Safety Aspects of Genetically Modified Lactic Acid Bacteria


{kind=link}
Abstract
:1. Introduction
2. ‘Generally Recognized as Safe’ Status: Definition and Determination
3. The Consequence of Genome Modification
4. Acceptable Methods for Genome Modification in the European Union
4.1. Natural Methods
4.2. Random Mutagenesis
5. Unacceptable Methods for Genome Modification in the European Union
5.1. Transformation with Plasmids and Genome Integration
5.2. Recombineering
5.3. CRISPR-Cas9–Supported Genome Rearrangement, Recombineering, and Gene Integration
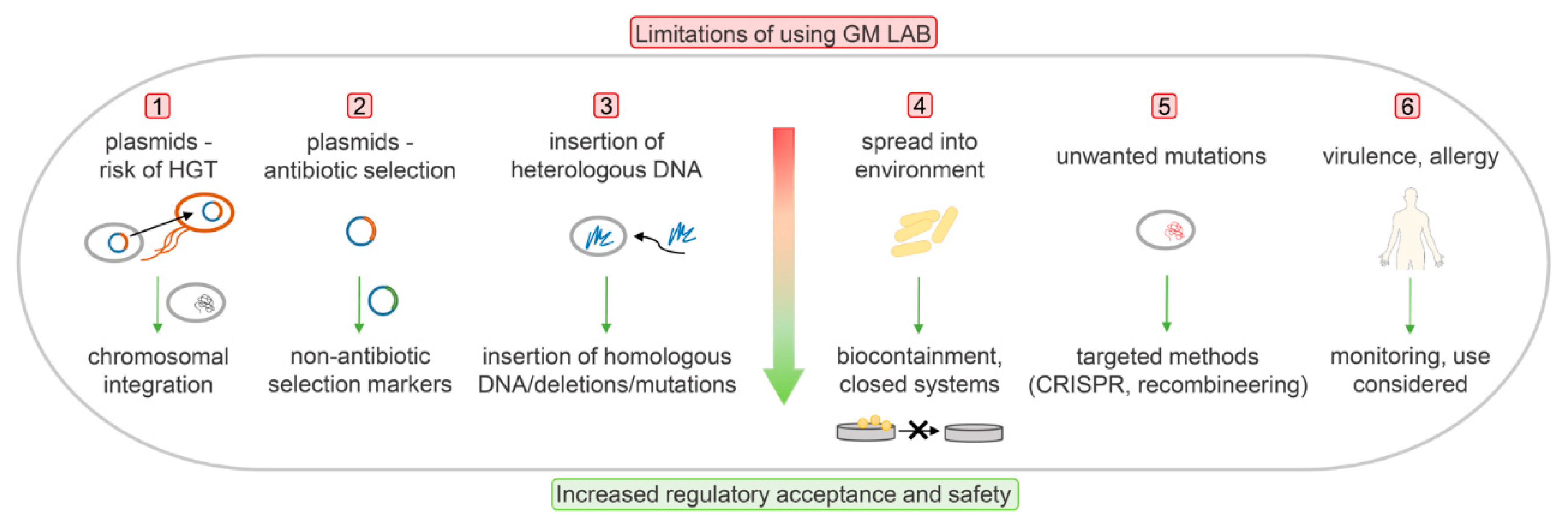
6. The Main Issues with Regard to Genetically Modified Lactic Acid Bacteria, and the Attempts to Address Them
6.1. Biocontainment
6.2. Antibiotic Resistance in Lactic Acid Bacteria and its Use in the Selection of Genetically Modified Organisms
6.3. Antibiotic-Free Selection Systems
6.4. Use of Homologous DNA
6.5. Virulence Factors
6.6. Delayed Adverse Effects
6.7. Surface Display
7. Genetically Modified Lactic Acid Bacteria as Cell Factories
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stiles, M.E.; Holzapfel, W.H. Lactic acid bacteria of foods and their current taxonomy. Int. J. Food Microbiol. 1997, 36, 1–29. [Google Scholar] [CrossRef]
- Chowdhury, M.Y.; Li, R.; Kim, J.H.; Park, M.E.; Kim, T.H.; Pathinayake, P.; Weeratunga, P.; Song, M.K.; Son, H.Y.; Hong, S.P.; et al. Mucosal vaccination with recombinant Lactobacillus casei-displayed CTA1-conjugated consensus matrix protein-2 (sM2) induces broad protection against divergent influenza subtypes in BALB/c mice. PLoS ONE 2014, 9, e94051. [Google Scholar] [CrossRef]
- Van Braeckel-Budimir, N.; Haijema, B.J.; Leenhouts, K. Bacterium-like particles for efficient immune stimulation of existing vaccines and new subunit vaccines in mucosal applications. Front. Immunol. 2013, 4, 282. [Google Scholar] [CrossRef] [Green Version]
- William Reed. NutraIngredients.com. Available online: https://www.nutraingredients.com/Article/2018/02/22/Consumer-acceptance-Novel-probiotics-are-beneficial-but-the-food-industry-is-its-own-worst-enemy-on-GM-technologies?utm_source=copyright&utm_medium=OnSite&utm_campaign=copyright (accessed on 18 November 2019).
- Ortiz-Velez, L.; Britton, R. Genetic tools for the enhancement of probiotic properties. Microbiol. Spectr. 2017, 5, 371–387. [Google Scholar] [CrossRef]
- Brodmann, T.; Endo, A.; Gueimonde, M.; Vinderola, G.; Kneifel, W.; de Vos, W.M.; Salminen, S.; Gomez-Gallego, C. Safety of novel microbes for human consumption: Practical examples of assessment in the European Union. Front. Microbiol. 2017, 8, 1725. [Google Scholar] [CrossRef]
- Laulund, S.; Wind, A.; Derkx, P.M.F.; Zuliani, V. Regulatory and safety requirements for food cultures. Microorganisms 2017, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, O.; O’Callaghan, J.; Sangrador-Vegas, A.; McAuliffe, O.; Slattery, L.; Kaleta, P.; Callanan, M.; Fitzgerald, G.F.; Ross, R.P.; Beresford, T. Comparative genomics of lactic acid bacteria reveals a niche-specific gene set. BMC Microbiol. 2009, 9, 50. [Google Scholar] [CrossRef] [Green Version]
- Platteeuw, C.; van Alen-Boerrigter, I.; van Schalkwijk, S.; de Vos, W.M. Food-grade cloning and expression system for Lactococcus lactis. Appl. Environ. Microbiol. 1996, 62, 1008–1013. [Google Scholar] [CrossRef] [Green Version]
- Borner, R.A.; Kandasamy, V.; Axelsen, A.M.; Nielsen, A.T.; Bosma, E.F. Genome editing of lactic acid bacteria: Opportunities for food, feed, pharma and Biotechnol. FEMS Microbiol. Lett. 2019, 366, 31–40. [Google Scholar] [CrossRef]
- Hatti-Kaul, R.; Chen, L.; Dishisha, T.; Enshasy, H.E. Lactic acid bacteria: From starter cultures to producers of chemicals. FEMS Microbiol. Lett. 2018, 366, 365. [Google Scholar] [CrossRef] [Green Version]
- Tauer, C.; Heinl, S.; Egger, E.; Heiss, S.; Grabherr, R. Tuning constitutive recombinant gene expression in Lactobacillus plantarum. Microb. Cell Fact. 2014, 13, 150. [Google Scholar] [CrossRef]
- Ogaugwu, C.E.; Cheng, Q.; Fieck, A.; Hurwitz, I.; Durvasula, R. Characterization of a Lactococcus lactis promoter for heterologous protein production. Biotechnol. Rep. 2017, 17, 86–92. [Google Scholar] [CrossRef]
- de Vos, W.M. Gene expression systems for lactic acid bacteria. Curr. Opin. Microbiol. 1999, 2, 289–295. [Google Scholar] [CrossRef]
- Mays, Z.J.; Nair, N.U. Synthetic biology in probiotic lactic acid bacteria: At the frontier of living therapeutics. Curr. Opin. Biotechnol. 2018, 53, 224–231. [Google Scholar] [CrossRef]
- Dahmane, N.; Robert, E.; Deschamps, J.; Meylheuc, T.; Delorme, C.; Briandet, R.; Leblond-Bourget, N.; Guedon, E.; Payot, S. Impact of cell surface molecules on conjugative transfer of the integrative and conjugative element ICESt3 of Streptococcus thermophilus. Appl. Environ. Microbiol. 2018, 84, 17. [Google Scholar]
- Chiang, Y.N.; Penades, J.R.; Chen, J. Genetic transduction by phages and chromosomal islands: The new and noncanonical. PLoS Pathog. 2019, 15, e1007878. [Google Scholar] [CrossRef] [Green Version]
- Baugher, J.L.; Durmaz, E.; Klaenhammer, T.R. Spontaneously induced prophages in Lactobacillus gasseri contribute to horizontal gene transfer. Appl. Environ. Microbiol. 2014, 80, 3508–3517. [Google Scholar] [CrossRef] [Green Version]
- Bron, P.A.; Marcelli, B.; Mulder, J.; van der Els, S.; Morawska, L.P.; Kuipers, O.P.; Kok, J.; Kleerebezem, M. Renaissance of traditional DNA transfer strategies for improvement of industrial lactic acid bacteria. Curr. Opin. Biotechnol. 2019, 56, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Samson, J.E.; Moineau, S. Characterization of Lactococcus lactis phage 949 and comparison with other lactococcal phages. Appl. Environ. Microbiol. 2010, 76, 6843–6852. [Google Scholar] [CrossRef] [Green Version]
- Pujato, S.A.; Guglielmotti, D.M.; Martinez-Garcia, M.; Quiberoni, A.; Mojica, F.J.M. Leuconostoc mesenteroides and Leuconostoc pseudomesenteroides bacteriophages: Genomics and cross-species host ranges. Int. J. Food Microbiol. 2017, 257, 128–137. [Google Scholar] [CrossRef] [Green Version]
- Blokesch, M. Natural competence for transformation. Curr. Biol. 2016, 26, R1126–R1130. [Google Scholar] [CrossRef] [PubMed]
- Derkx, P.M.; Janzen, T.; Sorensen, K.I.; Christensen, J.E.; Stuer-Lauridsen, B.; Johansen, E. The art of strain improvement of industrial lactic acid bacteria without the use of recombinant DNA technology. Microb. Cell Fact. 2014, 13, S5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Vestergaard, M.; Jensen, T.G.; Shen, J.; Dufva, M.; Solem, C.; Jensen, P.R. Finding the needle in the haystack-the use of microfluidic droplet technology to identify vitamin-secreting lactic acid bacteria. mBio 2017, 8, e00526-517. [Google Scholar] [CrossRef] [Green Version]
- Goodarzi, A. UV—Induced mutagenesis in lactic acid bacteria. Int. J. Genet. Genom. 2016, 4, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Arihara, K.; Itoh, M. UV-induced Lactobacillus gasseri mutants resisting sodium chloride and sodium nitrite for meat fermentation. Int. J. Food Microbiol. 2000, 56, 227–230. [Google Scholar] [CrossRef]
- Almalki, M.A. Production of medically important lactic acid by Lactobacillus Pentosus: A biological conversion method. Indian J. Sci. Technol. 2016, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Mierau, I.; Kleerebezem, M. 10 years of the nisin-controlled gene expression system (NICE) in Lactococcus lactis. Appl. Microbiol. Biotechnol. 2005, 68, 705–717. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Nguyen, H.M.; Geiger, B.; Mathiesen, G.; Eijsink, V.G.; Peterbauer, C.K.; Haltrich, D.; Nguyen, T.H. Heterologous expression of a recombinant lactobacillal beta-galactosidase in Lactobacillus plantarum: Effect of different parameters on the sakacin P-based expression system. Microb. Cell Fact. 2015, 14, 30. [Google Scholar] [CrossRef] [Green Version]
- Karlskas, I.L.; Maudal, K.; Axelsson, L.; Rud, I.; Eijsink, V.G.; Mathiesen, G. Heterologous protein secretion in Lactobacilli with modified pSIP vectors. PLoS ONE 2014, 9, e91125. [Google Scholar] [CrossRef] [Green Version]
- Sorvig, E.; Mathiesen, G.; Naterstad, K.; Eijsink, V.G.; Axelsson, L. High-level, inducible gene expression in Lactobacillus sakei and Lactobacillus plantarum using versatile expression vectors. Microbiology 2005, 151, 2439–2449. [Google Scholar] [CrossRef] [Green Version]
- Walker, D.C.; Klaenhammer, T.R. Isolation of a novel IS3 group insertion element and construction of an integration vector for Lactobacillus spp. J. Bacteriol. 1994, 176, 5330–5340. [Google Scholar] [CrossRef] [Green Version]
- Law, J.; Buist, G.; Haandrikman, A.; Kok, J.; Venema, G.; Leenhouts, K. A system to generate chromosomal mutations in Lactococcus lactis which allows fast analysis of targeted genes. J. Bacteriol. 1995, 177, 7011–7018. [Google Scholar] [CrossRef] [Green Version]
- Russell, W.M.; Klaenhammer, T.R. Efficient system for directed integration into the Lactobacillus acidophilus and Lactobacillus gasseri chromosomes via homologous recombination. Appl. Environ. Microbiol. 2001, 67, 4361–4364. [Google Scholar] [CrossRef] [Green Version]
- Neu, T.; Henrich, B. New thermosensitive delivery vector and its use to enable nisin-controlled gene expression in Lactobacillus gasseri. Appl. Environ. Microbiol. 2003, 69, 1377–1382. [Google Scholar] [CrossRef] [Green Version]
- Goh, Y.J.; Azcarate-Peril, M.A.; O’Flaherty, S.; Durmaz, E.; Valence, F.; Jardin, J.; Lortal, S.; Klaenhammer, T.R. Development and application of a upp-based counterselective gene replacement system for the study of the S-layer protein SlpX of Lactobacillus acidophilus NCFM. Appl. Environ. Microbiol. 2009, 75, 3093–3105. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Cui, H.; Tang, L.; Qiao, X.; Liu, M.; Jiang, Y.; Cui, W.; Li, Y. Construction of upp deletion mutant strains of Lactobacillus casei and Lactococcus lactis based on counterselective system using temperature-sensitive plasmid. J. Microbiol. Meth. 2014, 102, 37–44. [Google Scholar] [CrossRef]
- Lambert, J.M.; Bongers, R.S.; Kleerebezem, M. Cre-lox-based system for multiple gene deletions and selectable-marker removal in Lactobacillus plantarum. Appl. Environ. Microbiol. 2007, 73, 1126–1135. [Google Scholar] [CrossRef] [Green Version]
- Campo, N.; Daveran-Mingot, M.L.; Leenhouts, K.; Ritzenthaler, P.; Le Bourgeois, P. Cre-loxP recombination system for large genome rearrangements in Lactococcus lactis. Appl. Environ. Microbiol. 2002, 68, 2359–2367. [Google Scholar] [CrossRef] [Green Version]
- van Pijkeren, J.P.; Britton, R.A. High efficiency recombineering in lactic acid bacteria. Nucleic Acids Res. 2012, 40, e76. [Google Scholar] [CrossRef]
- Guo, T.; Xin, Y.; Zhang, Y.; Gu, X.; Kong, J. A rapid and versatile tool for genomic engineering in Lactococcus lactis. Microb. Cell Fact. 2019, 18, 22. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.H.; van Pijkeren, J.P. CRISPR-Cas9-assisted recombineering in Lactobacillus reuteri. Nucleic Acids Res. 2014, 42, e131. [Google Scholar] [CrossRef]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and applications of CRISPR-Cas9 for genome engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doudna, J.A.; Charpentier, E. Genome editing. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef] [PubMed]
- Selle, K.; Klaenhammer, T.R.; Barrangou, R. CRISPR-based screening of genomic island excision events in bacteria. Proc. Natl. Acad. Sci. USA 2015, 112, 8076–8081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Els, S.; James, J.K.; Kleerebezem, M.; Bron, P.A. Development of a versatile Cas9-driven subpopulation-selection toolbox in Lactococcus lactis. Appl. Environ. Microbiol. 2018, 84, 8. [Google Scholar] [CrossRef]
- Berlec, A.; Škrlec, K.; Kocjan, J.; Olenic, M.; Štrukelj, B. Single plasmid systems for inducible dual protein expression and for CRISPR-Cas9/CRISPRi gene regulation in lactic acid bacterium Lactococcus lactis. Sci. Rep. 2018, 8, 1009. [Google Scholar] [CrossRef]
- Song, X.; Huang, H.; Xiong, Z.; Ai, L.; Yang, S. CRISPR-Cas9(D10A) nickase-assisted genome editing in Lactobacillus casei. Appl. Environ. Microbiol. 2017, 83, 17. [Google Scholar] [CrossRef] [Green Version]
- Qi, L.S.; Larson, M.H.; Gilbert, L.A.; Doudna, J.A.; Weissman, J.S.; Arkin, A.P.; Lim, W.A. Repurposing CRISPR as an RNA-guided platform for sequence-specific control of gene expression. Cell 2013, 152, 1173–1183. [Google Scholar] [CrossRef] [Green Version]
- Mougiakos, I.; Mohanraju, P.; Bosma, E.F.; Vrouwe, V.; Finger Bou, M.; Naduthodi, M.I.S.; Gussak, A.; Brinkman, R.B.L.; van Kranenburg, R.; van der Oost, J. Characterizing a thermostable Cas9 for bacterial genome editing and silencing. Nat. Commun. 2017, 8, 1647. [Google Scholar] [CrossRef] [Green Version]
- Nakade, S.; Yamamoto, T.; Sakuma, T. Cas9, Cpf1 and C2c1/2/3-What’s next? Bioengineered 2017, 8, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Qian, F.; Yang, J.; Liu, Y.; Dong, F.; Xu, C.; Sun, B.; Chen, B.; Xu, X.; Li, Y.; et al. CRISPR-Cpf1 assisted genome editing of Corynebacterium glutamicum. Nat. Commun. 2017, 8, 15179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, M.; Li, Z.; Wang, W.; Liu, J.; Liu, L.; Zhu, G.; Karthik, L.; Wang, M.; Wang, K.F.; Wang, Z.; et al. A CRISPR-Cas12a-derived biosensing platform for the highly sensitive detection of diverse small molecules. Nat. Commun. 2019, 10, 3672. [Google Scholar] [CrossRef]
- Shen, W.; Zhang, J.; Geng, B.; Qiu, M.; Hu, M.; Yang, Q.; Bao, W.; Xiao, Y.; Zheng, Y.; Peng, W.; et al. Establishment and application of a CRISPR-Cas12a assisted genome-editing system in Zymomonas mobilis. Microb. Cell Fact. 2019, 18, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.B.; Komor, A.C.; Levy, J.M.; Packer, M.S.; Zhao, K.T.; Liu, D.R. Increasing the genome-targeting scope and precision of base editing with engineered Cas9-cytidine deaminase fusions. Nat. Biotechnol. 2017, 35, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Gaudelli, N.M.; Komor, A.C.; Rees, H.A.; Packer, M.S.; Badran, A.H.; Bryson, D.I.; Liu, D.R. Programmable base editing of A*T to G*C in genomic DNA without DNA cleavage. Nature 2017, 551, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Eid, A.; Alshareef, S.; Mahfouz, M.M. CRISPR base editors: Genome editing without double-stranded breaks. Biochem. J. 2018, 475, 1955–1964. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, S.; Chen, W.; Song, L.; Zhang, Y.; Shen, Z.; Yu, F.; Li, M.; Ji, Q. CRISPR-Cas9 and CRISPR-assisted cytidine deaminase enable precise and efficient genome editing in Klebsiella pneumoniae. Appl. Environ. Microbiol. 2018, 84, 18. [Google Scholar] [CrossRef] [Green Version]
- Zheng, K.; Wang, Y.; Li, N.; Jiang, F.F.; Wu, C.X.; Liu, F.; Chen, H.C.; Liu, Z.F. Highly efficient base editing in bacteria using a Cas9-cytidine deaminase fusion. Commun. Biol. 2018, 1, 32. [Google Scholar] [CrossRef]
- Wilson, D.J. NIH guidelines for research involving recombinant DNA molecules. Account. Res. 1993, 3, 177–185. [Google Scholar] [CrossRef]
- Lee, P. Biocontainment strategies for live lactic acid bacteria vaccine vectors. Bioeng. Bugs 2010, 1, 75–77. [Google Scholar] [CrossRef] [Green Version]
- Steidler, L.; Neirynck, S.; Huyghebaert, N.; Snoeck, V.; Vermeire, A.; Goddeeris, B.; Cox, E.; Remon, J.P.; Remaut, E. Biological containment of genetically modified Lactococcus lactis for intestinal delivery of human interleukin 10. Nat. Biotechnol. 2003, 21, 785–789. [Google Scholar] [CrossRef]
- Bahey-El-Din, M.; Gahan, C.G. Lactococcus lactis: From the dairy industry to antigen and therapeutic protein delivery. Discov. Med. 2010, 9, 455–461. [Google Scholar]
- Zhou, H.; Li, X.; Wang, Z.; Yin, J.; Tan, H.; Wang, L.; Qiao, X.; Jiang, Y.; Cui, W.; Liu, M.; et al. Construction and characterization of thymidine auxotrophic (ΔthyA) recombinant Lactobacillus casei expressing bovine lactoferricin. BMC Vet. Res. 2018, 14, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanin, A.; Culligan, E.P.; Casey, P.G.; Bahey-El-Din, M.; Hill, C.; Gahan, C.G. Two-tiered biological containment strategy for Lactococcus lactis-based vaccine or immunotherapy vectors. Hum. Vaccines Immunother. 2014, 10, 333–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callura, J.M.; Dwyer, D.J.; Isaacs, F.J.; Cantor, C.R.; Collins, J.J. Tracking, tuning, and terminating microbial physiology using synthetic riboregulators. Proc. Natl. Acad. Sci. USA 2010, 107, 15898–15903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.W.; Chan, C.T.Y.; Slomovic, S.; Collins, J.J. Next-generation biocontainment systems for engineered organisms. Nat. Chem. Biol. 2018, 14, 530–537. [Google Scholar] [CrossRef]
- Gallagher, R.R.; Patel, J.R.; Interiano, A.L.; Rovner, A.J.; Isaacs, F.J. Multilayered genetic safeguards limit growth of microorganisms to defined environments. Nucleic Acids Res. 2015, 43, 1945–1954. [Google Scholar] [CrossRef]
- Mandell, D.J.; Lajoie, M.J.; Mee, M.T.; Takeuchi, R.; Kuznetsov, G.; Norville, J.E.; Gregg, C.J.; Stoddard, B.L.; Church, G.M. Biocontainment of genetically modified organisms by synthetic protein design. Nature 2015, 518, 55. [Google Scholar] [CrossRef]
- Huang, S.; Lee, A.J.; Tsoi, R.; Wu, F.; Zhang, Y.; Leong, K.W.; You, L. Coupling spatial segregation with synthetic circuits to control bacterial survival. Mol. Syst. Biol. 2016, 12, 859. [Google Scholar] [CrossRef]
- Alvarez, Y.; Ponce-Alquicira, E. Antibiotic Resistance in Lactic Acid Bacteria; IntechOpen: London, UK, 2018. [Google Scholar]
- Mendelson, M.; Matsoso, M.P. The World Health Organization global action plan for antimicrobial resistance. S. Afr. Med. J. 2015, 105, 325. [Google Scholar] [CrossRef] [Green Version]
- Markwart, R.; Willrich, N.; Haller, S.; Noll, I.; Koppe, U.; Werner, G.; Eckmanns, T.; Reuss, A. The rise in vancomycin-resistant Enterococcus faecium in Germany: Data from the German Antimicrobial Resistance Surveillance (ARS). Antimicrob. Resist. Infect. Control. 2019, 8, 147. [Google Scholar] [CrossRef] [PubMed]
- Faron, M.L.; Ledeboer, N.A.; Buchan, B.W. Resistance mechanisms, epidemiology, and approaches to screening for vancomycin-resistant Enterococcus in the health care setting. J. Clin. Microbiol. 2016, 54, 2436–2447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erginkaya, Z.; Turhan, E.U.; Tatli, D. Determination of antibiotic resistance of lactic acid bacteria isolated from traditional Turkish fermented dairy products. Iran. J. Vet. Res. 2018, 19, 53–56. [Google Scholar] [PubMed]
- Gad, G.F.; Abdel-Hamid, A.M.; Farag, Z.S. Antibiotic resistance in lactic acid bacteria isolated from some pharmaceutical and dairy products. Braz. J. Microbiol. 2014, 45, 25–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Pan, L.; Li, L.; Lu, J.; Kwok, L.; Menghe, B.; Zhang, H.; Zhang, W. Characterization of antibiotic resistance genes from Lactobacillus isolated from traditional dairy products. J. Food Sci. 2017, 82, 724–730. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhang, Z.Y.; Dong, K.; Yuan, J.P.; Guo, X.K. Antibiotic resistance of probiotic strains of lactic acid bacteria isolated from marketed foods and drugs. Biomed. Environ. Sci. 2009, 22, 401–412. [Google Scholar] [CrossRef]
- Doucet-Populaire, F.; Trieu-Cuot, P.; Andremont, A.; Courvalin, P. Conjugal transfer of plasmid DNA from Enterococcus faecalis to Escherichia coli in digestive tracts of gnotobiotic mice. Antimicrob. Agents Chemother. 1992, 36, 502–504. [Google Scholar] [CrossRef] [Green Version]
- Bolotin, A.; Quinquis, B.; Sorokin, A.; Ehrlich, D.S. Recent genetic transfer between Lactococcus lactis and enterobacteria. J. Bacteriol. 2004, 186, 6671–6677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mater, D.D.G.; Langella, P.; Corthier, G.; Flores, M.J. A probiotic Lactobacillus strain can acquire vancomycin resistance during digestive transit in mice. J. Mol. Microb. Biotechnol. 2008, 14, 123–127. [Google Scholar] [CrossRef]
- International Organization for Standardization. Milk and Milk Products: Determination of the Minimal Inhibitory Concentration (MIC) of Antibiotics Applicable to Bifidobacteria and Non-Enterococcal Lactic Acid Bacteria. Available online: https://www.iso.org/standard/46434.html (accessed on 18 November 2019).
- Clementi, F.; Aquilanti, L. Recent investigations and updated criteria for the assessment of antibiotic resistance in food lactic acid bacteria. Anaerobe 2011, 17, 394–398. [Google Scholar] [CrossRef]
- De, R. Metagenomics: Aid to combat antimicrobial resistance in diarrhea. Gut Pathog. 2019, 11, 47. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Handelsman, J. Biotechnological prospects from metagenomics. Curr. Opin. Biotechnol. 2003, 14, 303–310. [Google Scholar] [CrossRef]
- European Food Safety, A. Technical guidance - Update of the criteria used in the assessment of bacterial resistance to antibiotics of human or veterinary importance. EFSA J. 2008, 6, 732. [Google Scholar] [CrossRef]
- Cox, G.; Wright, G.D. Intrinsic antibiotic resistance: Mechanisms, origins, challenges and solutions. Int. J. Med. Microbiol. 2013, 303, 287–292. [Google Scholar] [CrossRef]
- Mignon, C.; Sodoyer, R.; Werle, B. Antibiotic-free selection in biotherapeutics: Now and forever. Pathogens 2015, 4, 157–181. [Google Scholar] [CrossRef]
- He, S.; Gong, F.; Zhang, D.; Guo, Y. Food-grade selection markers in lactic acid bacteria. TAF Prev. Med. Bull. 2012, 11, 1. [Google Scholar] [CrossRef] [Green Version]
- Hughes, B.F.; McKay, L.L. Deriving phage-insensitive Lactococci using a food-grade vector encoding phage and nisin resistance. J. Dairy Sci. 1992, 75, 914–923. [Google Scholar] [CrossRef]
- von Wright, A.; Wessels, S.; Tynkkynen, S.; Saarela, M. Isolation of a replication region of a large lactococcal plasmid and use in cloning of a nisin resistance determinant. Appl. Environ. Microbiol. 1990, 56, 2029–2035. [Google Scholar] [CrossRef] [Green Version]
- Takala, T.; Saris, P. A food-grade cloning vector for lactic acid bacteria based on the nisin immunity gene nisI. Appl. Microbiol. Biotechnol. 2002, 59, 467–471. [Google Scholar]
- Leelawatcharamas, V.; Chia, L.G.; Charoenchai, P.; Kunajakr, N.; Liu, C.-Q.; Dunn, N.W. Plasmid-encoded copper resistance in Lactococcus lactis. Biotechnol. Lett. 1997, 19, 639–643. [Google Scholar] [CrossRef]
- Liu, C.Q.; Su, P.; Khunajakr, N.; Deng, Y.M.; Sumual, S.; Kim, W.S.; Tandianus, J.E.; Dunn, N.W. Development of food-grade cloning and expression vectors for Lactococcus lactis. J. Appl. Microbiol. 2005, 98, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.Y.; Su, P.; Allison, G.E.; Liu, C.-Q.; Dunn, N.W. A potential food-grade cloning vector for Streptococcus thermophilus that uses cadmium resistance as the selectable marker. Appl. Environ. Microbiol. 2003, 69, 5767–5771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Demerdash, H.A.M.; Heller, K.J.; Geis, A. Application of the shsp gene, encoding a small heat shock protein, as a food-grade selection marker for lactic acid bacteria. Appl. Environ. Microbiol. 2003, 69, 4408–4412. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, K.I.; Larsen, R.; Kibenich, A.; Junge, M.P.; Johansen, E. A food-grade cloning system for industrial strains of Lactococcus lactis. Appl. Environ. Microbiol. 2000, 66, 1253–1258. [Google Scholar] [CrossRef] [Green Version]
- Bron, P.A.; Benchimol, M.G.; Lambert, J.; Palumbo, E.; Deghorain, M.; Delcour, J.; De Vos, W.M.; Kleerebezem, M.; Hols, P. Use of the alr gene as a food-grade selection marker in lactic acid bacteria. Appl. Environ. Microbiol. 2002, 68, 5663–5670. [Google Scholar] [CrossRef] [Green Version]
- Takala, T.; Saris, P.; Tynkkynen, S. Food-grade host/vector expression system for Lactobacillus casei based on complementation of plasmid-associated phospho-β-galactosidase gene lacG. Appl. Microbiol. Biotechnol. 2003, 60, 564–570. [Google Scholar] [CrossRef]
- Boucher, I.; Parrot, M.; Gaudreau, H.; Champagne, C.P.; Vadeboncoeur, C.; Moineau, S. Novel food-grade plasmid vector based on melibiose fermentation for the genetic engineering of Lactococcus lactis. Appl. Environ. Microbiol. 2002, 68, 6152–6161. [Google Scholar] [CrossRef] [Green Version]
- Labrie, S.; Bart, C.; Vadeboncoeur, C.; Moineau, S. Use of an α-galactosidase gene as a food-grade selection marker for Streptococcus thermophilus. J. Dairy Sci. 2005, 88, 2341–2347. [Google Scholar] [CrossRef]
- Jeong, D.-W.; Lee, J.-H.; Kim, K.H.; Lee, H.J. A food-grade expression/secretion vector for Lactococcus lactis that uses an α-galactosidase gene as a selection marker. Food Microbiol. 2006, 23, 468–475. [Google Scholar] [CrossRef]
- Peubez, I.; Chaudet, N.; Mignon, C.; Hild, G.; Husson, S.; Courtois, V.; De Luca, K.; Speck, D.; Sodoyer, R. Antibiotic-free selection in E. coli: New considerations for optimal design and improved production. Microb. Cell Fact. 2010, 9, 65. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Harrison, E.M.; Bi, D.; Tai, C.; He, X.; Ou, H.-Y.; Rajakumar, K.; Deng, Z. TADB: A web-based resource for type 2 toxin-antitoxin loci in bacteria and archaea. Nucleic Acids Res. 2011, 39, D606–D611. [Google Scholar] [CrossRef] [Green Version]
- Diago-Navarro, E.; Hernandez-Arriaga, A.M.; López-Villarejo, J.; Muñoz-Gómez, A.J.; Kamphuis, M.B.; Boelens, R.; Lemonnier, M.; Díaz-Orejas, R. parD toxin–antitoxin system of plasmid R1–basic contributions, biotechnological applications and relationships with closely-related toxin–antitoxin systems. FEBS J. 2010, 277, 3097–3117. [Google Scholar] [CrossRef]
- Kawano, M. Divergently overlapping cis-encoded antisense RNA regulating toxin-antitoxin systems from E. coli: Hok/sok, ldr/rdl, symE/symR. RNA Biol. 2012, 9, 1520–1527. [Google Scholar] [CrossRef] [Green Version]
- Weaver, K.E. The par toxin-antitoxin system from Enterococcus faecalis plasmid pAD1 and its chromosomal homologs. RNA Biol. 2012, 9, 1498–1503. [Google Scholar] [CrossRef] [Green Version]
- Weaver, K.E. The Type I toxin-antitoxin par locus from Enterococcus faecalis plasmid pAD1: RNA regulation by both cis- and trans-acting elements. Plasmid 2015, 78, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Mairhofer, J.; Pfaffenzeller, I.; Merz, D.; Grabherr, R. A novel antibiotic free plasmid selection system: Advances in safe and efficient DNA therapy. Biotechnol. J. 2008, 3, 83–89. [Google Scholar] [CrossRef]
- Pfaffenzeller, I.; Mairhofer, J.; Striedner, G.; Bayer, K.; Grabherr, R. Using ColE1-derived RNA I for suppression of a bacterially encoded gene: Implication for a novel plasmid addiction system. Biotechnol. J. 2006, 1, 675–681. [Google Scholar] [CrossRef]
- Stenler, S.; Blomberg, P.; Smith, C.I.E. Safety and efficacy of DNA vaccines: Plasmids vs. minicircles. Hum. Vaccin. Immunother. 2014, 10, 1306–1308. [Google Scholar] [CrossRef] [Green Version]
- Douglas, G.L.; Goh, Y.J.; Klaenhammer, T.R. Integrative food grade expression system for lactic acid bacteria. Methods Mol. Biol. 2011, 765, 373–387. [Google Scholar]
- Johansen, E. Future access and improvement of industrial lactic acid bacteria cultures. Microb. Cell Fact. 2017, 16, 230. [Google Scholar] [CrossRef] [Green Version]
- Phumkhachorn, P.; Rattanachaikunsopon, P. A broad host range food-grade cloning vector for lactic acid bacteria. Biologia 2016, 71, 457. [Google Scholar] [CrossRef]
- Tagliavia, M.; Nicosia, A. Advanced strategies for food-grade protein production: A new E. coli/lactic acid bacteria shuttle vector for improved cloning and food-grade expression. Microorganisms 2019, 7, 116. [Google Scholar] [CrossRef] [Green Version]
- Mathipa, M.G.; Thantsha, M.S. Probiotic engineering: Towards development of robust probiotic strains with enhanced functional properties and for targeted control of enteric pathogens. Gut Pathog. 2017, 9, 28. [Google Scholar] [CrossRef]
- Sybesma, W.; Hugenholtz, J.; Vos, W.M.D.; Smid, E.J. Safe use of genetically modified lactic acid bacteria in food. Bridging the gap between consumers, green groups, and industry. Electron. J. Biotechnol. 2006, 9, 424–448. [Google Scholar] [CrossRef] [Green Version]
- Salminen, M.K.; Tynkkynen, S.; Rautelin, H.; Saxelin, M.; Vaara, M.; Ruutu, P.; Sarna, S.; Valtonen, V.; Järvinen, A. Lactobacillus bacteremia during a rapid increase in probiotic use of Lactobacillus rhamnosus GG in Finland. Clin. Infect. Dis. 2002, 35, 1155–1160. [Google Scholar] [CrossRef] [Green Version]
- Zadravec, P.; Štrukelj, B.; Berlec, A. Heterologous surface display on lactic acid bacteria: Non-GMO alternative? Bioengineered 2015, 6, 179–183. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Kong, J.; Kong, W.; Guo, T.; Ji, M. Characterization of a novel LysM domain from Lactobacillus fermentum bacteriophage endolysin and its use as an anchor to display heterologous proteins on the surfaces of lactic acid bacteria. Appl. Environ. Microbiol. 2010, 76, 2410–2418. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Kong, J.; Sun, Z.; Han, L.; Kong, W.; Yang, P. Heterologous protein display on the cell surface of lactic acid bacteria mediated by the s-layer protein. Microb. Cell Fact. 2011, 10, 86. [Google Scholar] [CrossRef] [Green Version]
- Plavec, T.V.; Štrukelj, B.; Berlec, A. Screening for new surface anchoring domains for Lactococcus lactis. Front. Microbiol. 2019, 10, 1879. [Google Scholar] [CrossRef] [Green Version]
- Plavec, T.V.; Berlec, A. Surface anchoring on Lactococcus lactis by covalent isopeptide bond. Acta Chim. Slov. 2019, 66, 10. [Google Scholar] [CrossRef] [Green Version]
- Mao, R.; Wu, D.; Wang, Y. Surface display on lactic acid bacteria without genetic modification: Strategies and applications. Appl. Microbiol. Biotechnol. 2016, 100, 9407–9421. [Google Scholar] [CrossRef] [PubMed]
- Song, A.A.; In, L.L.A.; Lim, S.H.E.; Rahim, R.A. A review on Lactococcus lactis: From food to factory. Microb. Cell Fact. 2017, 16, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berlec, A.; Štrukelj, B. Novel applications of recombinant lactic acid bacteria in therapy and in metabolic engineering. Recent Pat. Biotechnol. 2009, 3, 77–87. [Google Scholar] [CrossRef]
- Hugenholtz, J.; Sybesma, W.; Nierop Groot, M.; Wisselink, W.; Ladero, V.; Burgess, K.; van Sinderen, D.; Piard, J.-C.; Eggink, G.; Smid, E.J.; et al. Metabolic engineering of lactic acid bacteria for the production of nutraceuticals. Lactic Acid Bacteria Genet. Metab. Appl. 2002, 82, 217–235. [Google Scholar]
- Liu, J.; Chan, S.H.J.; Chen, J.; Solem, C.; Jensen, P.R. Systems Biology-A guide for understanding and developing improved strains of lactic acid bacteria. Front. Microbiol. 2019, 10, 876. [Google Scholar] [CrossRef]
- Papagianni, M. Metabolic engineering of lactic acid bacteria for the production of industrially important cpmpounds. Comput. Struct. Biotechnol. 2012, 3, e201210003. [Google Scholar] [CrossRef] [Green Version]
- Kleerebezem, M.; Hugenholtz, J. Metabolic pathway engineering in lactic acid bacteria. Curr. Opin. Biotechnol. 2003, 14, 232–237. [Google Scholar] [CrossRef]
- Castro-Aguirre, E.; Iniguez-Franco, F.; Samsudin, H.; Fang, X.; Auras, R. Poly(lactic acid)-mass production, processing, industrial applications, and end of life. Adv. Drug Deliv. Rev. 2016, 107, 333–366. [Google Scholar] [CrossRef] [Green Version]
- Mack, D.R. D(-)-lactic acid-producing probiotics, D(-)-lactic acidosis and infants. Can. J. Gastroenterol. 2004, 18, 671–675. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Pei, X.; Lei, T.; Wang, Y.; Feng, Y. Genome shuffling enhanced L-lactic acid production by improving glucose tolerance of Lactobacillus rhamnosus. J. Biotechnol. 2008, 134, 154–159. [Google Scholar] [CrossRef]
- Yi, X.; Zhang, P.; Sun, J.; Tu, Y.; Gao, Q.; Zhang, J.; Bao, J. Engineering wild-type robust Pediococcus acidilactici strain for high titer L- and D-lactic acid production from corn stover feedstock. J. Biotechnol. 2016, 217, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Okano, K.; Hama, S.; Kihara, M.; Noda, H.; Tanaka, T.; Kondo, A. Production of optically pure D-lactic acid from brown rice using metabolically engineered Lactobacillus plantarum. Appl. Microbiol. Biotechnol. 2017, 101, 1869–1875. [Google Scholar] [CrossRef] [PubMed]
- Okano, K.; Zhang, Q.; Shinkawa, S.; Yoshida, S.; Tanaka, T.; Fukuda, H.; Kondo, A. Efficient production of optically pure D-lactic acid from raw corn starch by using a genetically modified L-lactate dehydrogenase gene-deficient and alpha-amylase-secreting Lactobacillus plantarum strain. Appl. Environ. Microbiol. 2009, 75, 462–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, Y.C.; Yuan, S.F.; Wang, C.A.; Huang, Y.J.; Guo, G.L.; Hwang, W.S. Production of optically pure L-lactic acid from lignocellulosic hydrolysate by using a newly isolated and D-lactate dehydrogenase gene-deficient Lactobacillus paracasei strain. Bioresour. Technol. 2015, 198, 651–657. [Google Scholar] [CrossRef]
- Kyla-Nikkila, K.; Hujanen, M.; Leisola, M.; Palva, A. Metabolic engineering of Lactobacillus helveticus CNRZ32 for production of pure L-(+)-lactic acid. Appl. Environ. Microbiol. 2000, 66, 3835–3841. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Dantoft, S.H.; Würtz, A.; Jensen, P.R.; Solem, C. A novel cell factory for efficient production of ethanol from dairy waste. Biotechnol. Biofuels 2016, 9, 33. [Google Scholar] [CrossRef] [Green Version]
- Florou-Paneri, P.; Christaki, E.; Bonos, E. Lactic Acid Bacteria as Source of Functional Ingredients; IntechOpen: Rijeka, Croatia, 2012; pp. 589–614. [Google Scholar]
- Linares, D.M.; Gómez, C.; Renes, E.; Fresno, J.M.; Tornadijo, M.E.; Ross, R.P.; Stanton, C. Lactic acid bacteria and Bifidobacteria with potential to design natural biofunctional health-promoting dairy foods. Front. Microbiol. 2017, 8, 846. [Google Scholar] [CrossRef]
- Zhang, J.; Caiyin, Q.; Feng, W.; Zhao, X.; Qiao, B.; Zhao, G.; Qiao, J. Enhance nisin yield via improving acid-tolerant capability of Lactococcus lactis F44. Sci. Rep. 2016, 6, 27973. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Mu, D.; Qiao, W.; Zhu, D.; Wang, X.; Liu, F.; Xu, H.; Saris, P.; Kuipers, O.P.; Qiao, M. Co-expression of nisin Z and leucocin C as a basis for effective protection against Listeria monocytogenes in pasteurized milk. Front. Microbiol. 2018, 9, 547. [Google Scholar] [CrossRef]
- Burgess, C.; O’Connell-Motherway, M.; Sybesma, W.; Hugenholtz, J.; van Sinderen, D. Riboflavin production in Lactococcus lactis: Potential for in situ production of vitamin-enriched foods. Appl. Environ. Microbiol. 2004, 70, 5769–5777. [Google Scholar] [CrossRef] [Green Version]
- LeBlanc, J.G.; Burgess, C.; Sesma, F.; de Giori, G.S.; van Sinderen, D. Ingestion of milk fermented by genetically modified Lactococcus lactis improves the riboflavin status of deficient rats. J. Dairy Sci. 2005, 88, 3435–3442. [Google Scholar] [CrossRef] [Green Version]
- Wegkamp, A.; van Oorschot, W.; de Vos, W.M.; Smid, E.J. Characterization of the role of para-aminobenzoic acid biosynthesis in folate production by Lactococcus lactis. Appl. Environ. Microbiol. 2007, 73, 2673–2681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wegkamp, A.; Starrenburg, M.; de Vos, W.M.; Hugenholtz, J.; Sybesma, W. Transformation of folate-consuming Lactobacillus gasseri into a folate producer. Appl. Environ. Microbiol. 2004, 70, 3146–3148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, F.; Wegkamp, A.; de Vos, W.M.; Smid, E.J.; Hugenholtz, J. High-level folate production in fermented foods by the B12 producer Lactobacillus reuteri JCM1112. Appl. Environ. Microbiol. 2008, 74, 3291–3294. [Google Scholar] [CrossRef] [Green Version]
- De Boeck, R.; Sarmiento-Rubiano, L.A.; Nadal, I.; Monedero, V.; Perez-Martinez, G.; Yebra, M.J. Sorbitol production from lactose by engineered Lactobacillus casei deficient in sorbitol transport system and mannitol-1-phosphate dehydrogenase. Appl. Microbiol. Biotechnol. 2010, 85, 1915–1922. [Google Scholar] [CrossRef]
- Ladero, V.; Ramos, A.; Wiersma, A.; Goffin, P.; Schanck, A.; Kleerebezem, M.; Hugenholtz, J.; Smid, E.J.; Hols, P. High-level production of the low-calorie sugar sorbitol by Lactobacillus plantarum through metabolic engineering. Appl. Environ. Microbiol. 2007, 73, 1864–1872. [Google Scholar] [CrossRef] [Green Version]
- Gaspar, P.; Neves, A.R.; Gasson, M.J.; Shearman, C.A.; Santos, H. High yields of 2,3-butanediol and mannitol in Lactococcus lactis through engineering of NAD(+) cofactor recycling. Appl. Environ. Microbiol. 2011, 77, 6826–6835. [Google Scholar] [CrossRef] [Green Version]
- Gaspar, P.; Carvalho, A.L.; Vinga, S.; Santos, H.; Neves, A.R. From physiology to systems metabolic engineering for the production of biochemicals by lactic acid bacteria. Biotechnol. Adv. 2013, 31, 764–788. [Google Scholar] [CrossRef]
- Mazzoli, R.; Bosco, F.; Mizrahi, I.; Bayer, E.A.; Pessione, E. Towards lactic acid bacteria-based biorefineries. Biotechnol. Adv. 2014, 32, 1216–1236. [Google Scholar] [CrossRef]
- Narita, J.; Ishida, S.; Okano, K.; Kimura, S.; Fukuda, H.; Kondo, A. Improvement of protein production in lactic acid bacteria using 5′-untranslated leader sequence of slpA from Lactobacillus acidophilus. Appl. Microbiol. Biotechnol. 2006, 73, 366–373. [Google Scholar] [CrossRef]
- Narita, J.; Nakahara, S.; Fukuda, H.; Kondo, A. Efficient production of L-(+)-lactic acid from raw starch by Streptococcus bovis 148. J. Biosci. Bioeng. 2004, 97, 423–425. [Google Scholar] [CrossRef]
- Okano, K.; Kimura, S.; Narita, J.; Fukuda, H.; Kondo, A. Improvement in lactic acid production from starch using α-amylase-secreting Lactococcus lactis cells adapted to maltose or starch. Appl. Microbiol. Biotechnol. 2007, 75, 1007–1013. [Google Scholar] [CrossRef]
- Okano, K.; Zhang, Q.; Yoshida, S.; Tanaka, T.; Ogino, C.; Fukuda, H.; Kondo, A. d-lactic acid production from cellooligosaccharides and β-glucan using l-LDH gene-deficient and endoglucanase-secreting Lactobacillus plantarum. Appl. Microbiol. Biotechnol. 2010, 85, 643–650. [Google Scholar] [CrossRef]
- Moraïs, S.; Shterzer, N.; Grinberg, I.R.; Mathiesen, G.; Eijsink, V.G.H.; Axelsson, L.; Lamed, R.; Bayer, E.A.; Mizrahi, I. Establishment of a simple Lactobacillus plantarum cell consortium for cellulase-xylanase synergistic interactions. Appl. Environ. Microbiol. 2013, 79, 5242–5249. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.S.; Choi, Y.J.; Chung, D.K. Expression of Clostridium thermocellum endoglucanase gene in Lactobacillus gasseri and Lactobacillus johnsonii and characterization of the genetically modified probiotic Lactobacilli. Curr. Microbiol. 2000, 40, 257–263. [Google Scholar] [CrossRef]
- Raha, A.R.; Chang, L.Y.; Sipat, A.; Yusoff, K.; Haryanti, T. Expression of a thermostable xylanase gene from Bacillus coagulans ST-6 in Lactococcus lactis. Lett. Appl. Microbiol. 2006, 42, 210–214. [Google Scholar] [CrossRef]
- Ozkose, E.; Akyol, I.; Kar, B.; Comlekcioglu, U.; Ekinci, M.S. Expression of fungal cellulase gene in Lactococcus lactis to construct novel recombinant silage inoculants. Folia Microbiol. 2009, 54, 335–342. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Splechtna, B.; Yamabhai, M.; Haltrich, D.; Peterbauer, C. Cloning and expression of the beta-galactosidase genes from Lactobacillus reuteri in Escherichia coli. J. Biotechnol. 2007, 129, 581–591. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Nguyen, H.A.; Arreola, S.L.; Mlynek, G.; Djinovic-Carugo, K.; Mathiesen, G.; Nguyen, T.H.; Haltrich, D. Homodimeric beta-galactosidase from Lactobacillus delbrueckii subsp. bulgaricus DSM 20081: Expression in Lactobacillus plantarum and biochemical characterization. J. Agric. Food Chem. 2012, 60, 1713–1721. [Google Scholar]
- Halbmayr, E.; Mathiesen, G.; Nguyen, T.H.; Maischberger, T.; Peterbauer, C.K.; Eijsink, V.G.; Haltrich, D. High-level expression of recombinant beta-galactosidases in Lactobacillus plantarum and Lactobacillus sakei using a sakacin P-based expression system. J. Agric. Food Chem. 2008, 56, 4710–4719. [Google Scholar] [CrossRef]
- Nguyen, H.M.; Mathiesen, G.; Stelzer, E.M.; Pham, M.L.; Kuczkowska, K.; Mackenzie, A.; Agger, J.W.; Eijsink, V.G.; Yamabhai, M.; Peterbauer, C.K.; et al. Display of a beta-mannanase and a chitosanase on the cell surface of Lactobacillus plantarum towards the development of whole-cell biocatalysts. Microb. Cell Fact. 2016, 15, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, P.; Li, Y.; Wang, Y.; Meng, K.; Luo, H.; Yuan, T.; Bai, Y.; Zhan, Z.; Yao, B. A novel beta-mannanase with high specific activity from Bacillus circulans CGMCC1554: Gene cloning, expression and enzymatic characterization. Appl. Biochem. Biotechnol. 2009, 159, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Sak-Ubol, S.; Namvijitr, P.; Pechsrichuang, P.; Haltrich, D.; Nguyen, T.H.; Mathiesen, G.; Eijsink, V.G.; Yamabhai, M. Secretory production of a beta-mannanase and a chitosanase using a Lactobacillus plantarum expression system. Microb. Cell Fact. 2016, 15, 81. [Google Scholar] [CrossRef] [Green Version]
- Chien, L.-J.; Lee, C.-K. Hyaluronic acid production by recombinant Lactococcus lactis. Appl. Microbiol. Biotechnol. 2007, 77, 339–346. [Google Scholar] [CrossRef]
- Prasad, S.B.; Jayaraman, G.; Ramachandran, K.B. Hyaluronic acid production is enhanced by the additional co-expression of UDP-glucose pyrophosphorylase in Lactococcus lactis. Appl. Microbiol. Biotechnol. 2010, 86, 273–283. [Google Scholar] [CrossRef]
- Thakur, K.; Tomar, S.K.; De, S. Lactic acid bacteria as a cell factory for riboflavin production. Microb. Biotechnol. 2016, 9, 441–451. [Google Scholar] [CrossRef]
- Eric, J. Use of Natural selection and evolution to develop new starter cultures for fermented foods. Annu. Rev. Food Sci. Technol. 2018, 9, 411–428. [Google Scholar]
- Carvalho, R.D.; do Carmo, F.L.R.; de Oliveira Junior, A.; Langella, P.; Chatel, J.-M.; Bermúdez-Humarán, L.G.; Azevedo, V.; de Azevedo, M.S. Use of wild type or recombinant lactic acid bacteria as an alternative treatment for gastrointestinal inflammatory diseases: A focus on inflammatory bowel diseases and mucositis. Front. Microbiol. 2017, 8, 800. [Google Scholar] [CrossRef]
- de Moreno de LeBlanc, A.; Del Carmen, S.; Chatel, J.-M.; Miyoshi, A.; Azevedo, V.; Langella, P.; Bermúdez-Humarán, L.G.; LeBlanc, J.G. Current review of genetically modified lactic acid bacteria for the prevention and treatment of colitis using murine models. Gastroent. Res. Pract. 2015, 2015, 146972. [Google Scholar] [CrossRef]
- Plavec, T.V.; Berlec, A. Engineering of lactic acid bacteria for delivery of therapeutic proteins and peptides. Appl. Microbiol. Biotechnol. 2019, 103, 2053–2066. [Google Scholar] [CrossRef]
- Food and Drug Administration. Enforcement Policy Regarding Investigational New Drug Requirements for Use of Fecal Microbiota for Transplantation to Treat Clostridium difficile Infection Not Responsive to Standard Therapies. Center for Biologics Evaluation and Research. 2013. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/enforcement-policy-regarding-investigational-new-drug-requirements-use-fecal-microbiota (accessed on 18 November 2019).
- Bron, P.A.; Kleerebezem, M. Lactic acid bacteria for delivery of endogenous or engineered therapeutic molecules. Front. Microbiol. 2018, 9, 1821. [Google Scholar] [CrossRef]
- Research and Markets. Microbiome Therapeutics and Diagnostics Market (2nd Edition), 2017–2030. Available online: https://www.researchandmarkets.com/reports/4377904/microbiome-therapeutics-and-diagnostics-market (accessed on 19 November 2019).
- Custers, R. The regulatory status of gene-edited agricultural products in the EU and beyond. Emerg. Top. Life Sci. 2017, 1, 20170019. [Google Scholar]
- Callaway, E. CRISPR plants now subject to tough GM laws in European Union. Nature 2018, 560, 16. [Google Scholar] [CrossRef]
- Shew, A.M.; Nalley, L.L.; Snell, H.A.; Nayga, R.M.; Dixon, B.L. CRISPR versus GMOs: Public acceptance and valuation. Glob. Food Secur. 2018, 19, 71–80. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, H.; Zhu, H. CRISPR technology is revolutionizing the improvement of tomato and other fruit crops. Hortic. Res. 2019, 6, 77. [Google Scholar] [CrossRef] [Green Version]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plavec, T.V.; Berlec, A. Safety Aspects of Genetically Modified Lactic Acid Bacteria. Microorganisms 2020, 8, 297. https://doi.org/10.3390/microorganisms8020297
Plavec TV, Berlec A. Safety Aspects of Genetically Modified Lactic Acid Bacteria. Microorganisms. 2020; 8(2):297. https://doi.org/10.3390/microorganisms8020297
Chicago/Turabian StylePlavec, Tina Vida, and Aleš Berlec. 2020. "Safety Aspects of Genetically Modified Lactic Acid Bacteria" Microorganisms 8, no. 2: 297. https://doi.org/10.3390/microorganisms8020297
APA StylePlavec, T. V., & Berlec, A. (2020). Safety Aspects of Genetically Modified Lactic Acid Bacteria. Microorganisms, 8(2), 297. https://doi.org/10.3390/microorganisms8020297