification strategies: A mini review. Molecules 2018, 23, 309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pei, Y.; Li, Y.; Zhang, Y.; Yu, C.; Fu, T.; Zou, J.; Tu, Y.; Peng, L.; Chen, P. G-lignin and hemicellulosic monosaccharides distinctively affect biomass digestibility in rapeseed. Bioresour. Technol. 2016, 203, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.Y.; Cai, C.M.; Kumar, R.; Wyman, C.E. Overcoming factors limiting high-solids fermentation of lignocellulosic biomass to ethanol. Proc. Natl. Acad. Sci. USA. 2017, 114, 11673–11678. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Fan, C.; Hu, H.; Li, Y.; Sun, D.; Wang, Y.; Peng, L. Genetic modification of plant cell walls to enhance biomass yield and biofuel production in bioenergy crops. Biotechnol. Adv. 2016, 34, 997–1017. [Google Scholar] [CrossRef]
- Chandra, R.P.; Bura, R.; Mabee, W.E.; Berlin, A.; Pan, X.; Saddler, J.N. Substrate pretreatment: The key to effective enzymatic hydrolysis of lignocellulosics? Adv. Biochem. Eng. Biotechnol. 2007, 108, 67–93. [Google Scholar]
- Mosier, N.; Wyman, C.; Dale, B.; Elander, R.; Lee, Y.Y.; Holtzapple, M.; Ladisch, M. Features of promising technologies for pretreatment of lignocellulosic biomass. Bioresour. Technol. 2005, 96, 673–686. [Google Scholar] [CrossRef]
- Pienkos, P.T.; Zhang, M. Role of pretreatment and conditioning processes on toxicity of lignocellulosic biomass hydrolysates. Cellulose 2009, 16, 743–762. [Google Scholar] [CrossRef]
- Janusz, G.; Pawlik, A.; Sulej, J.; Swiderska-Burek, U.; Jarosz-Wilkolazka, A.; Paszczynski, A. Lignin degradation: Microorganisms, enzymes involved, genomes analysis and evolution. FEMS Microbiol. Rev. 2017, 41, 941–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammel, K.E.; Cullen, D. Role of fungal peroxidases in biological ligninolysis. Curr. Opin. Plant. Biol. 2008, 11, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Knezevic, A.; Stajic, M.; Jovanovic, V.M.; Kovacevic, V.; Cilerdzic, J.; Milovanovic, I.; Vukojevic, J. Induction of wheat straw delignification by Trametes species. Sci. Rep. 2016, 6, 26529. [Google Scholar] [CrossRef] [PubMed]
- Manavalan, T.; Manavalan, A.; Heese, K. Characterization of lignocellulolytic enzymes from white-rot fungi. Curr. Microbiol. 2015, 70, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Pollegioni, L.; Tonin, F.; Rosini, E. Lignin-degrading enzymes. FEBS J. 2015, 282, 1190–1213. [Google Scholar] [CrossRef] [PubMed]
- Knop, D.; Yarden, O.; Hadar, Y. The ligninolytic peroxidases in the genus Pleurotus: Divergence in activities, expression, and potential applications. Appl. Microbiol. Biotechnol. 2015, 99, 1025–1038. [Google Scholar] [CrossRef]
- Rodríguez, E.; Nuero, O.; Guillén, F.; Martínez, A.T.; Martínez, M.J. Degradation of phenolic and non-phenolic aromatic pollutants by four Pleurotus species: The role of laccase and versatile peroxidase. Soil Biol. Biochem. 2004, 36, 909–916. [Google Scholar] [CrossRef]
- Kong, W.; Fu, X.; Wang, L.; Alhujaily, A.; Zhang, J.; Ma, F.; Zhang, X.; Yu, H. A novel and efficient fungal delignification strategy based on versatile peroxidase for lignocellulose bioconversion. Biotechnol. Biofuels 2017, 10, 218. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhang, S.; Shi, Q.; Wang, L.; Kong, W.; Yu, H.; Ma, F. Highly efficient oxidation of synthetic and natural lignin-related compounds by Physisporinus vitreus versatile peroxidase. Int. Biodeterior. Biodegradation 2019, 136, 41–48. [Google Scholar] [CrossRef]
- Sun, A.; Peterson, R.; Te’o, J.; Nevalainen, H. Expression of the mammalian peptide hormone obestatin in Trichoderma reesei. Nat. Biotechnol. 2016, 33, 99–106. [Google Scholar] [CrossRef]
- Rosgaard, L.; Pedersen, S.; Langston, J.; Akerhielm, D.; Cherry, J.R.; Meyer, A.S. Evaluation of minimal Trichoderma reesei cellulase mixtures on differently pretreated Barley straw substrates. Biotechnol. Prog. 2007, 23, 1270–1276. [Google Scholar] [CrossRef] [PubMed]
- Kubicek, C.P.; Mikus, M.; Schuster, A.; Schmoll, M.; Seiboth, B. Metabolic engineering strategies for the improvement of cellulase production by Hypocrea jecorina. Biotechnol. Biofuels 2009, 2, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahamed, A.; Vermette, P. Enhanced enzyme production from mixed cultures of Trichoderma reesei RUT-C30 and Aspergillus niger LMA grown as fed batch in a stirred tank bioreactor. Biochem. Eng. J. 2008, 42, 41–46. [Google Scholar] [CrossRef]
- Saloheimo, M.; Niku-Paavola, M.L.; Knowles, J.K. Isolation and structural analysis of the laccase gene from the lignin-degrading fungus Phlebia radiata. J. Gen. Microbiol. 1991, 137, 1537–1544. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Qin, L.; Tao, Y.; Huang, J.; Dong, Z. [Overexpression and characterization of a laccase gene from Pleurotus ostreatus in Trichoderma reesei]. Wei Sheng Wu Xue Bao 2012, 52, 850–856. [Google Scholar]
- Zhao, J.; Zeng, S.; Xia, Y.; Xia, L. Expression of a thermotolerant laccase from Pycnoporus sanguineus in Trichoderma reesei and its application in the degradation of bisphenol A. J. Biosci. Bioeng. 2018, 125, 371–376. [Google Scholar] [CrossRef]
- Singh, A.; Taylor, L.E.; Vander Wall, T.A.; Linger, J.; Himmel, M.E.; Podkaminer, K.; Adney, W.S.; Decker, S.R. Heterologous protein expression in Hypocrea jecorina: A historical perspective and new developments. Biotechnol. Adv. 2015, 33, 142–154. [Google Scholar] [CrossRef] [Green Version]
- Manavalan, T.; Manavalan, A.; Thangavelu, K.P.; Heese, K. Characterization of optimized production, purification and application of laccase from Ganoderma lucidum. Biochem. Eng. J. 2013, 70, 106–114. [Google Scholar] [CrossRef]
- Lazo, G.R.; Stein, P.A.; Ludwig, R.A. A DNA transformation-competent Arabidopsis genomic library in Agrobacterium. Nat. Biotechnol. 1991, 9, 963–967. [Google Scholar] [CrossRef]
- Moller, E.M.; Bahnweg, G.; Sandermann, H.; Geiger, H.H. A simple and efficient protocol for isolation of high molecular weight DNA from filamentous fungi, fruit bodies, and infected plant tissues. Nucleic Acids Res. 1992, 20, 6115–6116. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Zhang, J.; Zou, G.; Wang, C.; Zhou, Z. Improvement of cellulase activity in Trichoderma reesei by heterologous expression of a beta-glucosidase gene from Penicillium decumbens. Enzym. Microb. Technol. 2011, 49, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Hou, Q.; Li, W.; Ju, M.; Liu, L.; Chen, Y.; Yang, Q.; Wang, J. Separation of polysaccharides from rice husk and wheat bran using solvent system consisting of BMIMOAc and DMI. Carbohydr. Polym. 2015, 133, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Zhang, W.; Ren, S.; Liu, F.; Zhao, C.; Liao, H.; Xu, Z.; Huang, J.; Li, Q.; Tu, Y.; et al. Hemicelluloses negatively affect lignocellulose crystallinity for high biomass digestibility under NaOH and H2SO4 pretreatments in Miscanthus. Biotechnol. Biofuels. 2012, 5, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Blum, H.; Beier, H.; Gross, H.J. Improved silver staining of plant proteins, RNA and DNA in polyacrylamide gels. Electrophoresis 1987, 8, 93–99. [Google Scholar] [CrossRef]
- Bischof, R.H.; Ramoni, J.; Seiboth, B. Cellulases and beyond: The first 70 years of the enzyme producer Trichoderma reesei. Microb. Cell Factories 2016, 15, 106. [Google Scholar] [CrossRef] [Green Version]
- Gruber, F.; Visser, J.; Kubicek, C.P.; de Graaff, L.H. The development of a heterologous transformation system for the cellulolytic fungus Trichoderma reesei based on a pyrG-negative mutant strain. Curr. Genet. 1990, 18, 71–76. [Google Scholar] [CrossRef]
- Penttila, M.; Nevalainen, H.; Ratto, M.; Salminen, E.; Knowles, J. A versatile transformation system for the cellulolytic filamentous fungus Trichoderma reesei. Gene 1987, 61, 155–164. [Google Scholar] [CrossRef]
- Nevalainen, H.; Peterson, R. Making recombinant proteins in filamentous fungi- are we expecting too much? Front. Microbiol. 2014, 5, 75. [Google Scholar]
- Zhong, Y.; Liu, X.; Xiao, P.; Wei, S.; Wang, T. Expression and secretion of the human erythropoietin using an optimized cbh1 promoter and the native CBH I signal sequence in the industrial fungus Trichoderma reesei. Appl. Biochem. Biotechnol. 2011, 165, 1169–1177. [Google Scholar] [CrossRef]
- Rantasalo, A.; Vitikainen, M.; Paasikallio, T.; Jantti, J.; Landowski, C.P.; Mojzita, D. Novel genetic tools that enable highly pure protein production in Trichoderma reesei. Sci. Rep. 2019, 9, 5032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Wang, J.; Wang, S.; Xing, M.; Yu, S.; Liu, G. Achieving efficient protein expression in Trichoderma reesei by using strong constitutive promoters. Microb. Cell Factories 2012, 11, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Sun, X.; Xue, X.; Luo, H.; Yao, B.; Xie, X.; Su, X. Overexpressing key component genes of the secretion pathway for enhanced secretion of an Aspergillus niger glucose oxidase in Trichoderma reesei. Enzyme Microb. Technol. 2017, 106, 83–87. [Google Scholar] [CrossRef] [PubMed]
- De Souza, P.M.; Bittencourt, M.L.; Caprara, C.C.; de Freitas, M.; de Almeida, R.P.; Silveira, D.; Fonseca, Y.M.; Ferreira Filho, E.X.; Pessoa Junior, A.; Magalhaes, P.O. A biotechnology perspective of fungal proteases. Braz. J. Microbiol. 2015, 46, 337–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu-Chau, T.A.; Ruiz-Duenas, F.J.; Camarero, S.; Feijoo, G.; Martinez, M.J.; Lema, J.M.; Martinez, A.T. Effect of pH on the stability of Pleurotus eryngii versatile peroxidase during heterologous production in Emericella nidulans. Bioprocess. Biosyst. Eng. 2004, 26, 287–293. [Google Scholar] [CrossRef]
- Haakana, H.; Miettinen-Oinonen, A.; Joutsjoki, V.; Mäntylä, A.; Suominen, P.; Vehmaanperä, J. Cloning of cellulase genes from Melanocarpus albomyces and their efficient expression in Trichoderma reesei. Enzyme Microb. Technol. 2004, 34, 159–167. [Google Scholar] [CrossRef]
- Nevalainen, K.M.; Te’o, V.S.; Bergquist, P.L. Heterologous protein expression in filamentous fungi. Trends Biotechnol. 2005, 23, 468–474. [Google Scholar] [CrossRef]
- Manavalan, T.; Liu, R.; Zhou, Z.; Zou, G. Optimization of acetyl xylan esterase gene expression in Trichoderma reesei and its application to improve the saccharification efficiency on different biomasses. Process. Biochem. 2017, 58, 160–166. [Google Scholar] [CrossRef]
- Meyer, V. Genetic engineering of filamentous fungi—Progress, obstacles and future trends. Biotechnol. Adv. 2008, 26, 177–185. [Google Scholar] [CrossRef]
- Zhang, X.; Xia, L. Expression of Talaromyces thermophilus lipase gene in Trichoderma reesei by homologous recombination at the cbh1 locus. J. Ind. Microbiol. Biotechnol. 2017, 44, 377–385. [Google Scholar] [CrossRef]
- Garcia-Ruiz, E.; Gonzalez-Perez, D.; Ruiz-Duenas, F.J.; Martinez, A.T.; Alcalde, M. Directed evolution of a temperature-, peroxide- and alkaline pH-tolerant versatile peroxidase. Biochem. J. 2012, 441, 487–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schüttmann, I.; Bouws, H.; Szweda, R.T.; Suckow, M.; Czermak, P.; Zorn, H. Induction, characterization, and heterologous expression of a carotenoid degrading versatile peroxidase from Pleurotus sapidus. J. Mol. Catal. B Enzym. 2014, 103, 79–84. [Google Scholar] [CrossRef]
- Ruiz-Duenas, F.J.; Martinez, M.J.; Martinez, A.T. Molecular characterization of a novel peroxidase isolated from the ligninolytic fungus Pleurotus eryngii. Mol. Microbiol. 1999, 31, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Boada, M.; Doyle, W.A.; Ruiz-Dueñas, F.J.; Martínez, M.J.; Martínez, A.T.; Smith, A.T. Expression of Pleurotus eryngii versatile peroxidase in Escherichia coli and optimisation of in vitro folding. Enzym. Microb. Technol. 2002, 30, 518–524. [Google Scholar] [CrossRef]
- Tsukihara, T.; Honda, Y.; Sakai, R.; Watanabe, T.; Watanabe, T. Exclusive overproduction of recombinant versatile peroxidase MnP2 by genetically modified white rot fungus, Pleurotus ostreatus. J. Biotechnol. 2006, 126, 431–439. [Google Scholar] [CrossRef]
- Tsukihara, T.; Honda, Y.; Watanabe, T.; Watanabe, T. Molecular breeding of white rot fungus Pleurotus ostreatus by homologous expression of its versatile peroxidase MnP2. Appl. Microbiol. Biotechnol. 2006, 71, 114–120. [Google Scholar] [CrossRef]
- Ruiz-Dueñas, F.J.; Aguilar, A.; Martínez, M.J.; Zorn, H.; Martínez, Á.T. Gene cloning, heterologous expression, in vitro reconstitution and catalytic properties of a versatile peroxidase. Biocatal. Biotransfor. 2007, 25, 276–285. [Google Scholar] [CrossRef]
- Mohorcic, M.; Bencina, M.; Friedrich, J.; Jerala, R. Expression of soluble versatile peroxidase of Bjerkandera adusta in Escherichia coli. Bioresour. Technol. 2009, 100, 851–858. [Google Scholar] [CrossRef]
- Eibes, G.M.; Lu-Chau, T.A.; Ruiz-Dueñas, F.J.; Feijoo, G.; Martinez, M.J.; Martinez, A.T.; Lema, J.M. Effect of culture temperature on the heterologous expression of Pleurotus eryngii versatile peroxidase in Aspergillus hosts. Bioprocess. Biosyst. Eng. 2009, 32, 129–134. [Google Scholar] [CrossRef] [Green Version]
- Marques, G.; Gamelas, J.A.F.; Ruiz-Dueñas, F.J.; del Rio, J.C.; Evtuguin, D.V.; Martínez, A.T.; Gutiérrez, A. Delignification of eucalypt kraft pulp with manganese-substituted polyoxometalate assisted by fungal versatile peroxidase. Bioresour. Technol. 2010, 101, 5935–5940. [Google Scholar] [CrossRef]
- Bao, X.; Liu, A.; Lu, X.; Li, J.J. Direct over-expression, characterization and H2O2 stability study of active Pleurotus eryngii versatile peroxidase in Escherichia coli. Biotechnol. Lett. 2012, 34, 1537–1543. [Google Scholar] [CrossRef] [PubMed]
- Salame, T.M.; Knop, D.; Tal, D.; Levinson, D.; Yarden, O.; Hadar, Y. Predominance of a versatile-peroxidase-encoding gene, mnp4, as demonstrated by gene replacement via a gene targeting system for Pleurotus ostreatus. Appl. Environ. Microbiol. 2012, 78, 5341–5352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coconi-Linares, N.; Magana-Ortiz, D.; Guzman-Ortiz, D.A.; Fernandez, F.; Loske, A.M.; Gomez-Lim, M.A. High-yield production of manganese peroxidase, lignin peroxidase, and versatile peroxidase in Phanerochaete chrysosporium. Appl. Microbiol. Biotechnol. 2014, 98, 9283–9294. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.; Huang, X.; Lu, X.; Li, J.J. Improvement of hydrogen peroxide stability of Pleurotus eryngii versatile ligninolytic peroxidase by rational protein engineering. Enzym. Microb. Technol. 2014, 54, 51–58. [Google Scholar] [CrossRef]
- Imami, A.; Riemer, S.; Schulze, M.; Amelung, F.; Gorshkov, V.; Rühl, M.; Ammenn, J.; Zorn, H. Depolymerization of lignosulfonates by submerged cultures of the basidiomycete Irpex consors and cloning of a putative versatile peroxidase. Enzyme Microb. Technol. 2015, 81, 8–15. [Google Scholar] [CrossRef]
- Sukumaran, R.K.; Surender, V.J.; Sindhu, R.; Binod, P.; Janu, K.U.; Sajna, K.V.; Rajasree, K.P.; Pandey, A. Lignocellulosic ethanol in India: Prospects, challenges and feedstock availability. Bioresour. Technol. 2010, 101, 4826–4833. [Google Scholar] [CrossRef]
- Ko, J.K.; Bak, J.S.; Jung, M.W.; Lee, H.J.; Choi, I.-G.; Kim, T.H.; Kim, K.H. Ethanol production from rice straw using optimized aqueous-ammonia soaking pretreatment and simultaneous saccharification and fermentation processes. Bioresour. Technol. 2009, 100, 4374–4380. [Google Scholar] [CrossRef]
- Alvira, P.; Tomás-Pejó, E.; Ballesteros, M.; Negro, M.J. Pretreatment technologies for an efficient bioethanol production process based on enzymatic hydrolysis: A review. Bioresour. Technol. 2010, 101, 4851–4861. [Google Scholar] [CrossRef]
- Kaur, P.P.; Arneja, J.S.; Singh, J. Enzymic hydrolysis of rice straw by crude cellulase from Trichoderma reesei. Bioresour. Technol. 1998, 66, 267–269. [Google Scholar] [CrossRef]
- Salehi, S.M.A.; Karimi, K.; Behzad, T.; Poornejad, N. Efficient conversion of rice straw to bioethanol using sodium carbonate pretreatment. Energy Fuels 2012, 26, 7354–7361. [Google Scholar] [CrossRef]
- Kshirsagar, S.D.; Waghmare, P.R.; Chandrakant Loni, P.; Patil, S.A.; Govindwar, S.P. Dilute acid pretreatment of rice straw, structural characterization and optimization of enzymatic hydrolysis conditions by response surface methodology. RSC Adv. 2015, 5, 46525–46533. [Google Scholar] [CrossRef]
- Kapoor, M.; Soam, S.; Agrawal, R.; Gupta, R.P.; Tuli, D.K.; Kumar, R. Pilot scale dilute acid pretreatment of rice straw and fermentable sugar recovery at high solid loadings. Bioresour. Technol. 2017, 224, 688–693. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Phutela, U.G. Enhancement of paddy straw digestibility and biogas production by sodium hydroxide-microwave pretreatment. Renew. Energy 2016, 92, 178–184. [Google Scholar] [CrossRef]
- Asgher, M.; Ahmad, Z.; Iqbal, H.M.N. Alkali and enzymatic delignification of sugarcane bagasse to expose cellulose polymers for saccharification and bio-ethanol production. Ind. Crops. Prod. 2013, 44, 488–495. [Google Scholar] [CrossRef]
- Masran, R.; Zanirun, Z.; Bahrin, E.K.; Ibrahim, M.F.; Lai Yee, P.; Abd-Aziz, S. Harnessing the potential of ligninolytic enzymes for lignocellulosic biomass pretreatment. Appl. Microbiol. Biotechnol. 2016, 100, 5231–5246. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Extracellular (mg/mL) | Protein VP Activity (U/L) | FPA (U/mL) | VP-Specific Activity (U/mg) |
|---|---|---|---|---|
| T. reesei Rut-C30 | 09.08 ± 0.03 | ND | 13.61 ± 0.24 | ND |
| tVP7 | 10.52 ± 0.01 | 437 ± 1.0 | 14.92 ± 0.32 | 41.53 ± 1.02 *** |
| tVP13 | 09.48 ± 0.02 | 384 ± 1.08 | 12.36 ± 0.25 | 40.50 ± 1.04 *** |
| Source | Host | Vector | Promoter | Enzyme Yield | Reference |
|---|---|---|---|---|---|
| Physisporinus vitreus | E. coli BL21(DE3) | pET-28a(+) | default | 18 mg/L | [19] |
| Pleurotus eryngii | Emerciella nidulans | - | alcA | 165 U/L | [46] |
| Pleurotus sapidus | Hansenula polymorpha RB11 | pFPMT121 | default | 1.80 ± 0.10 U/mg specific activity | [52] |
| Pleurotus eryngii | Aspergillus nidulans | palcA1 | alcA | 294.3 mg | [53] |
| Pleurotus eryngii | E. coli W3110 | pFFLAG1 | tac | - | [54] |
| Pleurotus ostreatus | Pleurotus ostreatus | pIpM2g | sdi I | 1.5 mg/L | [55] |
| Pleurotus ostreatus | Pleurotus ostreatus | pIpMc | sdi I | 370 mU/mL | [56] |
| Pleurotus eryngii | E. coli W3110 | pFFLAG1 | tac | 5.5 mg/L | [57] |
| Bjarkendra adjusta | E. coli BL21(DE3) | pET-32b(+) and pET-19b(+) | default | 322 U/L | [58] |
| Pleurotus eryngii | Aspergillus nidulans, | PAN7-1 | alcA | 466 U/L (A. nidulans); | [59] |
| Aspergillus niger | 412 U/L (A. niger) | ||||
| Pleurotus eryngii | E. coli W3110 | pFLAG-VPL2 | default | - | [60] |
| Pleurotus eryngii | E. coli BL21(DE3) | pET-28a(+) and pET-32a(+) | default | 12.5 mg/L | [61] |
| Pleurotus ostreatus | Pleurotus ostreatus | pTM1 | β-tubulin promoter | 96 mU/mg substrate | [62] |
| Pleurotus eryngii | Phanerochaete chrysosporium | Ppchph | gpd | 20 U/mL | [63] |
| Moniliophthora roreri | Pichia pastoris | pPICZaA | aox1 | 295 mg/L | [64] |
| Irpex consors | E. coli Top 10 | - | - | > 250 U/L | [65] |
| Ganoderma lucidum | T. reesei Rut-C30 | pCambia-1300 | cbh1 | 437 U/L | present study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gopalakrishnan, R.M.; Manavalan, T.; Ramesh, J.; Thangavelu, K.P.; Heese, K. Improvement of Saccharification and Delignification Efficiency of Trichoderma reesei Rut-C30 by Genetic Bioengineering. Microorganisms 2020, 8, 159. https://doi.org/10.3390/microorganisms8020159
Gopalakrishnan RM, Manavalan T, Ramesh J, Thangavelu KP, Heese K. Improvement of Saccharification and Delignification Efficiency of Trichoderma reesei Rut-C30 by Genetic Bioengineering. Microorganisms. 2020; 8(2):159. https://doi.org/10.3390/microorganisms8020159
Chicago/Turabian StyleGopalakrishnan, Raja Mohan, Tamilvendan Manavalan, Janani Ramesh, Kalaichelvan Puthupalayam Thangavelu, and Klaus Heese. 2020. "Improvement of Saccharification and Delignification Efficiency of Trichoderma reesei Rut-C30 by Genetic Bioengineering" Microorganisms 8, no. 2: 159. https://doi.org/10.3390/microorganisms8020159
APA StyleGopalakrishnan, R. M., Manavalan, T., Ramesh, J., Thangavelu, K. P., & Heese, K. (2020). Improvement of Saccharification and Delignification Efficiency of Trichoderma reesei Rut-C30 by Genetic Bioengineering. Microorganisms, 8(2), 159. https://doi.org/10.3390/microorganisms8020159