Measles: An Overview of a Re-Emerging Disease in Children and Immunocompromised Patients


 , ,
, ,  ,
,
Abstract
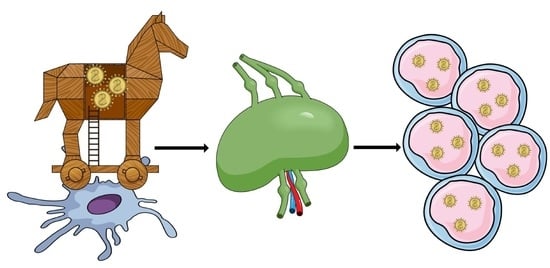
:
1. Introduction
2. The Pathogen
3. Epidemiology
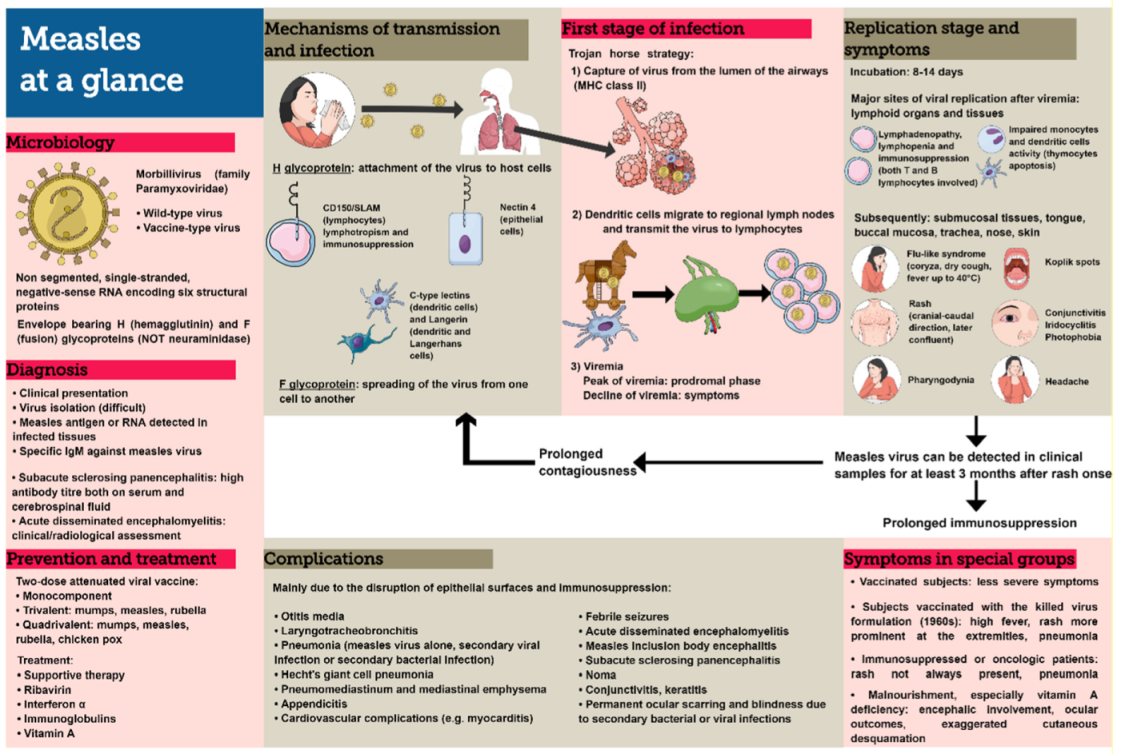
4. Pathophysiology
4.1. First Stage of Infection
4.2. Replication Stage
5. Mode of Transmission
6. Signs and Symptoms
6.1. Complications
6.2. Clinical Manifestation in Special Groups
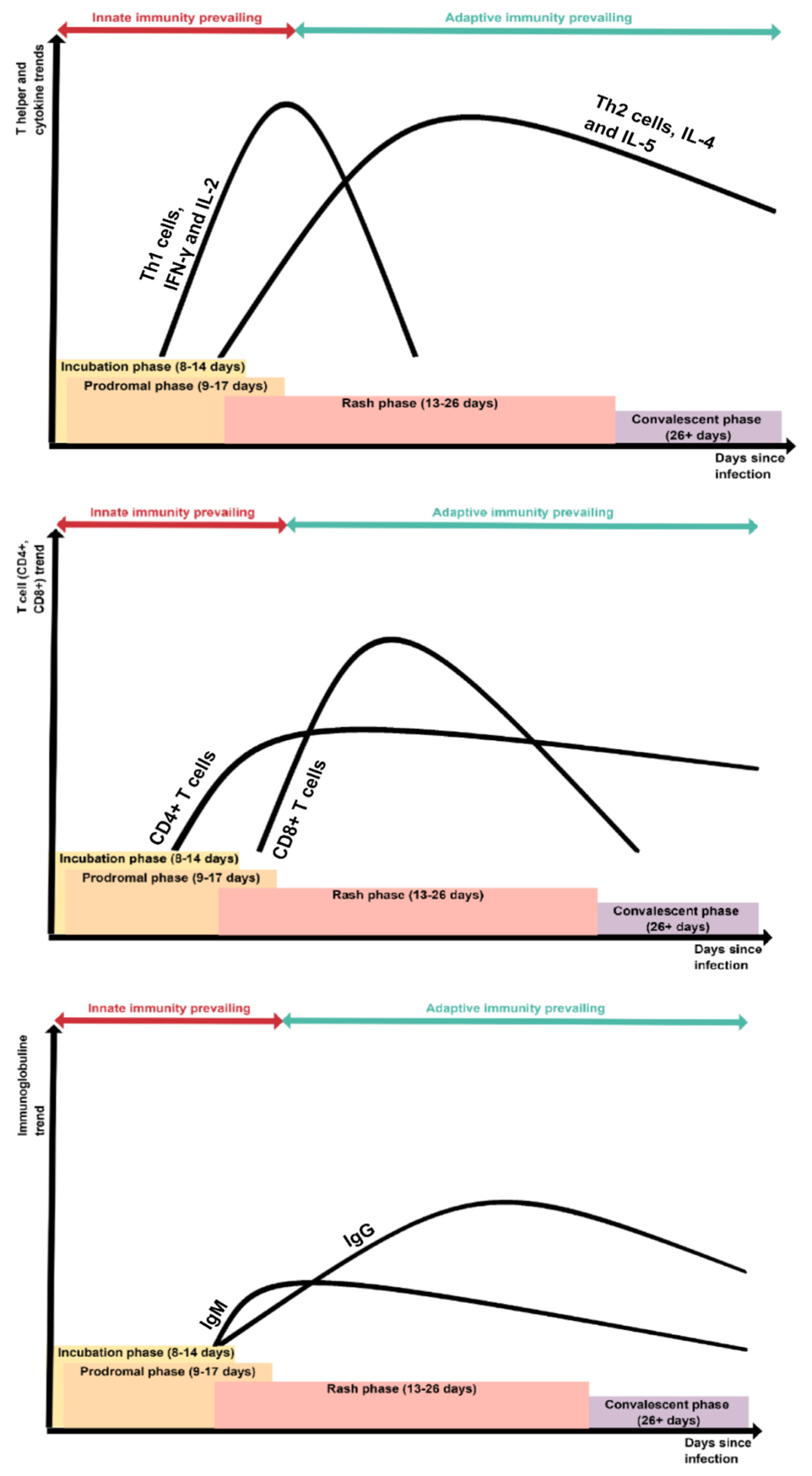
7. Measles and Immunity
8. Pregnancy
9. Diagnosis
10. Vaccine
Adverse Effects of Vaccination
11. Therapy
12. Summary and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Moss, W.J. Measles. Lancet 2017, 390, 2490–2502. [Google Scholar] [CrossRef]
- Perry, R.T.; Halsey, N.A. The clinical significance of measles: A review. J. Infect. Dis. 2004, 189, S4–S16. [Google Scholar]
- Kaplan, L.J.; Daum, R.S.; Smaron, M.; McCarthy, C.A. Severe measles in immunocompromised patients. JAMA 1992, 267, 1237–1241. [Google Scholar] [CrossRef]
- Rota, P.A.; Liffick, S.L.; Rota, J.S.; Katz, R.S.; Redd, S.; Papania, M.; Bellini, W.J. Molecular epidemiology of measles viruses in the United States, 1997–2001. Emerg. Infect. Dis. 2002, 8, 902–908. [Google Scholar] [CrossRef]
- El Najjar, F.; Schmitt, A.P.; Dutch, R.E. Paramyxovirus glycoprotein incorporation, assembly and budding: A three way dance for infectious particle production. Viruses 2014, 6, 3019–3054. [Google Scholar] [CrossRef] [Green Version]
- Tatsuo, H.; Ono, N.; Tanaka, K.; Yanagi, Y. Slam (cdw150) is a cellular receptor for measles virus. Nature 2000, 406, 893–897. [Google Scholar] [CrossRef]
- Muhlebach, M.D.; Mateo, M.; Sinn, P.L.; Prufer, S.; Uhlig, K.M.; Leonard, V.H.; Navaratnarajah, C.K.; Frenzke, M.; Wong, X.X.; Sawatsky, B.; et al. Adherens junction protein nectin-4 is the epithelial receptor for measles virus. Nature 2011, 480, 530–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kempe, C.H.; Fulginiti, V.A. The pathogenesis of measles virus infection. Arch. Gesamte Virusforsch. 1965, 16, 103–128. [Google Scholar] [CrossRef] [PubMed]
- Fine, P.E.; Clarkson, J.A. Measles in england and wales—I: An analysis of factors underlying seasonal patterns. Int. J. Epidemiol. 1982, 11, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Clements, C.J.; Strassburg, M.; Cutts, F.T.; Torel, C. The epidemiology of measles. World Health Stat. Q. 1992, 45, 285–291. [Google Scholar]
- Wallinga, J.; Heijne, J.C.; Kretzschmar, M. A measles epidemic threshold in a highly vaccinated population. PLoS Med. 2005, 2, e316. [Google Scholar] [CrossRef] [PubMed]
- Sugerman, D.E.; Barskey, A.E.; Delea, M.G.; Ortega-Sanchez, I.R.; Bi, D.; Ralston, K.J.; Rota, P.A.; Waters-Montijo, K.; Lebaron, C.W. Measles outbreak in a highly vaccinated population, San Diego, 2008: Role of the intentionally undervaccinated. Pediatrics 2010, 125, 747–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lancella, L.; Di Camillo, C.; Vittucci, A.C.; Boccuzzi, E.; Bozzola, E.; Villani, A. Measles lessons in an anti-vaccination era: Public health is a social duty, not a political option. Ital. J. Pediatrics 2017, 43, 102. [Google Scholar] [CrossRef] [Green Version]
- De Vries, R.D.; Lemon, K.; Ludlow, M.; McQuaid, S.; Yuksel, S.; van Amerongen, G.; Rennick, L.J.; Rima, B.K.; Osterhaus, A.D.; de Swart, R.L.; et al. In vivo tropism of attenuated and pathogenic measles virus expressing green fluorescent protein in macaques. J. Virol. 2010, 84, 4714–4724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Messling, V.; Svitek, N.; Cattaneo, R. Receptor (slam [cd150]) recognition and the v protein sustain swift lymphocyte-based invasion of mucosal tissue and lymphatic organs by a morbillivirus. J. Virol. 2006, 80, 6084–6092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Witte, L.; Abt, M.; Schneider-Schaulies, S.; van Kooyk, Y.; Geijtenbeek, T.B. Measles virus targets dc-sign to enhance dendritic cell infection. J. Virol. 2006, 80, 3477–3486. [Google Scholar] [CrossRef] [Green Version]
- Noyce, R.S.; Bondre, D.G.; Ha, M.N.; Lin, L.T.; Sisson, G.; Tsao, M.S.; Richardson, C.D. Tumor cell marker pvrl4 (nectin 4) is an epithelial cell receptor for measles virus. PLoS Pathog. 2011, 7, e1002240. [Google Scholar] [CrossRef]
- Andres, O.; Obojes, K.; Kim, K.S.; ter Meulen, V.; Schneider-Schaulies, J. Cd46- and cd150-independent endothelial cell infection with wild-type measles viruses. J. Gen. Virol. 2003, 84, 1189–1197. [Google Scholar] [CrossRef]
- Lemon, K.; de Vries, R.D.; Mesman, A.W.; McQuaid, S.; van Amerongen, G.; Yuksel, S.; Ludlow, M.; Rennick, L.J.; Kuiken, T.; Rima, B.K.; et al. Early target cells of measles virus after aerosol infection of non-human primates. PLoS Pathog. 2011, 7, e1001263. [Google Scholar] [CrossRef]
- De Swart, R.L.; Ludlow, M.; de Witte, L.; Yanagi, Y.; van Amerongen, G.; McQuaid, S.; Yuksel, S.; Geijtenbeek, T.B.; Duprex, W.P.; Osterhaus, A.D. Predominant infection of cd150+ lymphocytes and dendritic cells during measles virus infection of macaques. PLoS Pathog. 2007, 3, e178. [Google Scholar] [CrossRef]
- Laksono, B.M.; de Vries, R.D.; McQuaid, S.; Duprex, W.P.; de Swart, R.L. Measles virus host invasion and pathogenesis. Viruses 2016, 8, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludlow, M.; Rennick, L.J.; Sarlang, S.; Skibinski, G.; McQuaid, S.; Moore, T.; de Swart, R.L.; Duprex, W.P. Wild-type measles virus infection of primary epithelial cells occurs via the basolateral surface without syncytium formation or release of infectious virus. J. Gen. Virol. 2010, 91, 971–979. [Google Scholar] [CrossRef] [PubMed]
- De Vries, R.D.; Yuksel, S.; Osterhaus, A.D.; de Swart, R.L. Specific cd8(+) t-lymphocytes control dissemination of measles virus. Eur. J. Immunol. 2010, 40, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Riddell, M.A.; Moss, W.J.; Hauer, D.; Monze, M.; Griffin, D.E. Slow clearance of measles virus rna after acute infection. J. Clin. Virol. 2007, 39, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Sartwell, P.E. The incubation period and the dynamics of infectious disease. Am. J. Epidemiol. 1966, 83, 204–206. [Google Scholar] [CrossRef] [PubMed]
- Laksono, B.M.; de Vries, R.D.; Verburgh, R.J.; Visser, E.G.; de Jong, A.; Fraaij, P.L.A.; Ruijs, W.L.M.; Nieuwenhuijse, D.F.; van den Ham, H.J.; Koopmans, M.P.G.; et al. Studies into the mechanism of measles-associated immune suppression during a measles outbreak in the netherlands. Nat. Commun. 2018, 9, 4944. [Google Scholar] [CrossRef]
- Moss, W.J.; Strebel, P. Biological feasibility of measles eradication. J. Infect. Dis. 2011, 204, S47–S53. [Google Scholar] [CrossRef]
- Ferrari, M.J.; Grais, R.F.; Bharti, N.; Conlan, A.J.; Bjornstad, O.N.; Wolfson, L.J.; Guerin, P.J.; Djibo, A.; Grenfell, B.T. The dynamics of measles in sub-saharan Africa. Nature 2008, 451, 679–684. [Google Scholar] [CrossRef] [Green Version]
- Phadke, V.K.; Bednarczyk, R.A.; Salmon, D.A.; Omer, S.B. Association between vaccine refusal and vaccine-preventable diseases in the United States: A review of measles and pertussis. JAMA 2016, 315, 1149–1158. [Google Scholar] [CrossRef] [Green Version]
- Koplik, H. The diagnosis of the invasion of measles from a study of the exanthema as it appears on the buccal mucous membrane. Arch. Pediatr. 1962, 79, 162–165. [Google Scholar]
- Suringa, D.W.; Bank, L.J.; Ackerman, A.B. Role of measles virus in skin lesions and koplik’s spots. N. Engl. J. Med. 1970, 283, 1139–1142. [Google Scholar] [CrossRef] [PubMed]
- Robbins, F.C. Measles: Clinical features. Pathogenesis, pathology and complications. Am. J. Dis. Child. 1962, 103, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Scheifele, D.W.; Forbes, C.E. Prolonged giant cell excretion in severe African measles. Pediatrics 1972, 50, 867–873. [Google Scholar] [PubMed]
- Morley, D.C. Measles in the developing world. Proc. R. Soc. Med. 1974, 67, 1112–1115. [Google Scholar] [CrossRef]
- Hutchins, S.S.; Papania, M.J.; Amler, R.; Maes, E.F.; Grabowsky, M.; Bromberg, K.; Glasglow, V.; Speed, T.; Bellini, W.J.; Orenstein, W.A. Evaluation of the measles clinical case definition. J. Infect. Dis. 2004, 189, S153–S159. [Google Scholar] [PubMed]
- Dinh, A.; Fleuret, V.; Hanslik, T. Liver involvement in adults with measles. Int. J. Infect. Dis. 2013, 17, e1243–e1244. [Google Scholar] [CrossRef] [Green Version]
- Mohiuddin, S.A.; AlMaslamani, M.; Hashim, S.; Panthalayinitharayil, H.K.; Alkaabi, S.R.; Abdulwahab, A.; Derbala, M. Measles hepatitis in a vaccinated liver transplant recipient: Case report and review of literature. Clin. Case Rep. 2017, 5, 867–870. [Google Scholar] [CrossRef]
- Hussey, G.D.; Clements, C.J. Clinical problems in measles case management. Ann. Trop. Paediatr. 1996, 16, 307–317. [Google Scholar] [CrossRef]
- Quiambao, B.P.; Gatchalian, S.R.; Halonen, P.; Lucero, M.; Sombrero, L.; Paladin, F.J.; Meurman, O.; Merin, J.; Ruutu, P. Coinfection is common in measles-associated pneumonia. Pediatr. Infect. Dis. J. 1998, 17, 89–93. [Google Scholar] [CrossRef]
- Gremillion, D.H.; Crawford, G.E. Measles pneumonia in young adults. An analysis of 106 cases. Am. J. Med. 1981, 71, 539–542. [Google Scholar] [CrossRef]
- Loukides, S.; Panagou, P.; Kolokouris, D.; Kalogeropoulos, N. Bacterial pneumonia as a suprainfection in young adults with measles. Eur. Respir. J. 1999, 13, 356–360. [Google Scholar] [CrossRef] [PubMed]
- Yalaburgi, S.B. Subcutaneous and mediastinal emphysema following respiratory tract complications in measles. S. Afr. Med. J. 1980, 58, 521–524. [Google Scholar] [PubMed]
- Pancharoen, C.; Ruttanamongkol, P.; Suwangool, P.; Likitnukul, S.; Thisyakorn, U. Measles-associated appendicitis: Two case reports and literature review. Scand. J. Infect. Dis. 2001, 33, 632–633. [Google Scholar] [PubMed]
- Finkel, H.E. Measles myocarditis. Am. Heart J. 1964, 67, 679–683. [Google Scholar] [CrossRef]
- Greenberg, B.L.; Sack, R.B.; Salazar-Lindo, E.; Budge, E.; Gutierrez, M.; Campos, M.; Visberg, A.; Leon-Barua, R.; Yi, A.; Maurutia, D.; et al. Measles-associated diarrhea in hospitalized children in lima, Peru: Pathogenic agents and impact on growth. J. Infect. Dis. 1991, 163, 495–502. [Google Scholar] [CrossRef]
- Miller, D.L. Frequency of complications of measles, 1963. Report on a national inquiry by the public health laboratory service in collaboration with the society of medical officers of health. Br. Med. J. 1964, 2, 75–78. [Google Scholar] [CrossRef] [Green Version]
- Hanninen, P.; Arstila, P.; Lang, H.; Salmi, A.; Panelius, M. Involvement of the central nervous system in acute, uncomplicated measles virus infection. J. Clin. Microbiol. 1980, 11, 610–613. [Google Scholar] [CrossRef] [Green Version]
- Bitnun, A.; Shannon, P.; Durward, A.; Rota, P.A.; Bellini, W.J.; Graham, C.; Wang, E.; Ford-Jones, E.L.; Cox, P.; Becker, L.; et al. Measles inclusion-body encephalitis caused by the vaccine strain of measles virus. Clin. Infect. Dis. 1999, 29, 855–861. [Google Scholar] [CrossRef] [Green Version]
- Wendorf, K.A.; Winter, K.; Zipprich, J.; Schechter, R.; Hacker, J.K.; Preas, C.; Cherry, J.D.; Glaser, C.; Harriman, K. Subacute sclerosing panencephalitis: The devastating measles complication that might be more common than previously estimated. Clin. Infect. Dis. 2017, 65, 226–232. [Google Scholar] [CrossRef]
- Bellini, W.J.; Rota, J.S.; Lowe, L.E.; Katz, R.S.; Dyken, P.R.; Zaki, S.R.; Shieh, W.J.; Rota, P.A. Subacute sclerosing panencephalitis: More cases of this fatal disease are prevented by measles immunization than was previously recognized. J. Infect. Dis. 2005, 192, 1686–1693. [Google Scholar] [CrossRef]
- Garg, R.K. Subacute sclerosing panencephalitis. Postgrad. Med. J. 2002, 78, 63–70. [Google Scholar] [CrossRef]
- Breitfeld, V.; Hashida, Y.; Sherman, F.E.; Odagiri, K.; Yunis, E.J. Fatal measles infection in children with leukemia. Lab. Invest. 1973, 28, 279–291. [Google Scholar] [PubMed]
- Foster, A.; Sommer, A. Childhood blindness from corneal ulceration in Africa: Causes, prevention, and treatment. Bull. World Health Organ. 1986, 64, 619–623. [Google Scholar]
- Permar, S.R.; Griffin, D.E.; Letvin, N.L. Immune containment and consequences of measles virus infection in healthy and immunocompromised individuals. Clin. Vaccine Immunol. 2006, 13, 437–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Good, R.A.; Zak, S.J. Disturbances in gamma globulin synthesis as experiments of nature. Pediatrics 1956, 18, 109–149. [Google Scholar] [PubMed]
- Chen, R.T.; Markowitz, L.E.; Albrecht, P.; Stewart, J.A.; Mofenson, L.M.; Preblud, S.R.; Orenstein, W.A. Measles antibody: Reevaluation of protective titers. J. Infect. Dis. 1990, 162, 1036–1042. [Google Scholar] [CrossRef] [PubMed]
- Permar, S.R.; Moss, W.J.; Ryon, J.J.; Monze, M.; Cutts, F.; Quinn, T.C.; Griffin, D.E. Prolonged measles virus shedding in human immunodeficiency virus-infected children, detected by reverse transcriptase-polymerase chain reaction. J. Infect. Dis. 2001, 183, 532–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forthal, D.N.; Landucci, G.; Habis, A.; Zartarian, M.; Katz, J.; Tilles, J.G. Measles virus-specific functional antibody responses and viremia during acute measles. J. Infect. Dis. 1994, 169, 1377–1380. [Google Scholar] [CrossRef] [PubMed]
- van Binnendijk, R.S.; Versteeg-van Oosten, J.P.; Poelen, M.C.; Brugghe, H.F.; Hoogerhout, P.; Osterhaus, A.D.; Uytdehaag, F.G. Human hla class i- and hla class ii-restricted cloned cytotoxic t lymphocytes identify a cluster of epitopes on the measles virus fusion protein. J. Virol. 1993, 67, 2276–2284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, C.P.; Ammerlaan, W.; Fleckenstein, B.; Krauss, S.; Kalbacher, H.; Schneider, F.; Jung, G.; Wiesmuller, K.H. Activation of t cells by the ragged tail of mhc class ii-presented peptides of the measles virus fusion protein. Int. Immunol. 1996, 8, 445–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pette, M.; Liebert, U.G.; Gobel, U.; Grosse-Wilde, H.; Hartung, H.P.; Toyka, K.V. Measles virus-directed responses of cd4+ t lymphocytes in ms patients and healthy individuals. Neurology 1993, 43, 2019–2025. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.E.; Ward, B.J.; Jauregui, E.; Johnson, R.T.; Vaisberg, A. Immune activation in measles. N. Engl. J. Med. 1989, 320, 1667–1672. [Google Scholar] [CrossRef] [PubMed]
- Yanagi, Y.; Cubitt, B.A.; Oldstone, M.B. Measles virus inhibits mitogen-induced t cell proliferation but does not directly perturb the t cell activation process inside the cell. Virology 1992, 187, 280–289. [Google Scholar] [CrossRef]
- Sanchez-Lanier, M.; Guerin, P.; McLaren, L.C.; Bankhurst, A.D. Measles virus-induced suppression of lymphocyte proliferation. Cell. Immunol. 1988, 116, 367–381. [Google Scholar] [CrossRef]
- Bell, A.F.; Burns, J.B.; Fujinami, R.S. Measles virus infection of human t cells modulates cytokine generation and il-2 receptor alpha chain expression. Virology 1997, 232, 241–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubois, B.; Lamy, P.J.; Chemin, K.; Lachaux, A.; Kaiserlian, D. Measles virus exploits dendritic cells to suppress cd4+ t-cell proliferation via expression of surface viral glycoproteins independently of t-cell trans-infection. Cell. Immunol. 2001, 214, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Grosjean, I.; Caux, C.; Bella, C.; Berger, I.; Wild, F.; Banchereau, J.; Kaiserlian, D. Measles virus infects human dendritic cells and blocks their allostimulatory properties for cd4+ t cells. J. Exp. Med. 1997, 186, 801–812. [Google Scholar] [CrossRef]
- Petrova, V.N.; Sawatsky, B.; Han, A.X.; Laksono, B.M.; Walz, L.; Parker, E.; Pieper, K.; Anderson, C.A.; de Vries, R.D.; Lanzavecchia, A.; et al. Incomplete genetic reconstitution of b cell pools contributes to prolonged immunosuppression after measles. Sci. Immunol. 2019, 4. [Google Scholar] [CrossRef]
- Mina, M.J.; Kula, T.; Leng, Y.; Li, M.; de Vries, R.D.; Knip, M.; Siljander, H.; Rewers, M.; Choy, D.F.; Wilson, M.S.; et al. Measles virus infection diminishes preexisting antibodies that offer protection from other pathogens. Science 2019, 366, 599–606. [Google Scholar] [CrossRef] [Green Version]
- Moroi, K.; Saito, S.; Kurata, T.; Sata, T.; Yanagida, M. Fetal death associated with measles virus infection of the placenta. Am. J. Obstet. Gynecol. 1991, 164, 1107–1108. [Google Scholar] [CrossRef]
- Kobayashi, K.; Tajima, M.; Toishi, S.; Fujimori, K.; Suzuki, Y.; Udagawa, H. Fetal growth restriction associated with measles virus infection during pregnancy. J. Perinat. Med. 2005, 33, 67–68. [Google Scholar] [CrossRef] [PubMed]
- Siegel, M. Congenital malformations following chickenpox, measles, mumps, and hepatitis. Results of a cohort study. JAMA 1973, 226, 1521–1524. [Google Scholar] [CrossRef] [PubMed]
- Bar-On, S.; Ochshorn, Y.; Halutz, O.; Aboudy, Y.; Many, A. Detection of measles virus by reverse-transcriptase polymerase chain reaction in a placenta. J. Matern.-Fetal Neonatal Med. 2010, 23, 935–937. [Google Scholar] [CrossRef] [PubMed]
- Anselem, O.; Tsatsaris, V.; Lopez, E.; Krivine, A.; Le Ray, C.; Loulergue, P.; Floret, D.; Goffinet, F.; Launay, O. measles and pregnancy]. Presse Med. 2011, 40, 1001–1007. [Google Scholar] [CrossRef]
- White, S.J.; Boldt, K.L.; Holditch, S.J.; Poland, G.A.; Jacobson, R.M. Measles, mumps, and rubella. Clin. Obstet. Gynecol. 2012, 55, 550–559. [Google Scholar] [CrossRef]
- Garg, R.K.; Mahadevan, A.; Malhotra, H.S.; Rizvi, I.; Kumar, N.; Uniyal, R. Subacute sclerosing panencephalitis. Rev. Med. Virol. 2019, 29, e2058. [Google Scholar] [CrossRef]
- Bennetto, L.; Scolding, N. Inflammatory/post-infectious encephalomyelitis. J. Neurol. Neurosurg. Psychiatry 2004, 75, i22–i28. [Google Scholar] [CrossRef] [Green Version]
- Demicheli, V.; Rivetti, A.; Debalini, M.G.; Di Pietrantonj, C. Vaccines for measles, mumps and rubella in children. Evid.-Based Child Health A Cochrane Rev. J. 2013, 8, 2076–2238. [Google Scholar] [CrossRef]
- Ma, S.J.; Li, X.; Xiong, Y.Q.; Yao, A.L.; Chen, Q. Combination measles-mumps-rubella-varicella vaccine in healthy children: A systematic review and meta-analysis of immunogenicity and safety. Medicine 2015, 94, e1721. [Google Scholar] [CrossRef]
- World Health Organization. Measles vaccines: Who position paper, April 2017—Recommendations. Vaccine 2019, 37, 219–222. [Google Scholar] [CrossRef]
- Bowes, J. Measles, misinformation, and risk: Personal belief exemptions and the mmr vaccine. J. Law Biosci. 2016, 3, 718–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Serres, G.; Boulianne, N.; Defay, F.; Brousseau, N.; Benoit, M.; Lacoursiere, S.; Guillemette, F.; Soto, J.; Ouakki, M.; Ward, B.J.; et al. Higher risk of measles when the first dose of a 2-dose schedule of measles vaccine is given at 12–14 months versus 15 months of age. Clin. Infect. Dis. 2012, 55, 394–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nic Lochlainn, L.M.; de Gier, B.; van der Maas, N.; van Binnendijk, R.; Strebel, P.M.; Goodman, T.; de Melker, H.E.; Moss, W.J.; Hahne, S.J.M. Effect of measles vaccination in infants younger than 9 months on the immune response to subsequent measles vaccine doses: A systematic review and meta-analysis. Lancet Infect. Dis. 2019, 19, 1246–1254. [Google Scholar] [CrossRef] [Green Version]
- Gay, N.J. The theory of measles elimination: Implications for the design of elimination strategies. J. Infect. Dis. 2004, 189, S27–S35. [Google Scholar] [PubMed]
- Griffin, D.E. The immune response in measles: Virus control, clearance and protective immunity. Viruses 2016, 8, 282. [Google Scholar] [CrossRef] [PubMed]
- Plotkin, S.A. Correlates of protection induced by vaccination. Clin. Vaccine Immunol. 2010, 17, 1055–1065. [Google Scholar] [CrossRef] [Green Version]
- Haralambieva, I.H.; Ovsyannikova, I.G.; O’Byrne, M.; Pankratz, V.S.; Jacobson, R.M.; Poland, G.A. A large observational study to concurrently assess persistence of measles specific b-cell and t-cell immunity in individuals following two doses of mmr vaccine. Vaccine 2011, 29, 4485–4491. [Google Scholar] [CrossRef] [Green Version]
- Hammarlund, E.; Thomas, A.; Amanna, I.J.; Holden, L.A.; Slayden, O.D.; Park, B.; Gao, L.; Slifka, M.K. Plasma cell survival in the absence of b cell memory. Nat. Commun. 2017, 8, 1781. [Google Scholar] [CrossRef] [Green Version]
- Radbruch, A.; Muehlinghaus, G.; Luger, E.O.; Inamine, A.; Smith, K.G.; Dorner, T.; Hiepe, F. Competence and competition: The challenge of becoming a long-lived plasma cell. Nat. Rev. Immunol. 2006, 6, 741–750. [Google Scholar] [CrossRef]
- Haralambieva, I.H.; Ovsyannikova, I.G.; Pankratz, V.S.; Kennedy, R.B.; Jacobson, R.M.; Poland, G.A. The genetic basis for interindividual immune response variation to measles vaccine: New understanding and new vaccine approaches. Expert Rev. Vaccines 2013, 12, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Bester, J.C. Measles and measles vaccination: A review. JAMA Pediatr. 2016, 170, 1209–1215. [Google Scholar] [CrossRef] [PubMed]
- Forni, A.L.; Schluger, N.W.; Roberts, R.B. Severe measles pneumonitis in adults: Evaluation of clinical characteristics and therapy with intravenous ribavirin. Clin. Infect. Dis. 1994, 19, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Ortac Ersoy, E.; Tanriover, M.D.; Ocal, S.; Ozisik, L.; Inkaya, C.; Topeli, A. Severe measles pneumonia in adults with respiratory failure: Role of ribavirin and high-dose vitamin A. Clin. Respir. J. 2016, 10, 673–675. [Google Scholar] [CrossRef] [PubMed]
- Wyde, P.R. Chemotherapy of respiratory viruses: Prospects and challenges. Drug Resist. Updat. 1999, 2, 244–258. [Google Scholar] [CrossRef] [PubMed]
- Barclay, A.J.; Foster, A.; Sommer, A. Vitamin A supplements and mortality related to measles: A randomised clinical trial. Br. Med. J. (Clin. Res. Ed.) 1987, 294, 294–296. [Google Scholar] [CrossRef] [Green Version]
- D’Souza, R.M.; D’Souza, R. Vitamin A for treating measles in children. Cochrane Database Syst. Rev. 2002, 4, CD001479. [Google Scholar]
- Rupp, M.E.; Schwartz, M.L.; Bechard, D.E. Measles pneumonia. Treatment of a near-fatal case with corticosteroids and vitamin A. Chest 1993, 103, 1625–1626. [Google Scholar] [CrossRef]
- Bichon, A.; Aubry, C.; Benarous, L.; Drouet, H.; Zandotti, C.; Parola, P.; Lagier, J.C. Case report: Ribavirin and vitamin A in a severe case of measles. Medicine 2017, 96, e9154. [Google Scholar] [CrossRef]
- Benecke, O.; DeYoung, S.E. Anti-vaccine decision-making and measles resurgence in the United States. Glob. Pediatr. Health 2019, 6, 2333794X19862949. [Google Scholar] [CrossRef] [Green Version]
- Shelby, A.; Ernst, K. Story and science: How providers and parents can utilize storytelling to combat anti-vaccine misinformation. Hum. Vaccines Immunother. 2013, 9, 1795–1801. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Measles Vaccination | Measles Infection | ||
|---|---|---|---|
| Total side effects per 1 million doses administered | 0.0033% | Total complications per 1 million of infected children | 30% |
| Main side effects | Main complications | ||
| Joint pain | 25% | Diarrhea | 8% |
| Fever | 5–15% | Otitis media | 7% |
| Rash | 5% | Pneumonia | 5% |
| Febrile seizures | 0.3–0.8% | Primary measles encephalitis | 0.1–0.3% |
| Thrombocytopenia | 0.003% | Acute postinfectious encephalomyelitis | 0.1% |
| Anaphylaxis | 0.0001% | Subacute sclerosing panencephalitis | 0.01% |
| Encephalitis | 0.00002% | Death | 0.2% |
| Parotitis | --- | ||
| Lymphadenopathy | --- | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Misin, A.; Antonello, R.M.; Di Bella, S.; Campisciano, G.; Zanotta, N.; Giacobbe, D.R.; Comar, M.; Luzzati, R. Measles: An Overview of a Re-Emerging Disease in Children and Immunocompromised Patients. Microorganisms 2020, 8, 276. https://doi.org/10.3390/microorganisms8020276
Misin A, Antonello RM, Di Bella S, Campisciano G, Zanotta N, Giacobbe DR, Comar M, Luzzati R. Measles: An Overview of a Re-Emerging Disease in Children and Immunocompromised Patients. Microorganisms. 2020; 8(2):276. https://doi.org/10.3390/microorganisms8020276
Chicago/Turabian StyleMisin, Andrea, Roberta Maria Antonello, Stefano Di Bella, Giuseppina Campisciano, Nunzia Zanotta, Daniele Roberto Giacobbe, Manola Comar, and Roberto Luzzati. 2020. "Measles: An Overview of a Re-Emerging Disease in Children and Immunocompromised Patients" Microorganisms 8, no. 2: 276. https://doi.org/10.3390/microorganisms8020276
APA StyleMisin, A., Antonello, R. M., Di Bella, S., Campisciano, G., Zanotta, N., Giacobbe, D. R., Comar, M., & Luzzati, R. (2020). Measles: An Overview of a Re-Emerging Disease in Children and Immunocompromised Patients. Microorganisms, 8(2), 276. https://doi.org/10.3390/microorganisms8020276