Mbov_0503 Encodes a Novel Cytoadhesin that Facilitates Mycoplasma bovis Interaction with Tight Junctions



Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Plasmids and DNA Constructions
2.3. Bacteria, Cell Lines and Culture Conditions
2.4. Transposon Mutagenesis in M. bovis and Mapping of Transposon Insertion Sites
2.5. Mycoplasma Adhesion Assay and Selection of Mutants with A Reduced Binding Capacity
2.6. Expression and Purification of rMbovP0503
2.7. Western and Colony Immunoblotting Analyses
2.8. Laser Scanning Confocal Microscopy
2.9. ELISA Binding Assays
2.10. Transwell Monolayer Translocation Assays
2.11. Bioinformatic Analysis
2.12. Statistical Analysis
3. Results
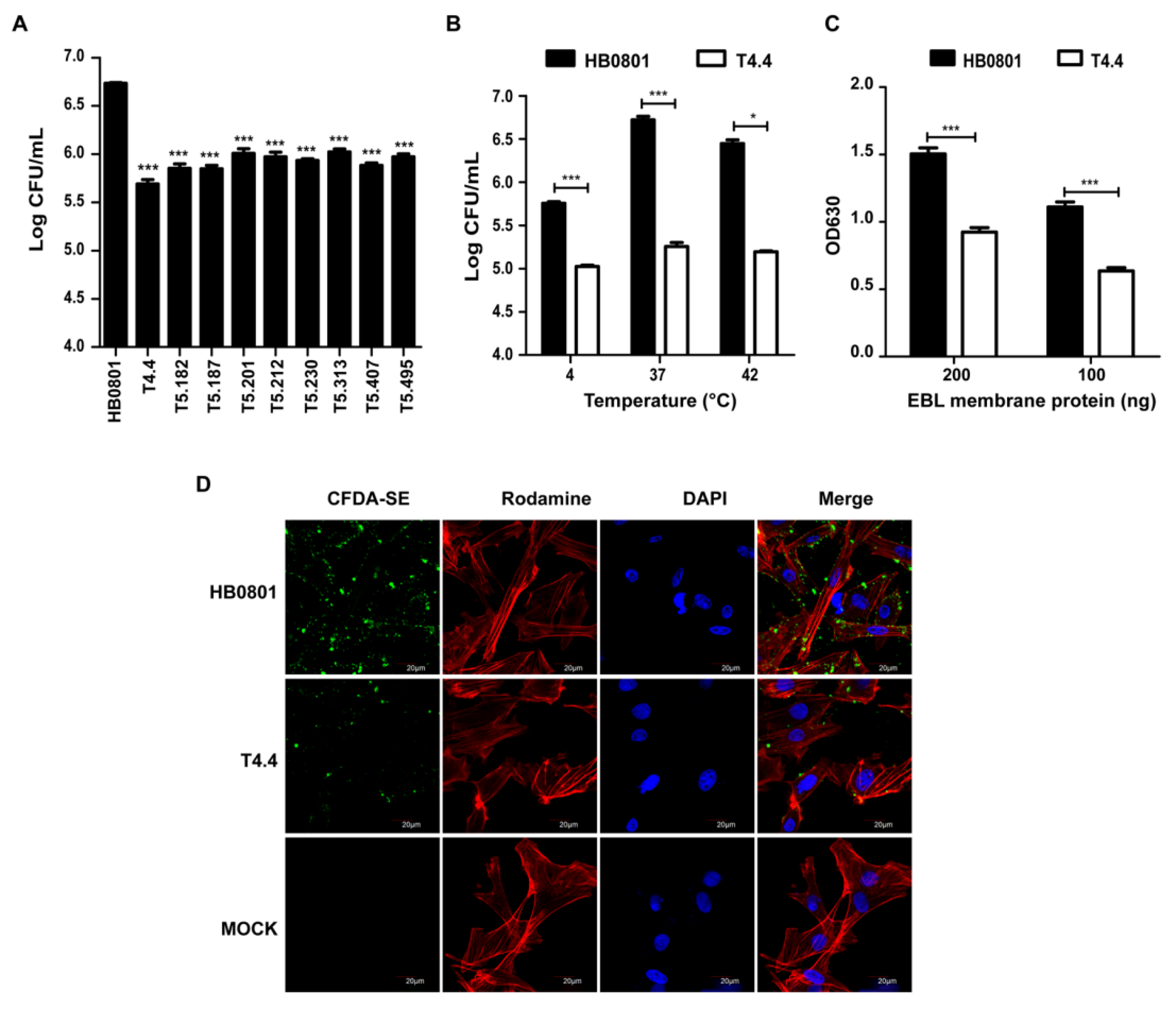
3.1. Selection of M. bovis Mutants with Reduced Binding to Host Cells
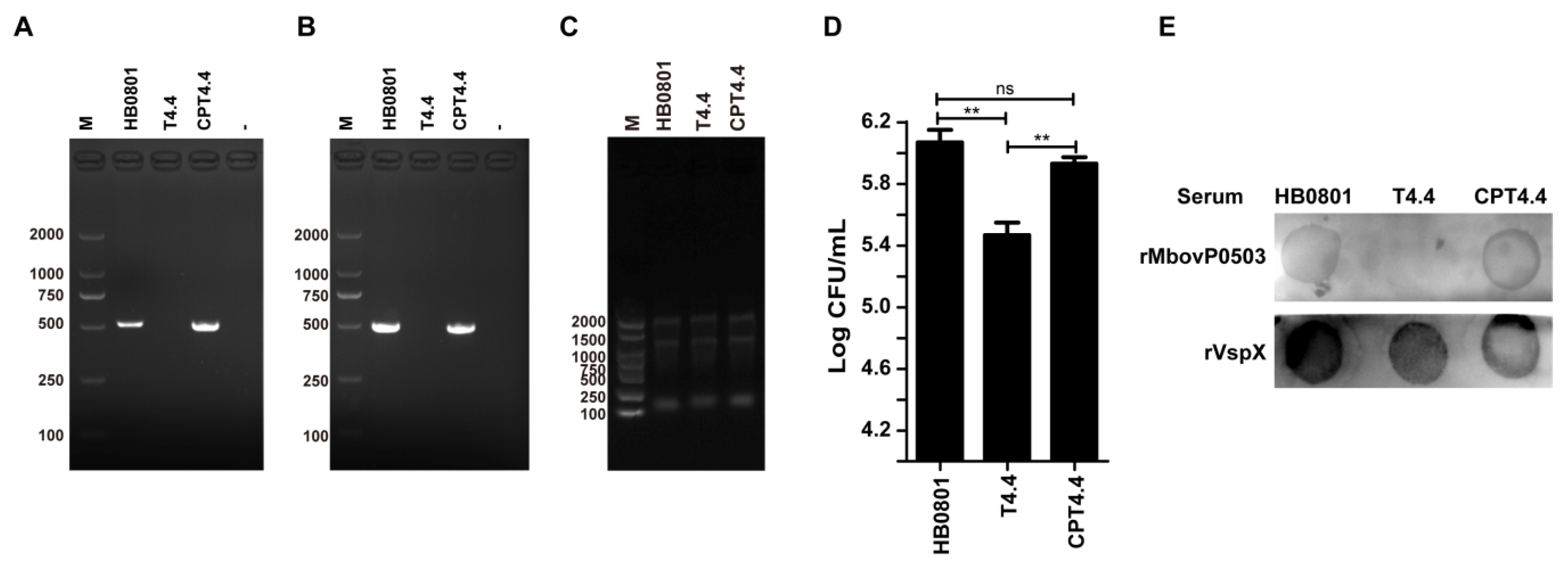
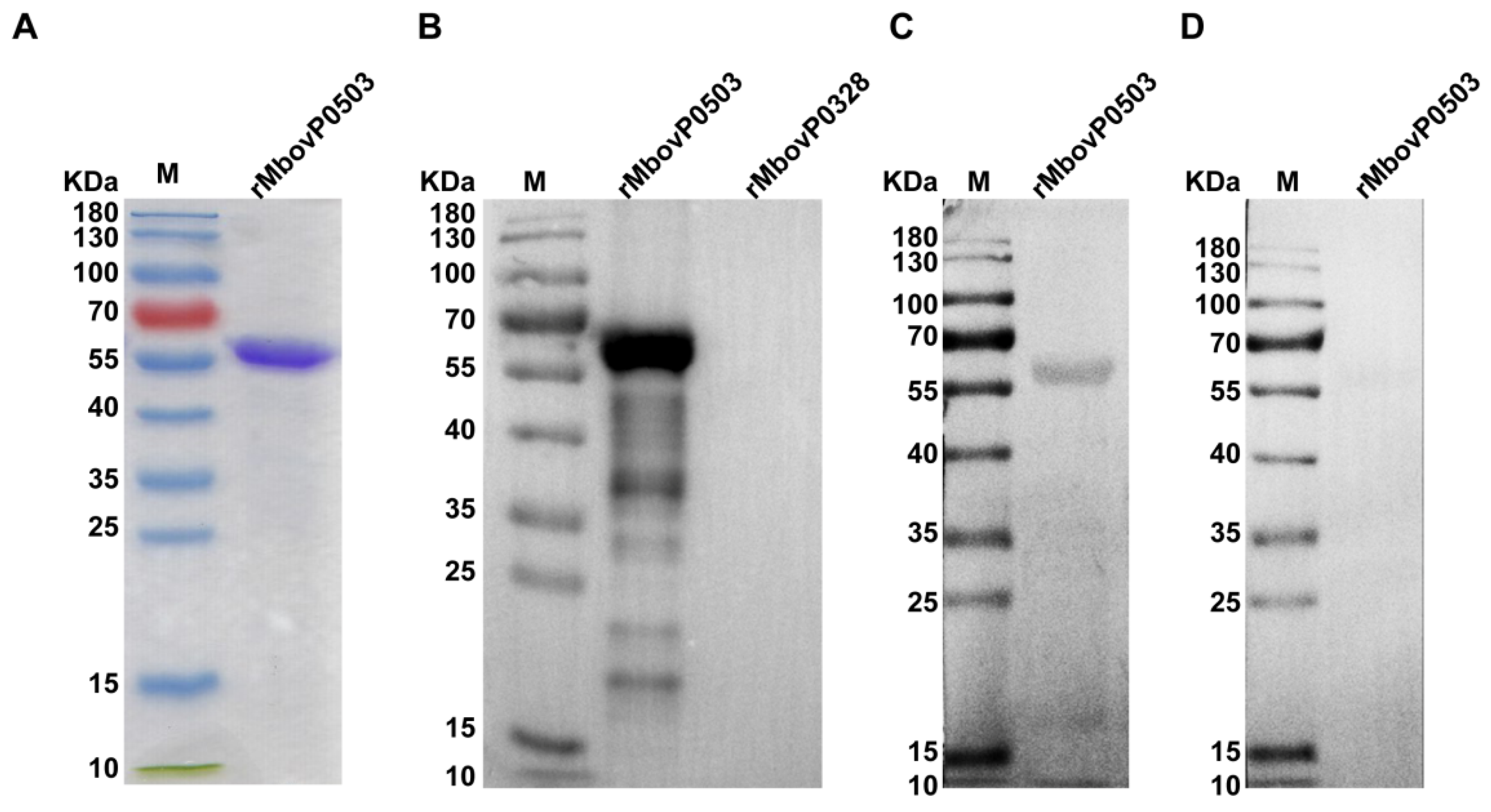
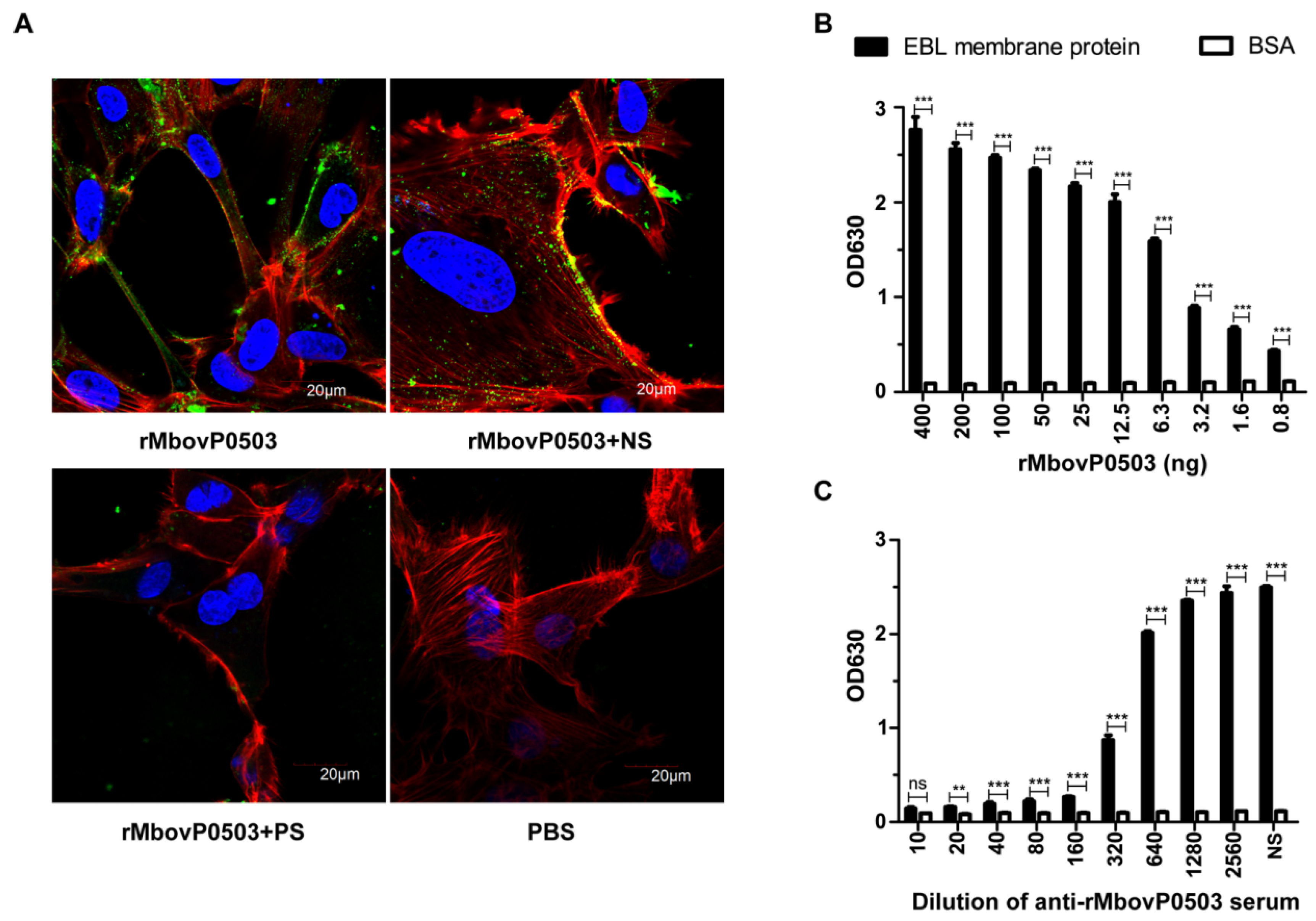
3.2. Mbov_0503 Encodes a Novel Cytoadhesin of M. bovis
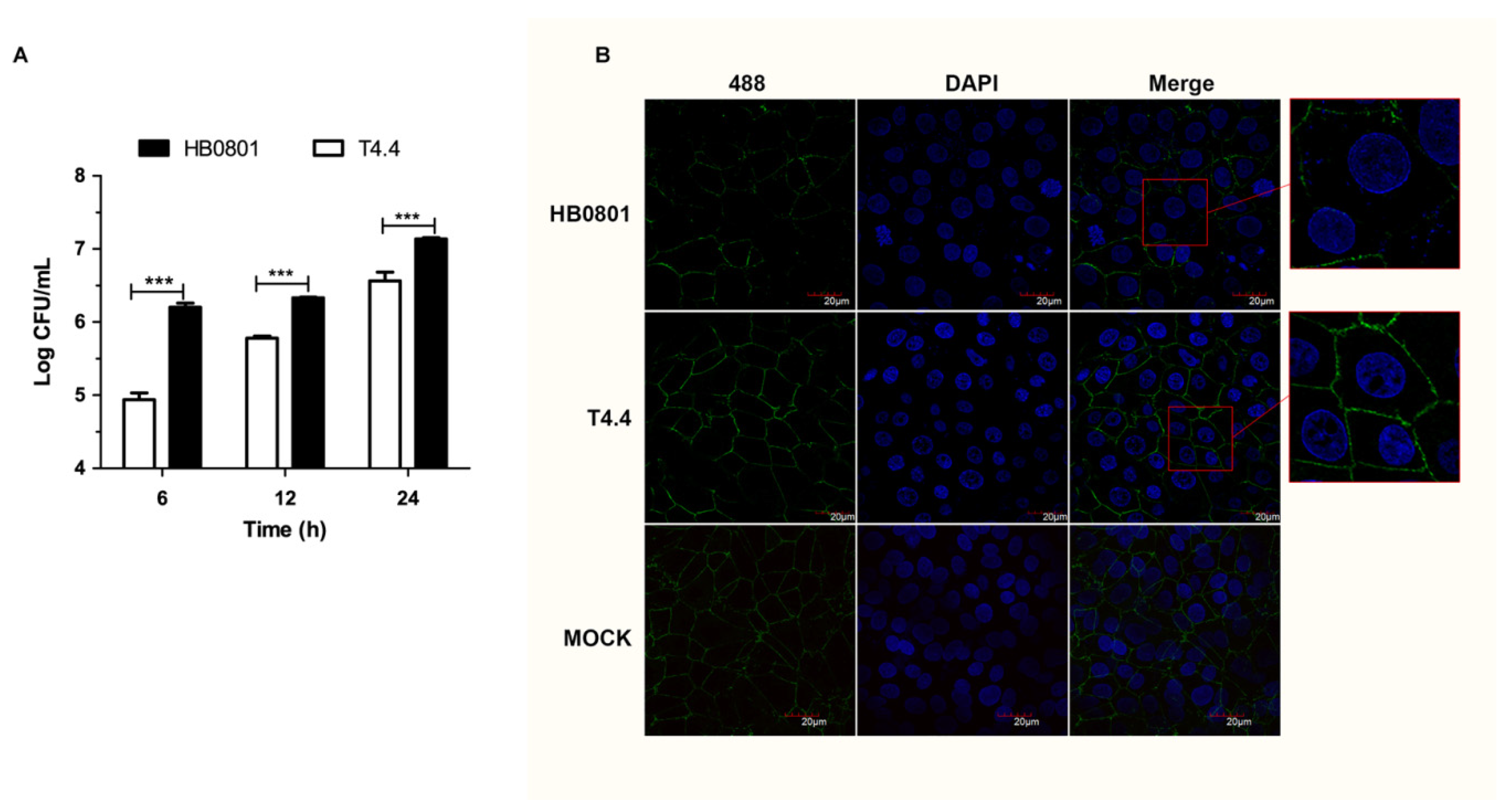
3.3. Mbov_0503 Facilitates M. bovis Interaction with Tight Junctions
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Citti, C.; Dordet-Frisoni, E.; Nouvel, L.X.; Kuo, C.H.; Baranowski, E. Horizontal Gene Transfers in Mycoplasmas (Mollicutes). Curr. Issues Mol. Biol. 2018, 29, 3–22. [Google Scholar] [CrossRef] [Green Version]
- Bischof, D.F.; Janis, C.; Vilei, E.M.; Bertoni, G.; Frey, J. Cytotoxicity of Mycoplasma mycoides subsp. mycoides small colony type to bovine epithelial cells. Infect. Immun. 2008, 76, 263–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kannan, T.R.; Baseman, J.B. ADP-ribosylating and vacuolating cytotoxin of Mycoplasma pneumoniae represents unique virulence determinant among bacterial pathogens. Proc. Natl. Acad. Sci. USA 2006, 103, 6724–6729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razin, S.; Yogev, D.; Naot, Y. Molecular biology and pathogenicity of mycoplasmas. Microbiol. Mol. Biol. Rev. 1998, 62, 1094–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aye, R.; Mwirigi, M.K.; Frey, J.; Pilo, P.; Jores, J.; Naessens, J. Cyto-adherence of Mycoplasma mycoides subsp. mycoides to bovine lung epithelial cells. BMC Vet. Res. 2015, 11, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, J.; Guo, A.; Cui, P.; Chen, Y.; Mustafa, R.; Ba, X.; Hu, C.; Bai, Z.; Chen, X.; Shi, L.; et al. Comparative geno-plasticity analysis of Mycoplasma bovis HB0801 (Chinese isolate). PLoS ONE 2012, 7, e38239. [Google Scholar] [CrossRef] [PubMed]
- Burki, S.; Frey, J.; Pilo, P. Virulence, persistence and dissemination of Mycoplasma bovis. Vet. Microbiol. 2015, 179, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Dordet-Frisoni, E.; Gillard, L.; Ba, A.; Hygonenq, M.C.; Sagne, E.; Nouvel, L.X.; Maillard, R.; Assie, S.; Guo, A.; et al. Extracellular DNA: A Nutritional Trigger of Mycoplasma bovis Cytotoxicity. Front. Microbiol. 2019, 10, 2753. [Google Scholar] [CrossRef]
- May, M.; Papazisi, L.; Gorton, T.S.; Geary, S.J. Identification of fibronectin-binding proteins in Mycoplasma gallisepticum strain R. Infect. Immun. 2006, 74, 1777–1785. [Google Scholar] [CrossRef] [Green Version]
- Szczepanek, S.M.; Majumder, S.; Sheppard, E.S.; Liao, X.; Rood, D.; Tulman, E.R.; Wyand, S.; Krause, D.C.; Silbart, L.K.; Geary, S.J. Vaccination of BALB/c mice with an avirulent Mycoplasma pneumoniae P30 mutant results in disease exacerbation upon challenge with a virulent strain. Infect. Immun. 2012, 80, 1007–1014. [Google Scholar] [CrossRef] [Green Version]
- Chaudhry, R.; Ghosh, A.; Chandolia, A. Pathogenesis of Mycoplasma pneumoniae: An update. Indian J. Med. Microbiol. 2016, 34, 7–16. [Google Scholar] [CrossRef]
- Popham, P.L.; Hahn, T.W.; Krebes, K.A.; Krause, D.C. Loss of HMW1 and HMW3 in noncytadhering mutants of Mycoplasma pneumoniae occurs post-translationally. Proc. Natl. Acad. Sci. USA 1997, 94, 13979–13984. [Google Scholar] [CrossRef] [Green Version]
- Widjaja, M.; Berry, I.J.; Pont, E.J.; Padula, M.P.; Djordjevic, S.P. P40 and P90 from Mpn142 are Targets of Multiple Processing Events on the Surface of Mycoplasma pneumoniae. Proteomes 2015, 3, 512–537. [Google Scholar] [CrossRef]
- Hopfe, M.; Dahlmanns, T.; Henrich, B. In Mycoplasma hominis the OppA-mediated cytoadhesion depends on its ATPase activity. BMC Microbiol. 2011, 11, 185. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Liu, M.; Ye, Z.; Tan, T.; Liu, X.; You, X.; Zeng, Y.; Wu, Y. Insights into the pathogenesis of Mycoplasma pneumoniae (Review). Mol. Med. Rep. 2016, 14, 4030–4036. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Zhu, H.; Wang, J.; Huang, J.; Khan, F.A.; Zhang, J.; Guo, A.; Chen, X. TrmFO, a Fibronectin-Binding Adhesin of Mycoplasma bovis. Int. J. Mol. Sci. 2017, 18, 1732. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Zhang, H.; Chen, X.; Zhu, X.; Guo, Y.; He, C.; Anwar Khan, F.; Chen, Y.; Hu, C.; Chen, H.; et al. Mycoplasma bovis NADH oxidase functions as both a NADH oxidizing and O2 reducing enzyme and an adhesin. Sci. Rep. 2017, 7, 44. [Google Scholar] [CrossRef]
- Huang, J.; Zhu, H.; Wang, J.; Guo, Y.; Zhi, Y.; Wei, H.; Li, H.; Guo, A.; Liu, D.; Chen, X. Fructose-1,6-bisphosphate aldolase is involved in Mycoplasma bovis colonization as a fibronectin-binding adhesin. Res. Vet. Sci. 2019, 124, 70–78. [Google Scholar] [CrossRef]
- Gao, X.; Bao, S.; Xing, X.; Fu, X.; Zhang, Y.; Xue, H.; Wen, F.; Wei, Y. Fructose-1,6-bisphosphate aldolase of Mycoplasma bovis is a plasminogen-binding adhesin. Microb. Pathog. 2018, 124, 230–237. [Google Scholar] [CrossRef]
- Sachse, K.; Grajetzki, C.; Rosengarten, R.; Hanel, I.; Heller, M.; Pfutzner, H. Mechanisms and factors involved in Mycoplasma bovis adhesion to host cells. Zent. Bakteriol. 1996, 284, 80–92. [Google Scholar] [CrossRef]
- Chen, X.; Huang, J.; Zhu, H.; Guo, Y.; Khan, F.A.; Menghwar, H.; Zhao, G.; Guo, A. P27 (MBOV_RS03440) is a novel fibronectin binding adhesin of Mycoplasma bovis. Int. J. Med. Microbiol. 2018, 308, 848–857. [Google Scholar] [CrossRef] [PubMed]
- Sachse, K.; Helbig, J.H.; Lysnyansky, I.; Grajetzki, C.; Muller, W.; Jacobs, E.; Yogev, D. Epitope mapping of immunogenic and adhesive structures in repetitive domains of Mycoplasma bovis variable surface lipoproteins. Infect. Immun. 2000, 68, 680–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Z.; Li, Y.; Liu, Y.; Xin, J.; Zou, X.; Sun, W. alpha-Enolase, an adhesion-related factor of Mycoplasma bovis. PLoS ONE 2012, 7, e38836. [Google Scholar] [CrossRef]
- Josi, C.; Burki, S.; Vidal, S.; Dordet-Frisoni, E.; Citti, C.; Falquet, L.; Pilo, P. Large-Scale Analysis of the Mycoplasma bovis Genome Identified Non-essential, Adhesion- and Virulence-Related Genes. Front. Microbiol. 2019, 10, 2085. [Google Scholar] [CrossRef] [PubMed]
- Rottem, S.; Yogev, D. Mycoplasma interaction with eukaryotic cells. Subcell. Biochem. 2000, 33, 199–227. [Google Scholar] [PubMed]
- Zimmerman, C.U.; Herrmann, R. Synthesis of a small, cysteine-rich, 29 amino acids long peptide in Mycoplasma pneumoniae. FEMS Microbiol. Lett. 2005, 253, 315–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baranowski, E.; Dordet-Frisoni, E.; Sagne, E.; Hygonenq, M.C.; Pretre, G.; Claverol, S.; Fernandez, L.; Nouvel, L.X.; Citti, C. The Integrative Conjugative Element (ICE) of Mycoplasma agalactiae: Key Elements Involved in Horizontal Dissemination and Influence of Coresident ICEs. MBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Dordet Frisoni, E.; Marenda, M.S.; Sagne, E.; Nouvel, L.X.; Guerillot, R.; Glaser, P.; Blanchard, A.; Tardy, F.; Sirand-Pugnet, P.; Baranowski, E.; et al. ICEA of Mycoplasma agalactiae: A new family of self-transmissible integrative elements that confers conjugative properties to the recipient strain. Mol. Microbiol. 2013, 89, 1226–1239. [Google Scholar] [CrossRef]
- Han, X.; Khan, F.A.; Zhu, X.; Zhang, R.; Mustafa, R.; Hu, C.; Chen, Y.; Chen, H.; Guo, A. Establishment of an antibody avidity test to differentiate vaccinated cattle from those naturally infected with Mycoplasma bovis. Vet. J. 2015, 203, 79–84. [Google Scholar] [CrossRef]
- Baranowski, E.; Guiral, S.; Sagne, E.; Skapski, A.; Citti, C. Critical role of dispensable genes in Mycoplasma agalactiae interaction with mammalian cells. Infect. Immun. 2010, 78, 1542–1551. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.; Schwarzwaelder, K.; Bartholomae, C.; Zaoui, K.; Ball, C.; Pilz, I.; Braun, S.; Glimm, H.; von Kalle, C. High-resolution insertion-site analysis by linear amplification-mediated PCR (LAM-PCR). Nat. Methods 2007, 4, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Han, X.; Chen, Y.; Mustafa, R.; Qi, J.; Chen, X.; Hu, C.; Chen, H.; Guo, A. Attenuated Mycoplasma bovis strains provide protection against virulent infection in calves. Vaccine 2014, 32, 3107–3114. [Google Scholar] [CrossRef]
- Zhang, Q.; Young, T.F.; Ross, R.F. Microtiter plate adherence assay and receptor analogs for Mycoplasma hyopneumoniae. Infect. Immun. 1994, 62, 1616–1622. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.M.; Jiang, Y.; Xiao-Liang, B.A.; Chang-Min, H.U.; Chen, Y.Y.; Peng, Q.J.; Chen, H.C.; Guo, A.Z. Preparation and Identification of Monoclonal Antibodies against Mycoplasma bovis. China Anim. Husb. Vet. Med. 2012, 39, 58–61. [Google Scholar]
- Everman, J.L.; Bermudez, L.E. Antibodies against invasive phenotype-specific antigens increase Mycobacterium avium subspecies paratuberculosis translocation across a polarized epithelial cell model and enhance killing by bovine macrophages. Front. Cell. Infect. Microbiol. 2015, 5, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenenbaum, T.; Papandreou, T.; Gellrich, D.; Friedrichs, U.; Seibt, A.; Adam, R.; Wewer, C.; Galla, H.J.; Schwerk, C.; Schroten, H. Polar bacterial invasion and translocation of Streptococcus suis across the blood-cerebrospinal fluid barrier in vitro. Cell. Microbiol. 2009, 11, 323–336. [Google Scholar] [CrossRef]
- Yu, N.Y.; Wagner, J.R.; Laird, M.R.; Melli, G.; Rey, S.; Lo, R.; Dao, P.; Sahinalp, S.C.; Ester, M.; Foster, L.J.; et al. PSORTb 3.0: Improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics 2010, 26, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Bendtsen, J.D.; Kiemer, L.; Fausboll, A.; Brunak, S. Non-classical protein secretion in bacteria. BMC Microbiol. 2005, 5, 58. [Google Scholar] [CrossRef] [Green Version]
- Chaudhry, R.; Varshney, A.K.; Malhotra, P. Adhesion proteins of Mycoplasma pneumoniae. Front. Biosci. 2007, 12, 690–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffery, C.J. Expression, Solubilization, and Purification of Bacterial Membrane Proteins. Curr. Protoc. Protein Sci. 2016, 83, 29. [Google Scholar] [CrossRef] [PubMed]
- Lv, D.D.; Zhang, Y.Y.; Ge, H.T.; Huang, X.H.; Wang, Y.C. Advances of the technologies in large-scale membrane proteome identification. Yi Chuan 2019, 41, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Rollauer, S.E.; Sooreshjani, M.A.; Noinaj, N.; Buchanan, S.K. Outer membrane protein biogenesis in Gram-negative bacteria. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370. [Google Scholar] [CrossRef]
- Rosengarten, R.; Behrens, A.; Stetefeld, A.; Heller, M.; Ahrens, M.; Sachse, K.; Yogev, D.; Kirchhoff, H. Antigen heterogeneity among isolates of Mycoplasma bovis is generated by high-frequency variation of diverse membrane surface proteins. Infect. Immun. 1994, 62, 5066–5074. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.; Li, Y.; Wang, Y.; Zhou, Y.; Liu, Y.; Xin, J. Molecular cloning and characterization of a surface-localized adhesion protein in Mycoplasma bovis Hubei-1 strain. PLoS ONE 2013, 8, e69644. [Google Scholar] [CrossRef] [Green Version]
- Aparicio, D.; Torres-Puig, S.; Ratera, M.; Querol, E.; Pinol, J.; Pich, O.Q.; Fita, I. Mycoplasma genitalium adhesin P110 binds sialic-acid human receptors. Nat. Commun. 2018, 9, 4471. [Google Scholar] [CrossRef] [Green Version]
- Yeung, C.Y.; Chiang Chiau, J.S.; Chan, W.T.; Jiang, C.B.; Cheng, M.L.; Liu, H.L.; Lee, H.C. In vitro prevention of salmonella lipopolysaccharide-induced damages in epithelial barrier function by various lactobacillus strains. Gastroenterol. Res. Pract. 2013, 2013, 973209. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Zhao, Z.; Zhang, L.; Ma, H.; Lu, K.; Ren, W.; Liu, Z.; Chang, H.; Bei, W.; Qiu, Y.; et al. Trigger factor of Streptococcus suis is involved in stress tolerance and virulence. Microb. Pathog. 2011, 51, 69–76. [Google Scholar] [CrossRef]
- Liu, W.; Huang, L.; Su, Y.; Qin, Y.; Zhao, L.; Yan, Q. Contributions of the oligopeptide permeases in multistep of Vibrio alginolyticus pathogenesis. Microbiologyopen 2017, 6. [Google Scholar] [CrossRef]
- Theodoulou, F.L.; Kerr, I.D. ABC transporter research: Going strong 40 years on. Biochem. Soc. Trans. 2015, 43, 1033–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, F.; He, J.; Navarro-Alvarez, N.; Xu, J.; Li, X.; Li, P.; Wu, W. Correction: Elongation Factor Tu and Heat Shock Protein 70 Are Membrane-Associated Proteins from Mycoplasma ovipneumoniae Capable of Inducing Strong Immune Response in Mice. PLoS ONE 2017, 12, e0189562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Wang, H.; Wang, J.; Feng, Z.; Wu, M.; Liu, B.; Xin, J.; Xiong, Q.; Liu, M.; Shao, G. Elongation Factor Thermo Unstable (EF-Tu) Moonlights as an Adhesin on the Surface of Mycoplasma hyopneumoniae by Binding to Fibronectin. Front. Microbiol. 2018, 9, 974. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Oligonucleotide sequence (5′→3′) | Purpose |
|---|---|---|
| mTn For 1 | GACCTACACCGAACTGAGATACC | Mapping of mTn insertions |
| Link Rev 1 | CCAGTGTGCTGGAATTGCCC | |
| mTn For 2 | CCTGCGTTATCCCCTGATTCTG | |
| Link Rev 2 | GCAGATATCCATCACACTGGCG | |
| mTn Seq 3 | CCTTTGAGTGAGCTGATACCGCTC | |
| 0503A1 | CGCGGATCCTTCAAAATTAATTTAGAAAAGAAAAATGTAATTAG | Mbov_0503 cloning |
| 0503A2 | TATAGTTAGGCGTAAAGCT[C]CAGTATATA | Mutagenesis |
| 0503B1 | TTACGCCTAACTATAATGAAAAAAATATG | |
| 0503B2 | CATCTTTAAAATAATA[C]CAGCCTGGGT | |
| 0503C1 | TTAAAGATGGCATTTTACACTTAATAATTG[G] | |
| 0503C2 | TTACTGTCTAAAAA[C]CACTGATAATACTTTG | |
| 0503D1 | TTTAGACAGTAATGAGCATAACGAACAC | |
| 0503D2 | TTATATAAATATCTAAA[C]CAAGGGACATTT | |
| 0503E1 | AGATATTTATATAATAATTTATG[G]ACCGAAAATT | |
| 0503E2 | CAGTTGAAAGTAAATTTTCGGT[C]CATAAATTATT | |
| 0503F1 | CTTTCAACTGAAGTAAACAGTGATGATT | |
| 0503F2 | CCGCTCGAGTTATACATTATAAAAATATTTTATTTTTAATCTTTGTACA | Mbov_0503 cloning |
| Mu0503F | CTCCAATACTCTAAGTGGTCTAAGT | PCR amplification of Mbov_0503 |
| Mu0503R | CATCTTTAAAATAATATCAGCCTGGGT |
| Genomic Position a | CDS b | Relative CDS Position c | Mutant d | Predicted CDS Identity |
|---|---|---|---|---|
| 035818 (-) | Mbov_0034 | 0.07 (+) | T5.407 | oligopeptide ABC transporter ATP-binding protein (oppD) |
| 068606 (+) | Mbov_0059 | 0.99 (-) | T5.182 | tRNA modification GTPase (trmE) |
| 175200 (-) | Mbov_0154 | 0.61 (-) | T5.187 | transmembrane protein |
| 191067 (+) | Mbov_0168 | 0.31 (-) | T5.201 | trigger factor (tig) |
| 359714 (-) | Mbov_0305 | 0.41 (+) | T5.230 | transmembrane protein |
| 420981 (+) | Mbov_0353 | 0.45 (+) | T5.212 | alcohol dehydrogenase (adh) |
| 436842 (+) | Mbov_0370 | 0.38 (+) | T5.313 | tRNA-methyltransferase (trmU) |
| 573330 (+) | Mbov_0490 | 0.97 (+) | T5.495 | ATP-binding cassette subfamily B |
| 586897 (-) | Mbov_0503 | 0.23 (-) | T4.4 | transmembrane protein |
| CDS | Signal Peptide a | Transmembrane Domain a | SecP Score c | Subcellular Localization (Score) d |
|---|---|---|---|---|
| Mbov_0034 | No | No | 0.800 | CM (9.99) |
| Mbov_0059 | No | No | - | C (9.97) |
| Mbov_0154 | No | Yes | 0.918 | unknown |
| Mbov_0168 | No | No | - | C (7.5) |
| Mbov_0305 | No | Yes | 0.512 | C (7.5) |
| Mbov_0353 | No | No | - | C (9.97) |
| Mbov_0370 | No | No | 0.593 | C (9.97) |
| Mbov_0490 | Yes | Yes | 0.926 | CM (10) |
| Mbov_0503 | No | Yes | 0.657 | unknown |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, X.; Dong, Y.; Baranowski, E.; Li, X.; Zhao, G.; Hao, Z.; Zhang, H.; Chen, Y.; Hu, C.; Chen, H.; et al. Mbov_0503 Encodes a Novel Cytoadhesin that Facilitates Mycoplasma bovis Interaction with Tight Junctions. Microorganisms 2020, 8, 164. https://doi.org/10.3390/microorganisms8020164
Zhu X, Dong Y, Baranowski E, Li X, Zhao G, Hao Z, Zhang H, Chen Y, Hu C, Chen H, et al. Mbov_0503 Encodes a Novel Cytoadhesin that Facilitates Mycoplasma bovis Interaction with Tight Junctions. Microorganisms. 2020; 8(2):164. https://doi.org/10.3390/microorganisms8020164
Chicago/Turabian StyleZhu, Xifang, Yaqi Dong, Eric Baranowski, Xixi Li, Gang Zhao, Zhiyu Hao, Hui Zhang, Yingyu Chen, Changmin Hu, Huanchun Chen, and et al. 2020. "Mbov_0503 Encodes a Novel Cytoadhesin that Facilitates Mycoplasma bovis Interaction with Tight Junctions" Microorganisms 8, no. 2: 164. https://doi.org/10.3390/microorganisms8020164
APA StyleZhu, X., Dong, Y., Baranowski, E., Li, X., Zhao, G., Hao, Z., Zhang, H., Chen, Y., Hu, C., Chen, H., Citti, C., & Guo, A. (2020). Mbov_0503 Encodes a Novel Cytoadhesin that Facilitates Mycoplasma bovis Interaction with Tight Junctions. Microorganisms, 8(2), 164. https://doi.org/10.3390/microorganisms8020164