Streptomyces lydicamycinicus sp. nov. and Its Secondary Metabolite Biosynthetic Gene Clusters for Polyketide and Nonribosomal Peptide Compounds

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Phenotypic and Chemotaxonomic Characterization
2.3. Phylogenetic Analysis Based on 16S rDNA Sequences
2.4. Genome Analysis
3. Results
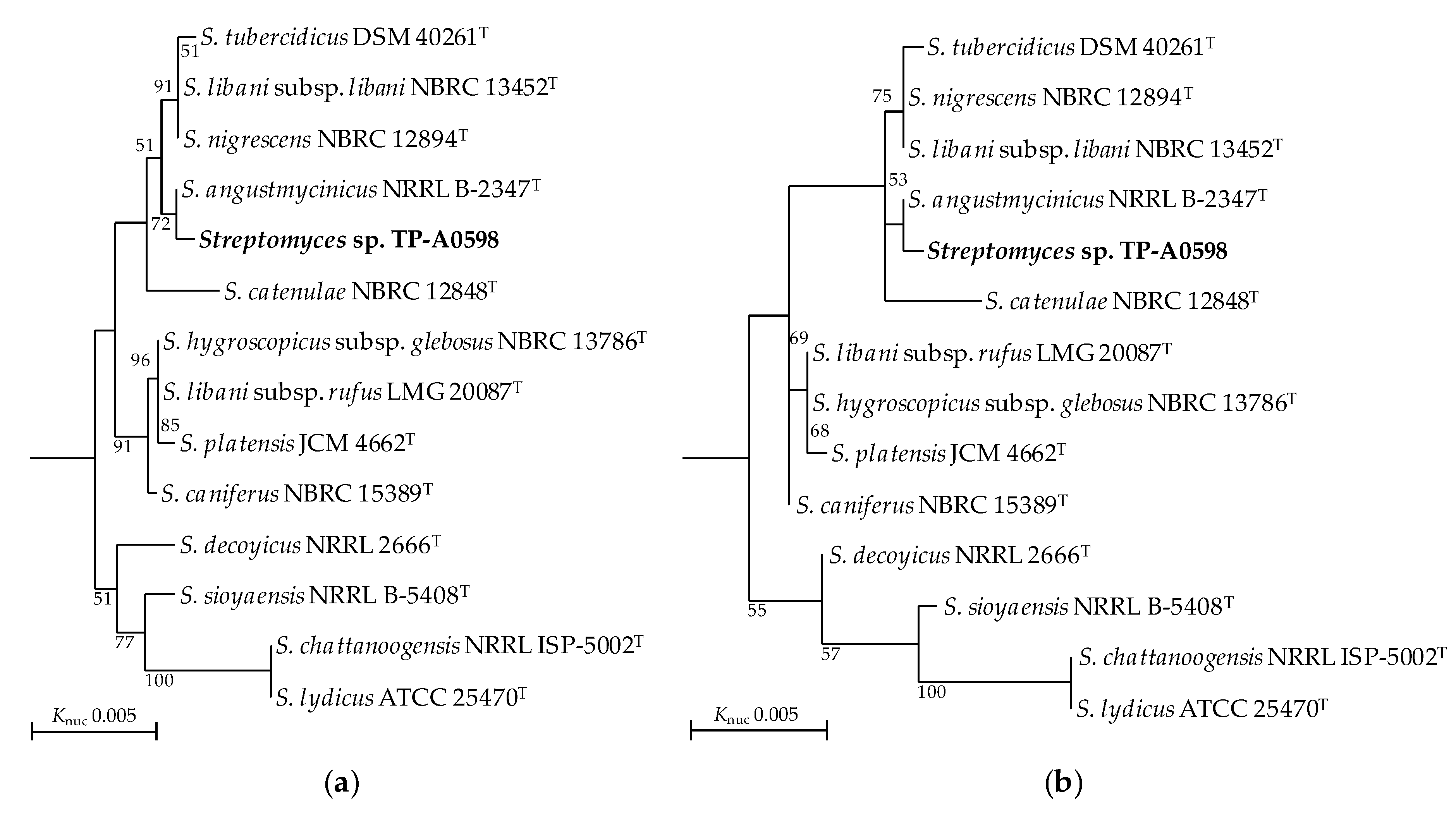
3.1. Classification of Streptomyces sp. TP-A0598
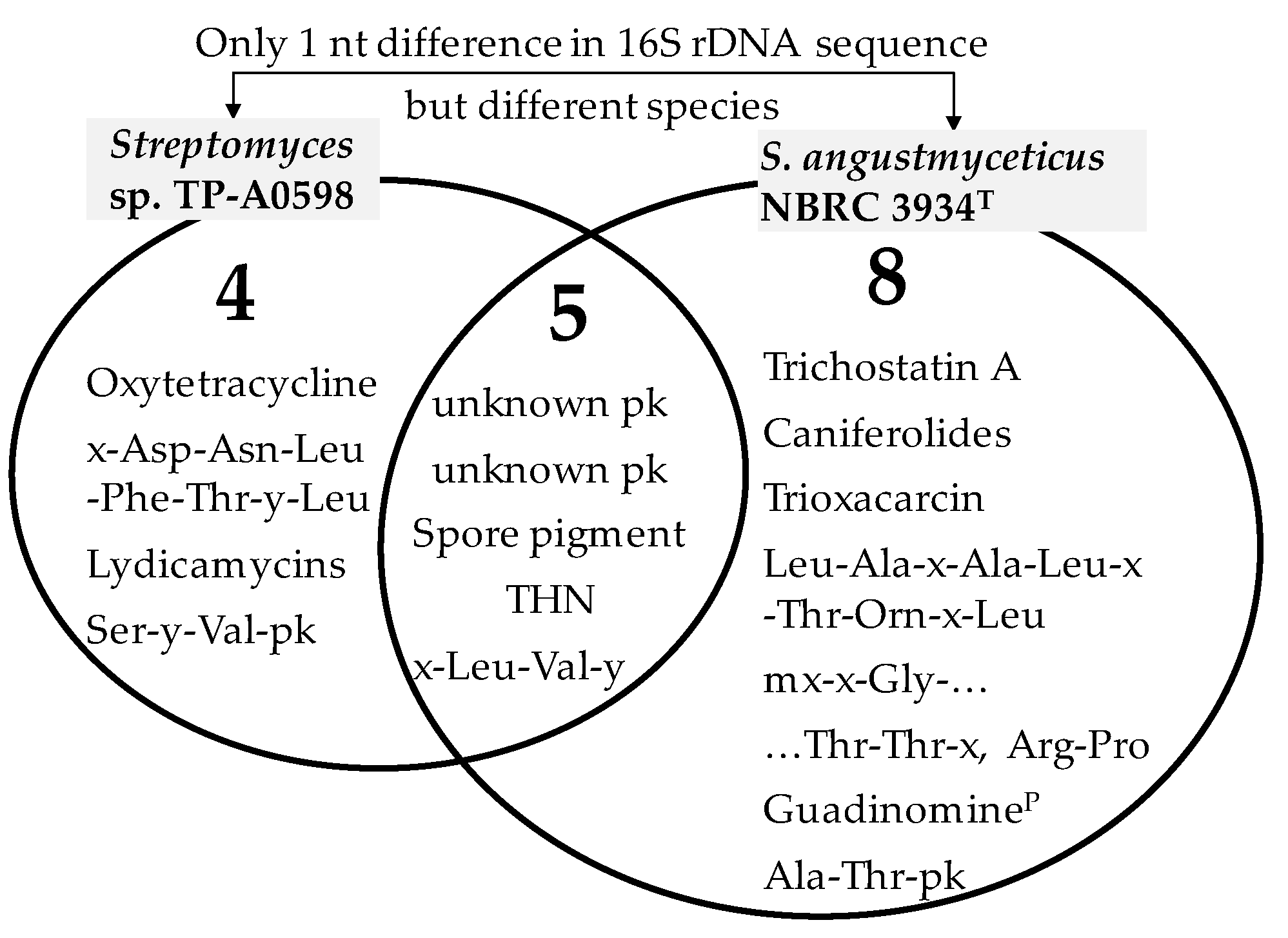
3.2. PKS and NRPS Gene Clusters of Streptomyces sp. TP-A0598 and S. angustmyceticus NBRC 3934T
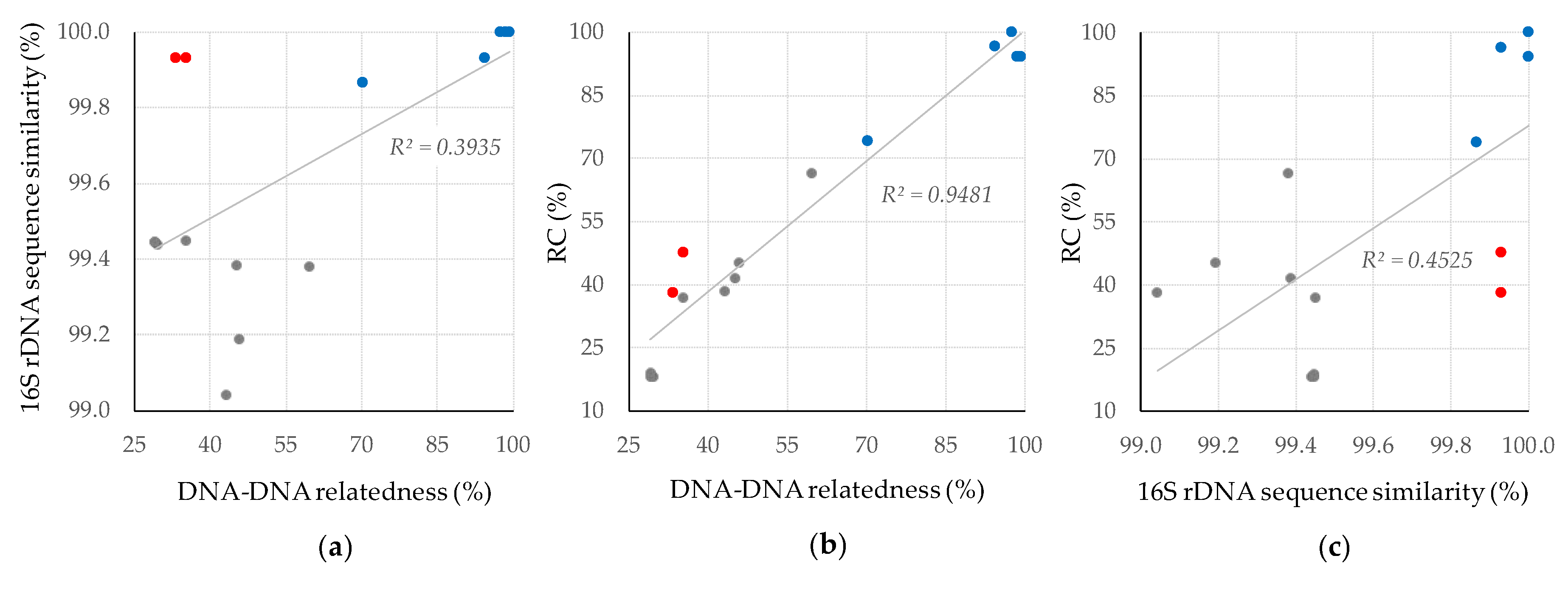
4. Discussion
5. Description of S. lydicamycinicus sp. nov.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Berdy, J. Bioactive microbial metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watve, M.G.; Tickoo, R.; Jog, M.M.; Bhole, B.D. How many antibiotics are produced by the genus Streptomyces? Arch. Microbiol. 2001, 176, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Nett, M.; Ikeda, H.; Moore, B.S. Genomic basis for natural product biosynthetic diversity in the actinomycetes. Nat. Prod. Rep. 2009, 26, 1362–1384. [Google Scholar] [CrossRef] [PubMed]
- Fischbach, M.A.; Walsh, C.T. Assembly-line enzymology for polyketide and nonribosomal peptide antibiotics: Logic, machinery, and mechanisms. Chem. Rev. 2006, 106, 3468–3496. [Google Scholar] [CrossRef] [PubMed]
- Furumai, T.; Eto, K.; Sasaki, T.; Higuchi, H.; Onaka, H. TPU-0037-A, B, C and D, novel lydicamycin congeners with anti-MRSA activity from Streptomyces platensis TP-A0598. J. Antibiot. 2002, 55, 873–880. [Google Scholar] [CrossRef] [Green Version]
- Komaki, H.; Ichikawa, N.; Hosoyama, A.; Fujita, N.; Igarashi, Y. Draft genome sequence of marine-derived Streptomyces sp. TP-A0598, a producer of anti-MRSA antibiotic lydicamycins. Stand. Genomic Sci. 2015, 10, 58. [Google Scholar] [CrossRef] [Green Version]
- Komaki, H.; Tamura, T. Reclassification of Streptomyces rimosus subsp. paromomycinus as Streptomyces paromomycinus sp. nov. Int. J. Syst. Evol. Microbiol. 2019, 69, 2577–2583. [Google Scholar] [CrossRef]
- Pridham, T.G.; Gottlieb, D. The utilization of carbon compounds by some Actinomycetales as an aid for species determination. J. Bacteriol. 1948, 56, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Goker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.H.; Ha, S.M.; Lim, J.; Kwon, S.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie van Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef]
- Komaki, H.; Sakurai, K.; Hosoyama, A.; Kimura, A.; Igarashi, Y. Diversity of nonribosomal peptide synthetase and polyketide synthase gene clusters among taxonomically close Streptomyces strains. Sci. Rep. 2018, 8, 6888. [Google Scholar] [CrossRef] [Green Version]
- Rong, X.; Huang, Y. Taxonomic evaluation of the Streptomyces hygroscopicus clade using multilocus sequence analysis and DNA-DNA hybridization, validating the MLSA scheme for systematics of the whole genus. Syst. Appl. Microbiol. 2012, 35, 7–18. [Google Scholar] [CrossRef]
- Wayne, L.G.; Brenner, D.J.; Colwell, R.R.; Grimont, P.A.D.; Kandler, O. Report of the ad hoc committee on reconciliation of approaches to bacterial systematics. Int. J. Syst. Bacteriol. 1987, 37, 463–464. [Google Scholar] [CrossRef] [Green Version]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [Green Version]
- Kudo, K.; Ozaki, T.; Shin-ya, K.; Nishiyama, M.; Kuzuyama, T. Biosynthetic origin of the hydroxamic acid moiety of trichostatin A: Identification of unprecedented enzymatic machinery involved in hydroxylamine transfer. J. Am. Chem. Soc. 2017, 139, 6799–6802. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Victoria, I.; Oves-Costales, D.; Lacret, R.; Martín, J.; Sánchez-Hidalgo, M.; Díaz, C.; Cautain, B.; Vicente, F.; Genilloud, O.; Reyes, F. Structure elucidation and biosynthetic gene cluster analysis of caniferolides A-D, new bioactive 36-membered macrolides from the marine-derived Streptomyces caniferus CA-271066. Org. Biomol. Chem. 2019, 17, 2954–2971. [Google Scholar] [CrossRef]
- Holmes, T.C.; May, A.E.; Zaleta-Rivera, K.; Ruby, J.G.; Skewes-Cox, P. Molecular insights into the biosynthesis of guadinomine: A type III secretion system inhibitor. J. Am. Chem. Soc. 2012, 134, 17797–17806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier-Kolthoff, J.P.; Göker, M.; Spröer, C.; Klenk, H.P. When should a DDH experiment be mandatory in icrobial taxonomy? Arch. Microbiol. 2013, 195, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Komaki, H.; Sakurai, K.; Hosoyama, A.; Kimura, A.; Trujilo, M.E. Diversity of PKS and NRPS gene clusters between Streptomyces abyssomicinicus sp. nov. and its taxonomic neighbor. J. Antibiot. 2020, 73, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Komaki, H.; Ichikawa, N.; Oguchi, A.; Tamura, T.; Hamada, M. Identification of the gene cluster for maridomycin/platenomycin and a taxonomic study of the producers by genome analysis. In The Annual Conference of Japan Society for Bioscience, Biotechnology, and Agrochemistry, Sapporo, Japan, March 29; Japan Society for Bioscience, Biotechnology, and Agrochemistry: Tokyo, Japan, 2016. [Google Scholar]
- Komaki, H.; Ichikawa, N.; Hosoyama, A.; Takahashi-Nakaguchi, A.; Matsuzawa, T. Genome based analysis of type-I polyketide synthase and nonribosomal peptide synthetase gene clusters in seven strains of five representative Nocardia species. BMC Genom. 2014, 15, 323. [Google Scholar] [CrossRef] [Green Version]
- Seipke, R.F. Strain-level diversity of secondary metabolism in Streptomyces albus. PLoS ONE 2015, 10, e0116457. [Google Scholar] [CrossRef] [Green Version]
- Metsä-Ketelä, M.; Halo, L.; Munukka, E.; Hakala, J.; Mantsala, P. Molecular evolution of aromatic polyketides and comparative sequence analysis of polyketide ketosynthase and 16S ribosomal DNA genes from various Streptomyces species. Appl. Environ. Microbiol. 2002, 68, 4472–4479. [Google Scholar] [CrossRef] [Green Version]
- Vicente, C.M.; Thibessard, A.; Lorenzi, J.N.; Benhadj, M.; Hôtel, L. Comparative genomics among closely related Streptomyces strains revealed specialized metabolite biosynthetic gene cluster diversity. Antibiotics (Basel) 2018, 7, 86. [Google Scholar] [CrossRef] [Green Version]
- Park, C.J.; Andam, C.P. Within-species genomic variation and variable patterns of recombination in the tetracycline producer Streptomyces rimosus. Front. Microbiol. 2019, 10, 552. [Google Scholar] [CrossRef]
- Antony-Babu, S.; Stien, D.; Eparvier, V.; Parrot, D.; Tomasi, S. Multiple Streptomyces species with distinct secondary metabolomes have identical 16S rRNA gene sequences. Sci. Rep. 2017, 7, 11089. [Google Scholar] [CrossRef]
- Choudoir, M.J.; Pepe-Ranney, C.; Buckley, D.H. Diversification of secondary metabolite biosynthetic gene clusters coincides with lineage divergence in Streptomyces. Antibiotics (Basel) 2018, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Chevrette, M.G.; Carlos-Shanley, C.; Louie, K.B.; Bowen, B.P.; Northen, T.R.; Currie, C.R. Taxonomic and metabolic incongruence in the ancient genus Streptomyces. Front. Microbiol. 2019, 10, 2170. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Med. | Streptomyces sp. TP-A0598 | Streptomyces angustmyceticus NBRC 3934T | |
|---|---|---|---|
| ISP 2 | SM | ++, Vivid yellow | ++, Moderate yellow to moderate yellowish brown |
| AM | +, Light bluish grey | +, Yellowish grey to dark reddish brown | |
| SP | Moderate reddish brown | Moderate reddish brown | |
| ISP 3 | SM | +, Moderate yellowish brown to moderate reddish brown | ±, Moderate yellowish brown |
| AM | +, Light yellowish brown to dark greyish red | +, Light olive grey to olive grey | |
| SP | Light to moderate yellowish brown | Moderate reddish brown | |
| ISP 4 | SM | ±, Vivid yellow | ++, Pale yellow or moderate yellow |
| AM | +, Light bluish grey | +, Light olive grey to dark reddish brown | |
| SP | – | Moderate yellowish brown | |
| ISP 5 | SM | +, Light yellow | ++, Moderate yellow |
| AM | +, White | +, Light bluish grey | |
| SP | – | Moderate yellowish brown | |
| ISP 6 | SM | +, Pale yellow | +, Pale yellow |
| AM | – | +, White | |
| SP | – | – | |
| ISP 7 | SM | +, Pale yellow to moderate yellow | ++, Moderate yellow |
| AM | +, White to light bluish grey | +, White to light bluish grey | |
| SP | Moderate yellow | Moderate brown | |
| 266 | SM | +, Moderate yellow to strong brown | ++, Moderate yellowish brown to moderate reddish brown |
| AM | +, Light bluish grey | +, Light bluish grey | |
| SP | Deep orange | Moderate brown | |
| 228 | SM | +, Strong brown | ++, Moderate yellowish brown to moderate reddish brown |
| AM | +, Light bluish grey to light brownish grey | +, White to greyish blue | |
| SP | Deep orange | Moderate brown |
| Characteristic | Streptomyces sp. TP-A0598 | S. angustmyceticus NBRC 3934T |
|---|---|---|
| Morphological | ||
| Aerial mycelium * | White to grey | White to black |
| Substrate mycelium * | Light yellow or vivid yellow | Moderate yellow |
| Soluble pigment | Yellow to Reddish brown | – |
| Chemotaxonomic | ||
| Major fatty acid (%) | C16:0 (23), iso-C15:0 (12), iso-C16:0 (12), anteiso-C15:0 (11) | iso-C16:0 (31), anteiso-C15:0 (12), iso-C15:0 (9) |
| Physiological | ||
| Growth at/with: | ||
| 15 °C | + | w |
| 37 °C | +++ | + |
| pH 5 | w | + |
| pH 10 | + | +++ |
| pH 11 | w | +++ |
| 7%–15% NaCl | – | ++ or + |
| Gelatin hydrolysis | + | – |
| Acid production from: | ||
| d-Adonitol | + | – |
| Amygdalin | w | – |
| Arbutin | w | – |
| Esculin ferric citrate | w | – |
| Gentiobiose | + | – |
| Gluconate | – | + |
| d-Maltose | – | + |
| Methyl-α-d-mannopyranoside | + | – |
| Salicin | w | – |
| d-Turanose | – | + |
| Xylitol | w | + |
| d-Xylose | – | w |
| Biochemical | ||
| N-Acetyl-β-glucosaminidase | w | + |
| Chymotrypsin | – | w |
| β-Galactosidase | + | w |
| Pyrrolidonyl arylamidase | w | + |
| Gene Cluster | Predicted Product | Locus Tag (TPA0598_) | Size (aa) | Domain Organization |
|---|---|---|---|---|
| t1pks-1 | Unknown | 10_00280 | 2436 | KS/AT/KR |
| 10_00270 | 1690 | KS/AT/DH/ACP | ||
| t1pks-2 | Unknown | 04_06320 | 444 | KS |
| 04_06310 | 2113 | KS/AT/DH/ER/KR/ACP | ||
| t2pks-1 | Spore pigment | 03_01500 | 422 | KSα |
| 03_01510 | 426 | KSβ (CLF) | ||
| 03_01520 | 89 | ACP | ||
| t2pks-2 (oxy) | Oxytetracycline | 07_00590 | 425 | KSα |
| 07_00600 | 426 | KSβ (CLF) | ||
| 07_00610 | 95 | ACP | ||
| t3pks-1 (rpp) | THN | 03_03810 | 353 | KS |
| nrps-1 | x-Leu-Val-y | 07_04820 | 1066 | C/A/T |
| 07_04810 | 641 | A(leu)/T | ||
| 07_04800 | 637 | A(val)/T | ||
| 07_04790 | 950 | C/T | ||
| nrps-2 | x-Asp-Asn-Leu-Phe-Thr-y-Leu | 02_01330 | 863 | C/A/T |
| 02_01450 | 2734 | A(asp)/T-C/A(asn)/T-C/A(leu)/T | ||
| 02_01460 | 2627 | C/A(phe)/T-C/A(thr)/T/E | ||
| 02_01470 | 1883 | C/T-C/A(leu)/T-TE | ||
| pks/nrps-1 | lydicamycin,TPU-0037-A to -D | 03_00740 | 3598 | KS/AT/DH/KR/ACP -KS/AT/DH/KR/ACP |
| 03_00750 | 7054 | KS/AT/DH/KR/ACP -KS/AT/DH/KR/ACP -KS/AT/DH/KR/ACP -KS/AT/DH/KR/ACP | ||
| 03_00760 | 3548 | KS/AT/DH/KR/ACP -KS/AT/DH/KR/ACP | ||
| 03_00770 | 1846 | KS/AT/DH/KR/ACP | ||
| 03_00780 | 5648 | KS/AT/DH/ER/KR/ACP -KS/AT/DH/KR/ACP -KS/AT/DH/KR/ACP | ||
| 03_00790 | 3662 | KS/AT/KR/ACP -KS/AT/DH/ER/KR/ACP | ||
| 03_00800 | 3265 | KS/AT/DH/KR/ACP -KS/AT/KR/ACP | ||
| 03_00820 | 1031 | C/A/T | ||
| 03_00840 | 1923 | ACP-KS/AT/DH/KR/ACP | ||
| pks/nrps-2 | Ser-y-Val-pk | 08_01960 | 556 | C/T |
| 08_01950 | 1139 | T-C/A(val)/T | ||
| 08_01940 | 1207 | KS/AT/ACP-TE | ||
| 08_01890 | 783 | A(ser)/T |
| Gene Cluster | Predicted Product | Locus Tag (San01_) | Size (aa) | Domain Organization |
|---|---|---|---|---|
| t1pks-1 | Unknown | 16600 | 2469 | KS/AT/KR |
| 16610 | 1601 | KS/AT/ACP | ||
| t1pks-2 | Unknown | 20810 | 428 | KS |
| 20820 | 2117 | KS/AT/DH/ER/KR/ACP | ||
| t1pks-3 (tsn) | Trichostatin A | s29-1 t,* | >375 | ACP/KS |
| RS35710 t | >471 | DH/ACP | ||
| RS35715 t | >897 | KS/AT | ||
| RS31690 t | >1500 | KS/AT/DH/KR/ACP | ||
| 64470 | 2005 | KS/AT/DH/KR/ACP-TE | ||
| t1pks-4 | Caniferolides | s04-1 t,* | >3397 | KS/AT/ACP -KS/AT/KR/ACP -KS/AT |
| RS35695 t | >990 | AT/KR/ACP | ||
| s40-2 t,* | >571 | KS | ||
| RS35360 t | >4323 | AT/KR/ACP -KS/AT/KR/ACP -KS/AT/DH/KR/ACP | ||
| 71810 | 3953 | KS/AT/DH/KR/ACP -KS/AT/DH/ER/KR/ACP | ||
| RS35370 t | >2135 | KS/AT/KR/ACP -KS | ||
| RS35705 t | >1815 | AT/DH/ER/DH/KR/ACP -KS | ||
| RS35690 t | >1167 | AT/KR/ACP | ||
| s39-1 t,* | >577 | KS | ||
| RS35595 t | >6115 | AT/KR/ACP -KS/AT/KR/ACP -KS/AT/DH/ACP -KS/AT/KR/ACP -KS | ||
| RS31970t | >1754 | AT/DH/ER/KR/ACP | ||
| 65020 | 5281 | KS/AT/KR/ACP -KS/AT/DH/ER/KR/ACP -KS/AT/KR/ACP | ||
| 65010 | 3643 | KS/AT/KR/ACP -KS/AT/DH/KR/ACP -TE | ||
| 64920 | 2404 | CoL/T-KS/AT/DH/KR/ACP | ||
| t2pks-1 | Spore pigment | 26680 | 422 | KSα |
| 26670 | 416 | KSβ (CLF) | ||
| 26660 | 96 | ACP | ||
| t2pks-3 | Trioxacarcin | 00550 | 421 | KSα |
| 00560 | 417 | KSβ (CLF) | ||
| 00570 | 89 | ACP | ||
| 00580 | 660 | AT | ||
| t3pks-1 (rpp) | THN | 24600 | 354 | KS |
| nrps-1 | x-Leu-Val-y | 06160 | 1066 | C/A/T |
| 06150 | 641 | A(leu)/T | ||
| 06140 | 637 | A(val)/T | ||
| 06130 | 950 | C/T | ||
| nrps-3 | Leu-Ala-x-Ala-Leu-x-Thr -Orn-x-Leu | 24170 | 6209 | A(leu)/T-C/A(ala)/T/E -C/A/T/E-C/A(ala)/T/E-C/A(leu)/T |
| 24160 | 6677 | C/A/T/E-C/A(thr)/T-C/A(orn)/T/E -C/A/T-C/A(leu)/T/E | ||
| nrps-4 | mx-x-Gly- | 36160 | 3675 | A/MT/T-C/A/T-C/A(gly)/T |
| s07-1 t,* | >398 | C | ||
| nrps-5 | Thr-Thr-x, Arg-Pro | 27520 t | >3401 | A(thr)/T-C/A(thr)/T-C/A/T-TE |
| 27580 | 591 | A(arg)/T | ||
| 27590 | 1,446 | C/A(pro)/T-TD | ||
| pks/nrps-3 | Guadinomine, partial | 61570 | 252 | KR |
| 61560 | 303 | AT | ||
| 61550 | 302 | AT | ||
| 61530 | 2393 | A(pip)/T-KS/AT/DH/KR/ACP | ||
| 61520 | 270 | TE | ||
| 61190 | 1178 | A/T-TD | ||
| pks/nrps-4 | Ala-Thr-pk | 12760 | 93 | ACP |
| 12770 | 674 | KS/DH | ||
| 12780 | 526 | A(ala) | ||
| 12800 | 1050 | A/T(thr)-C | ||
| 12810 | 274 | TE |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komaki, H.; Hosoyama, A.; Igarashi, Y.; Tamura, T. Streptomyces lydicamycinicus sp. nov. and Its Secondary Metabolite Biosynthetic Gene Clusters for Polyketide and Nonribosomal Peptide Compounds. Microorganisms 2020, 8, 370. https://doi.org/10.3390/microorganisms8030370
Komaki H, Hosoyama A, Igarashi Y, Tamura T. Streptomyces lydicamycinicus sp. nov. and Its Secondary Metabolite Biosynthetic Gene Clusters for Polyketide and Nonribosomal Peptide Compounds. Microorganisms. 2020; 8(3):370. https://doi.org/10.3390/microorganisms8030370
Chicago/Turabian StyleKomaki, Hisayuki, Akira Hosoyama, Yasuhiro Igarashi, and Tomohiko Tamura. 2020. "Streptomyces lydicamycinicus sp. nov. and Its Secondary Metabolite Biosynthetic Gene Clusters for Polyketide and Nonribosomal Peptide Compounds" Microorganisms 8, no. 3: 370. https://doi.org/10.3390/microorganisms8030370
APA StyleKomaki, H., Hosoyama, A., Igarashi, Y., & Tamura, T. (2020). Streptomyces lydicamycinicus sp. nov. and Its Secondary Metabolite Biosynthetic Gene Clusters for Polyketide and Nonribosomal Peptide Compounds. Microorganisms, 8(3), 370. https://doi.org/10.3390/microorganisms8030370