Isolation, Identification and Characterization of Endophytic Bacterium Rhizobium oryzihabitans sp. nov., from Rice Root with Biotechnological Potential in Agriculture

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Culture Conditions
2.2. Morphological, Physiological and Biochemical Characteristics
2.3. Molecular Studies
2.4. Fatty Acid Analysis
2.5. Analysis of Plant Growth-Promoting Characteristics
2.6. Comparative Genomics of Rhizobium Species
3. Results and Discussion
3.1. Morphological, Physiological and Biochemical Characteristic
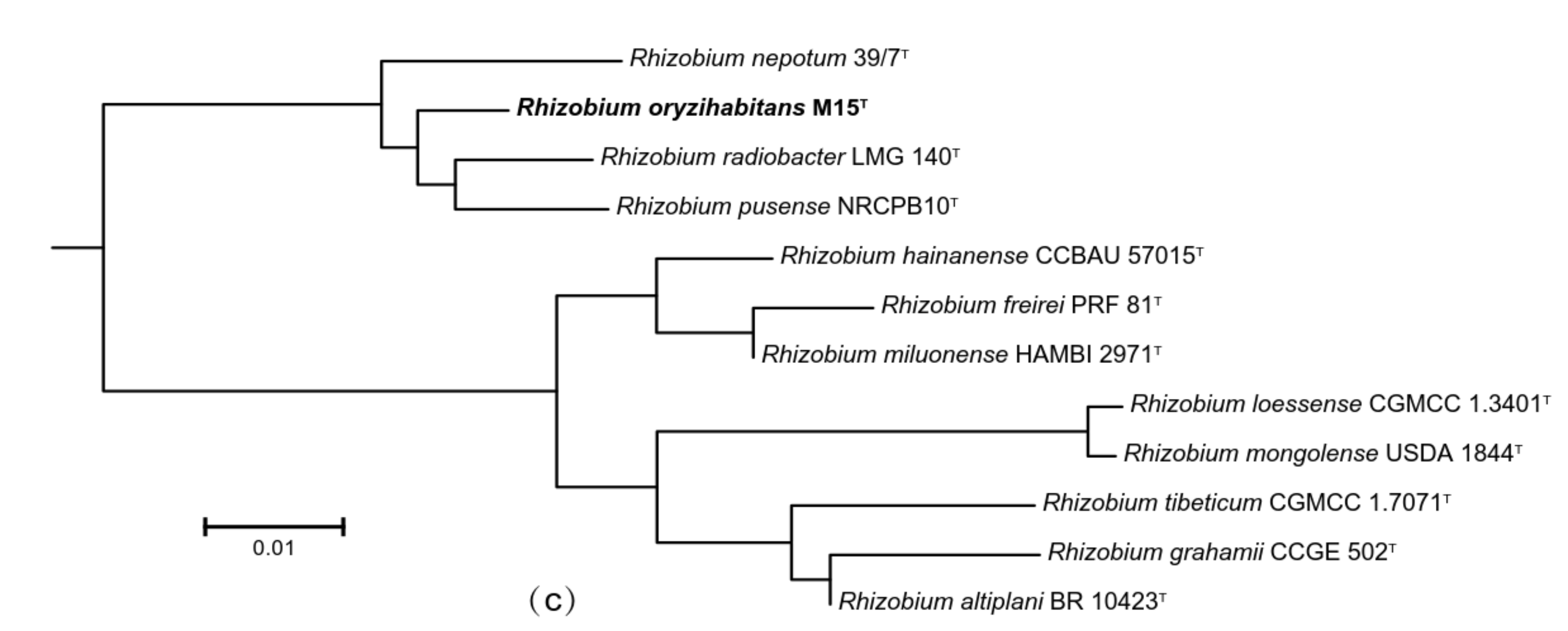
3.2. Molecular Studies
3.3. Cellular Fatty Acid Composition
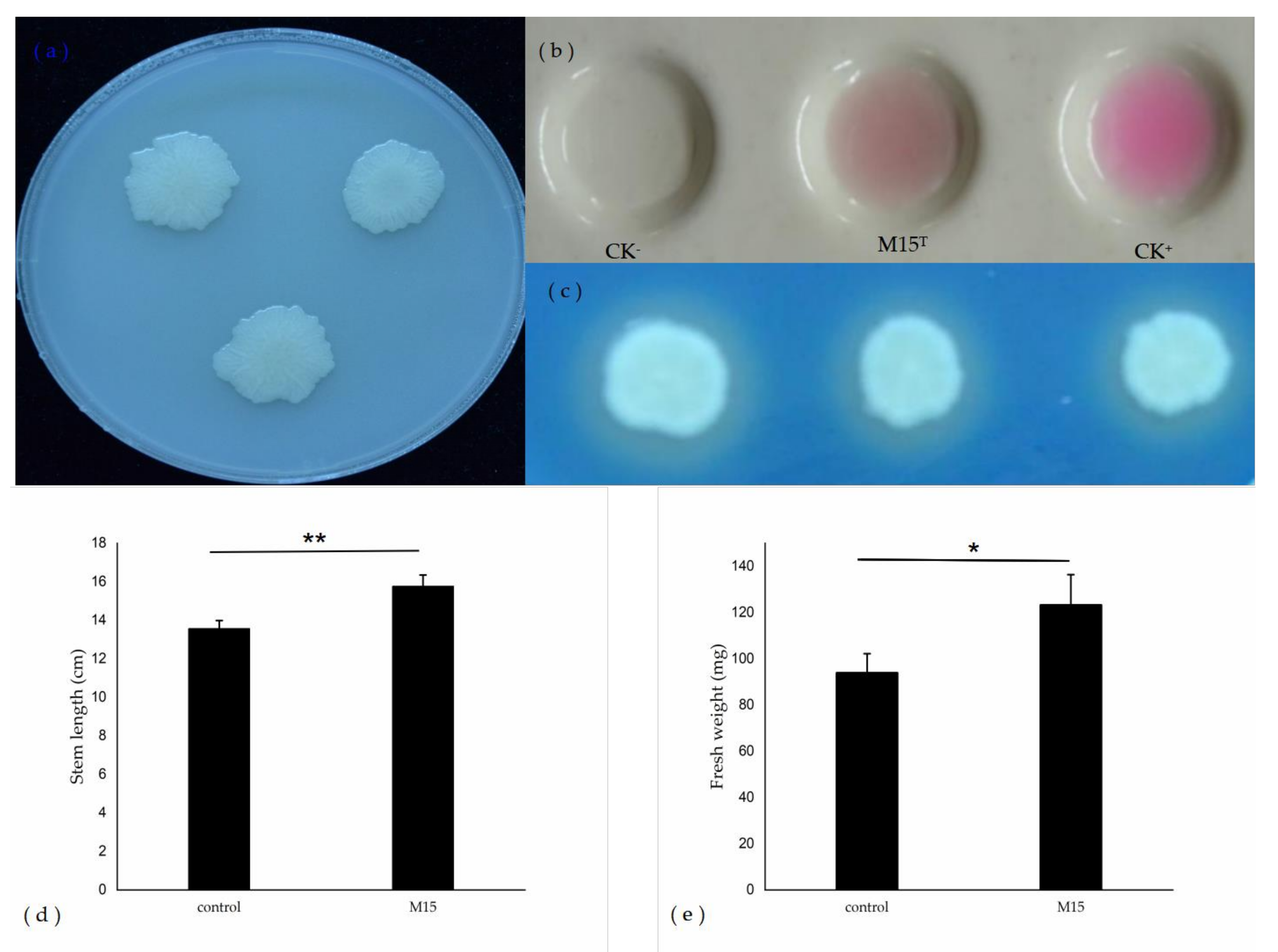
3.4. Analysis of Plant Growth-Promoting Characteristic
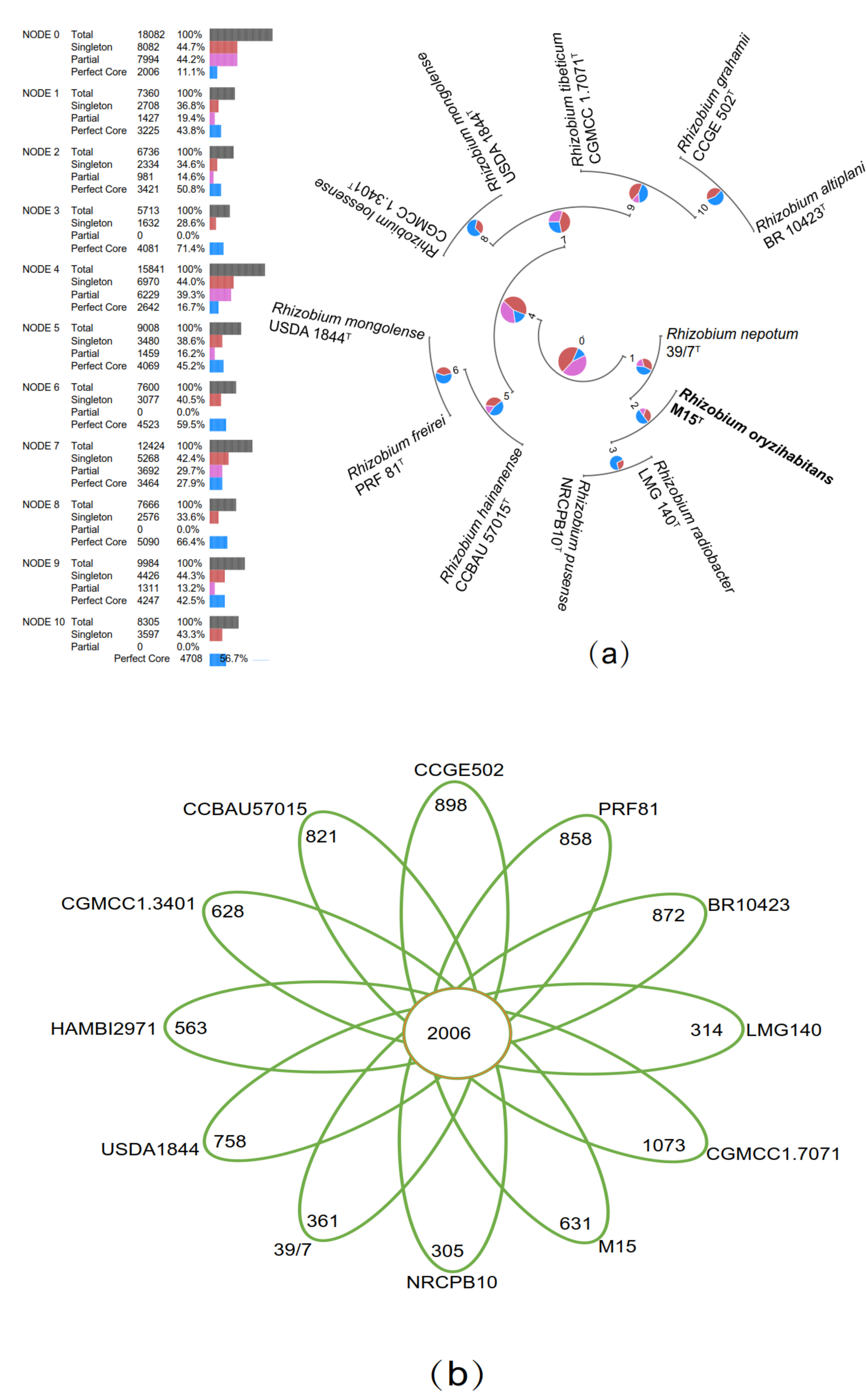
3.5. The Comparative Genome Analyses from 12 Closely Related Strains of the Genus Rhizobium
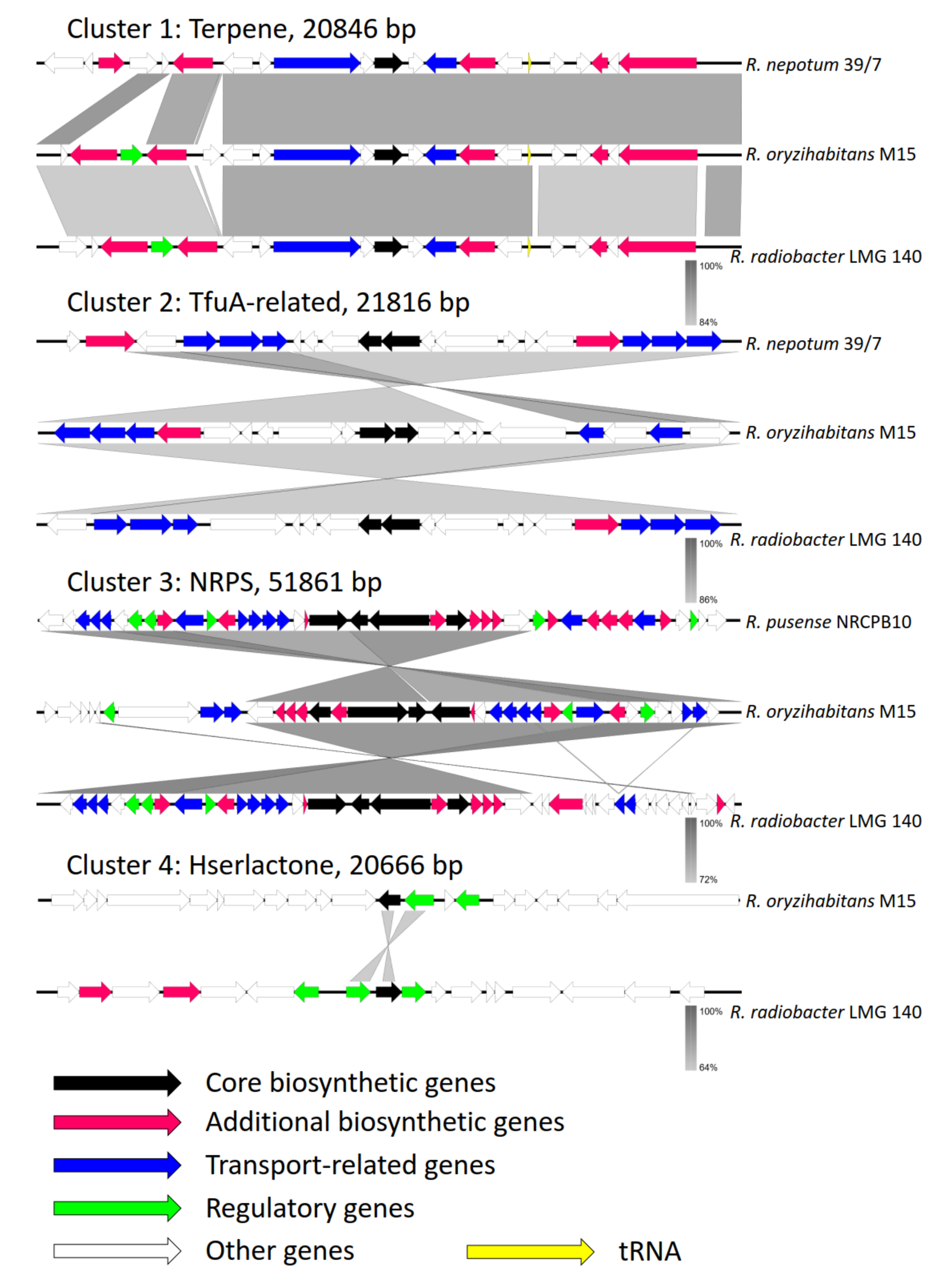
3.6. Secondary Metabolite Clusters
4. Conclusions
Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
References
- Frank, B. Ueber die pilzsymbiose der leguminosen. Ber. Deutsch. Bot. Ges. 1889, 7, 332–346. [Google Scholar]
- Behrendt, U.; Kämpfer, P.; Glaeser, S.P.; Augustin, J.; Ulrich, A. Characterization of the N2O-producing soil bacterium Rhizobium azooxidifex sp. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 2354–2361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, T.; Sun, L.N.; Zhang, J.; Sui, X.H.; Li, S.P. Rhizobium flavum sp. nov., a triazophos-degrading bacterium isolated from soil under the long-term application of triazophos. Int. J. Syst. Evol. Microbiol. 2014, 64, 2017–2022. [Google Scholar] [CrossRef]
- Zhang, X.; Li, B.; Wang, H.; Sui, X.; Ma, X.; Hong, Q.; Jiang, R. Rhizobium petrolearium sp. nov., isolated from oil-contaminated soil. Int. J. Syst. Evol. Microbiol. 2012, 62, 1871–1876. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.H.; Kang, S.J.; Yi, H.S.; Oh, T.K.; Ryu, C.M. Rhizobium soli sp. nov., isolated from soil. Int. J. Syst. Evol. Microbiol. 2010, 60, 1387–1393. [Google Scholar] [CrossRef] [Green Version]
- Sheu, S.Y.; Chen, Z.H.; Young, C.C.; Chen, W.M. Rhizobium ipomoeae sp. nov., isolated from a water convolvulus field. Int. J. Syst. Evol. Microbiol. 2016, 66, 1633–1640. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, R.P.; Ren, C.; Lai, Q.L.; Zeng, R.Y. Rhizobium marinum sp. nov., a malachite-green tolerant bacterium isolated from the sea water. Int. J. Syst. Evol. Microbiol. 2015, 65, 4449–4454. [Google Scholar] [CrossRef]
- Rosenblueth, M.; Martinez-Romero, E. Rhizobium etli maize populations and their competitiveness for root colonization. Arch. Microbiol. 2004, 181, 337–344. [Google Scholar] [CrossRef]
- Zhao, J.-J.; Zhang, X.; Sun, L.; Zhang, R.-J.; Zhang, C.-W.; Yin, H.-Q.; Zhang, X.-X. Rhizobium oryziradicis sp. nov. isolated from rice roots. Int. J. Syst. Evol. Microbiol. 2017, 67, 963–968. [Google Scholar] [CrossRef]
- Zhang, X.; Sun, L.; Ma, X.; Sui, X.H.; Jiang, R. Rhizobium pseudoryzae sp. nov. isolated from the rhizosphere of rice. Int. J. Syst. Evol. Microbiol. 2011, 61, 2425–2429. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-X.; Tang, X.; Sheirdil, R.A.; Sun, L.; Ma, X.-T. Rhizobium rhizoryzae sp. nov. isolated from rice roots. Int. J. Syst. Evol. Microbiol. 2014, 64, 1373–1377. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.X.; Gao, J.S.; Cao, Y.H.; Sheirdil, R.A.; Sheirdil, X.C.; Zhang, L. Isolation and Proposal Novel Rice Promoting Endophytic Bacteria, Rhizobium oryzicola sp. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 2931. [Google Scholar] [CrossRef]
- Pravin, V.; Rosazlin, A.; Tumirah, K.; Ismail, S.; Boyce, A.N. Role of Plant Growth Promoting Rhizobacteria in Agricultural Sustainability—A Review. Molecules 2016, 21, 573. [Google Scholar]
- Poole, P.; Ramachandran, V.; Terpolilli, J. Rhizobia: From saprophytes to endosymbionts. Nat. Rev. Microbiol. 2018, 16, 291–303. [Google Scholar] [CrossRef]
- Ferreira, C.M.H.; Soares, H.M.V.; Soares, E.V. Promising bacterial genera for agricultural practices: An insight on plant growth-promoting properties and microbial safety aspects. Sci. Total Environ. 2019, 682, 779–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Fraile, P.; Carro, L.; Robledo, M.; Bahena, M.H.R.; Flores-Félix, J.D.; Fernández, M.T.; Mateos, P.; Rivas, R.; Igual, J.M.; Martínez-Molina, E.; et al. Rhizobium promotes non-legumes growth and quality in several production steps: Towards a biofertilization of edible raw vegetables healthy for humans. PLoS ONE 2012, 7, e38122. [Google Scholar] [CrossRef] [Green Version]
- Vincent, J.M. The cultivation, isolation and maintenance of rhizobia. In A Manual for the Practical Study of the Root-Nodule Bacteria; Vincent, J.M., Ed.; Blackwell Scientific: Oxford, UK, 1970; pp. 1–13. [Google Scholar]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; Wiley: Chichester, UK, 1991; pp. 115–175. [Google Scholar]
- Martens, M.; Delaere, M.; Coopman, R.; De Vos, P.; Gillis, M.; Willems, A. Multilocus sequence analysis of Ensifer and related taxa. Int. J. Syst. Evol. Microbiol. 2007, 57, 489–503. [Google Scholar] [CrossRef] [Green Version]
- Martens, M.; Dawyndt, P.; Coopman, R.; Gillis, M.; De Vos, P.; Willems, A. Advantages of multilocus sequence analysis for taxonomic studies: A case study using 10 housekeeping genes in the genus Ensifer (including former Sinorhizobium). Int. J. Syst. Evol. Microbiol. 2008, 58, 200–214. [Google Scholar] [CrossRef] [Green Version]
- Kim, O.S.; Cho, Y.J.; Lee, K.; Yoon, S.H.; Kim, M.; Na, H.; Park, S.C.; Jeon, Y.S.; Lee, J.H.; Yi, H.; et al. Introducing EzTaxon-e: A prokaryotic 16S rRNA gene sequence database with phylotypes that represent uncultured species. Int. J. Syst. Evol. Microbiol. 2012, 62, 716–721. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTALW: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Boil. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Tritt, A.; Eisen, J.A.; Facciotti, M.T.; Darling, A.E. An Integrated Pipeline for de Novo Assembly of Microbial Genomes. PLoS ONE 2012, 7, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Chin, C.-S.; Peluso, P.; Sedlazeck, F.J.; Nattestad, M.; Concepcion, G.T.; Clum, A.; Dunn, C.; Omalley, R.; Figueroa-Balderas, R.; Morales-Cruz, A.; et al. Phased diploid genome assembly with single-molecule real-time sequencing. Nat. Methods 2016, 13, 1050–1054. [Google Scholar] [CrossRef] [Green Version]
- Sergey, K.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2012, 27, 722–736. [Google Scholar]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2012, 14, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sasser, M. Identification of Bacteria by Gas Chromatography of Cellular Fatty Acids; MIDI Inc.: Newark, DE, USA, 1990; MIDI Technical Note 101. [Google Scholar]
- Glickmann, E.; Dessaux, Y.A. Critical examination of the specificity of the Salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl. Environ. Microbiol. 1995, 619, 793–796. [Google Scholar] [CrossRef] [Green Version]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for detection and determination of siderophore. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Penrose, D.M.; Glick, B.R. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol. Plant. 2003, 118, 10–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habibi, S.; Djedidi, S.; Ohkama-Ohtsu, N.; Sarhadi, W.A.; Kojima, K.; Rallos, R.V.; Ramirez, M.D.A.; Yamaya, H.; Sekimoto, H.; Yokoyama, T. Isolation and Screening of Indigenous Plant Growth-promoting Rhizobacteria from Different Rice Cultivars in Afghanistan Soils. Microbes Environ. 2019, 34, 347–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arkin, A.P. KBase: The United States department of energy systems biology knowledgebase. Nat. Biotechnol. 2018, 36, 566–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2-approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Barton, I.S.; Fuqua, C.; Platt, T.G. Ecological and evolutionary dynamics of a model facultative pathogen: Agrobacterium and crown gall disease of plants. Environ Microbiol. 2018, 20, 16–29. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Jordan, D.C.; Genus, I. Rhizobium Frank 1889, 338AL. In Bergey’s Manual of Systematic Bacteriology; Krieg, N.R., Holt, J.G., Eds.; The Williams and Wilkins Co.: Baltimore, MD, USA, 1984; Volume 1, pp. 235–242. [Google Scholar]
- Kim, M.; Oh, H.S.; Park, S.C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [Green Version]
- Graham, P.H.; Sadowsky, M.J.; Keyser, H.H.; Barnet, Y.M.; Bradley, R.S. Proposed minimal standards for the description of new genera and species of root- and stem-nodulating bacteria. Int. J. Syst. Bacteriol. 1991, 41, 582–587. [Google Scholar] [CrossRef] [Green Version]
- Wayne, L.G.; Brenner, D.J.; Colwell, R.R.; Grimont, P.A.D.; Kandler, O. International Committee on Systematic Bacteriology. Report of the ad hoc committee on reconciliation of approaches to bacterial systematics. Int. J. Syst. Bacteriol. 1987, 37, 463–464. [Google Scholar] [CrossRef] [Green Version]
- Tighe, S.W.; De Lajudie, P.; DiPietro, K.; Lindström, K.; Nick, G.; Jarvis, B.D. Analysis of cellular fatty acids and phenotypic relationships of Agrobacterium, Bradyrhizobium, Mesorhizobium, Rhizobium and Sinorhizobium species using the Sherlock Microbial Identification System. Int. J. Syst. Evol. Microbiol. 2000, 50, 787–801. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.F.; Wei, J.Y.; Chen, H.W.; Liu, Y.Y.; Lu, H.Y.; Chou, J.Y. Indole-3-acetic acid: A widespread physiological code in interactions of fungi with other organisms. Plant Signal. Behav. 2015, 10, e1048052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, A.P.G.C.; Pires, C.; Moreira, H.; Ragel, A.O.S.S.; Castro, P.M.L. Assessment of the plant growth promotion abilities of six bacterial isolates using Zea mays as indicator plant. Soil Biol. Biochem. 2010, 42, 1229–1235. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; Kim, W.C. Plant Growth Promotion under Water: Decrease of Waterlogging-Induced ACC and Ethylene Levels by ACC Deaminase-Producing Bacteria. Front Microbiol. 2018, 9, 1096. [Google Scholar] [CrossRef] [PubMed]
- Tavares, M.J.; Nascimento, F.X.; Glick, B.R.; Rossi, M.J. The expression of an exogenous ACC deaminase by the endophyte Serratia grimesii BXF1 promotes the early nodulation and growth of common bean. Lett. Appl. Microbiol. 2018, 66, 252–259. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | 1 | 2 | 3 |
|---|---|---|---|
| Growth in/at | |||
| NaCl (7.0%) | + | - | - |
| pH 11 | + | - | + |
| Carbon-source utilization | |||
| Dextrin | - | w | w |
| N-acetyl-D-galactosamine | + | + | - |
| Lactulose | w | + | + |
| D-psicose | - | + | + |
| Methyl pyruvate | - | + | w |
| Mono-methyl-succinate | - | + | w |
| Formic acid | - | w | + |
| β-hydroxy butyric acid | - | + | w |
| α-keto glutaric acid | + | + | - |
| L-alanine | + | + | w |
| L-threonine | - | w | w |
| Inosine | w | - | - |
| Uridine | w | - | - |
| 2,3-Butanediol | - | - | w |
| Fatty Acid | 1 | 2 | 3 |
|---|---|---|---|
| C9:0 | 1.03 | - | - |
| C13:1 at 12–13 | 3.2 | 1.75 | 1.98 |
| C16:0 | 2.16 | 6.62 | 10.77 |
| C16:0 3-OH | 1.88 | 5.38 | 5.47 |
| C19:0 cycloω8c | 3.13 | 38.23 | 40.77 |
| Summed Feature 2 * | 38.54 | 11.93 | - |
| Summed Feature 3 * | 1.65 | 1.03 | 2.22 |
| Summed Feature 8 * | 40.99 | 26.54 | 30.94 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Zhao, X.; Wang, J.; Gong, Q.; Zhang, X.; Zhang, G. Isolation, Identification and Characterization of Endophytic Bacterium Rhizobium oryzihabitans sp. nov., from Rice Root with Biotechnological Potential in Agriculture. Microorganisms 2020, 8, 608. https://doi.org/10.3390/microorganisms8040608
Zhao J, Zhao X, Wang J, Gong Q, Zhang X, Zhang G. Isolation, Identification and Characterization of Endophytic Bacterium Rhizobium oryzihabitans sp. nov., from Rice Root with Biotechnological Potential in Agriculture. Microorganisms. 2020; 8(4):608. https://doi.org/10.3390/microorganisms8040608
Chicago/Turabian StyleZhao, Juanjuan, Xia Zhao, Junru Wang, Qi Gong, Xiaoxia Zhang, and Guishan Zhang. 2020. "Isolation, Identification and Characterization of Endophytic Bacterium Rhizobium oryzihabitans sp. nov., from Rice Root with Biotechnological Potential in Agriculture" Microorganisms 8, no. 4: 608. https://doi.org/10.3390/microorganisms8040608
APA StyleZhao, J., Zhao, X., Wang, J., Gong, Q., Zhang, X., & Zhang, G. (2020). Isolation, Identification and Characterization of Endophytic Bacterium Rhizobium oryzihabitans sp. nov., from Rice Root with Biotechnological Potential in Agriculture. Microorganisms, 8(4), 608. https://doi.org/10.3390/microorganisms8040608