Antifungal and Antibiofilm Activities and the Mechanism of Action of Repeating Lysine-Tryptophan Peptides against Candida albicans


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Peptide Synthesis
2.3. Antifungal Activity
2.4. Aggregation of KW5 Monitored by Thioflavin T (ThT) Fluorescence Assay
2.5. Fungicidal Action
2.6. Liposome Fusion Assay
2.7. Calcein LEAKAGE
2.8. Circular Dichroism (CD) Spectroscopy
2.9. Cytotoxicity
2.10. Biofilm Inhibition Assays
2.11. Cell Wall Binding
2.12. SYTOX Green
2.13. Flow Cytometry
2.14. Fluorescence Microscopy Analysis of Cell Permeabilization
2.15. Scanning Electron Microscopy
2.16. Tryptophan Fluorescence Spectroscopy and Acrylamide Quenching Assay
2.17. Nucleic Acid Binding
2.18. Confocal Laser-Scanning Microscopy (CLSM)
3. Results
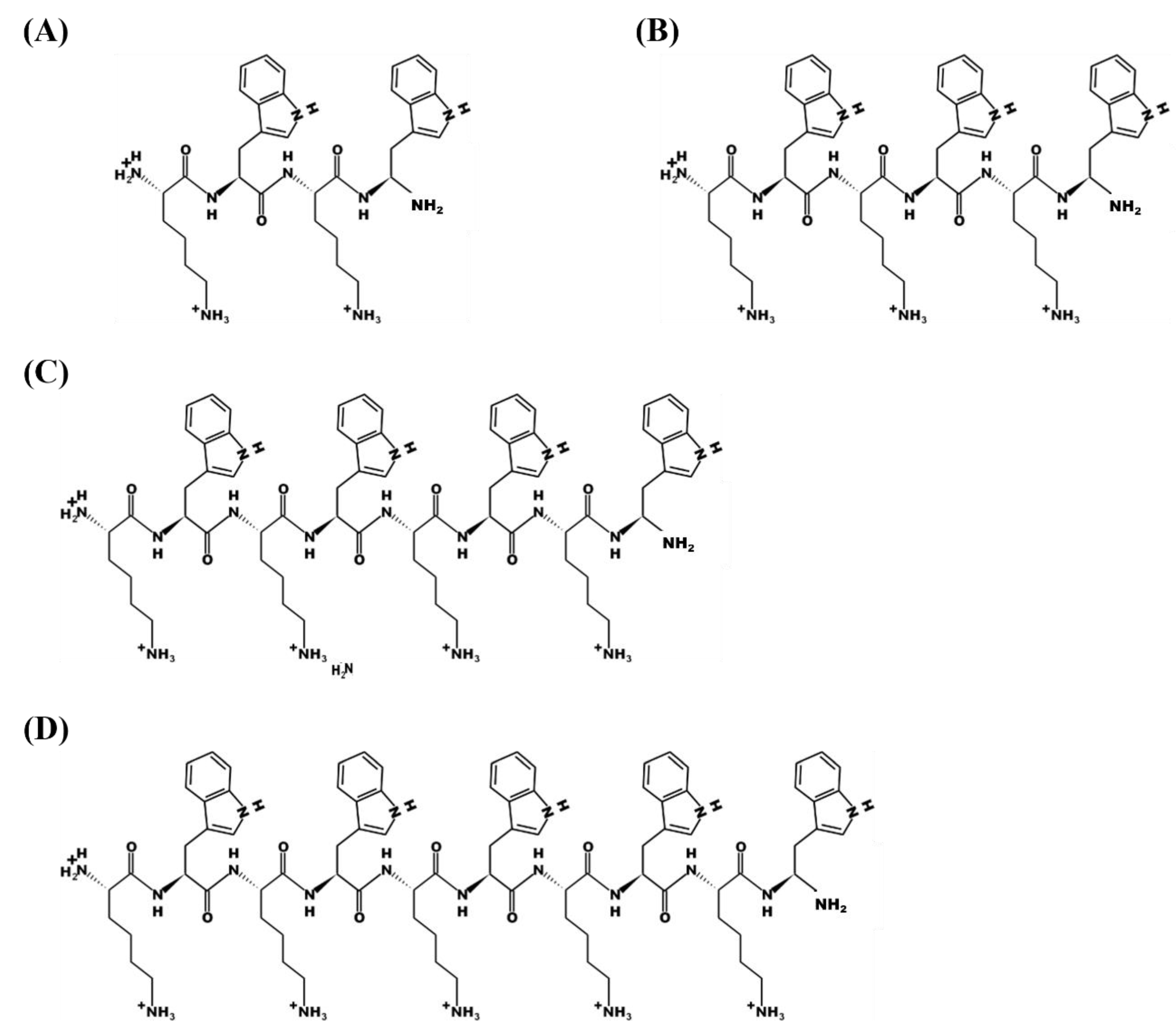
3.1. Effect of the Chain Length of the KWn Peptides on Their Antifungal Activity
3.2. Examining the Influence of Salt and pH on the KW5 Activity and Self-Association in Aqueous Solution
3.3. Fungicidal Activity of KWn Peptides in the Presence of Sodium Azide
3.4. Cytotoxicity of the KWn Peptides Against Mammalian Cells
3.5. Inhibitory Effect of KW4 on the Formation of C. albicans Biofilms
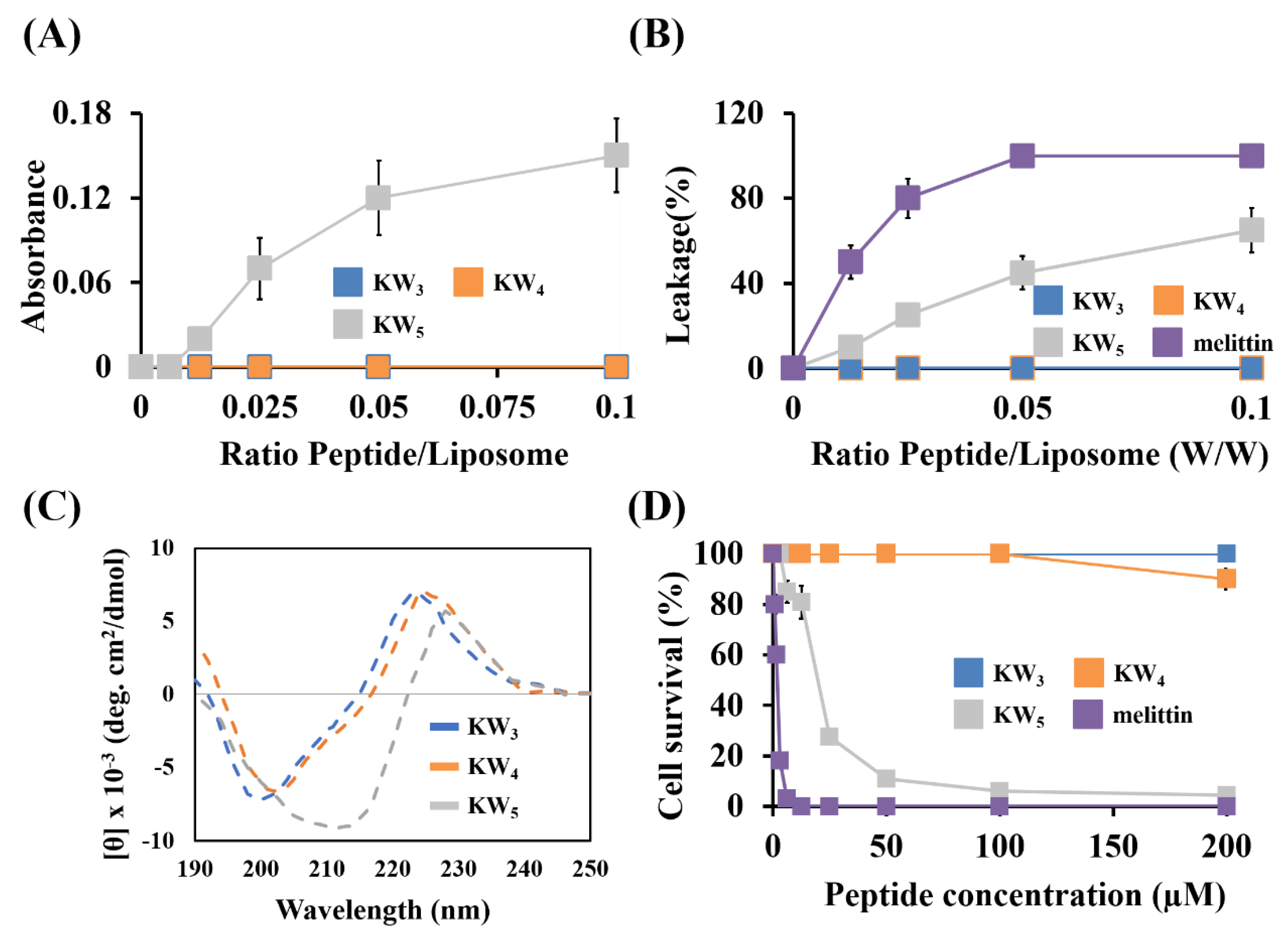
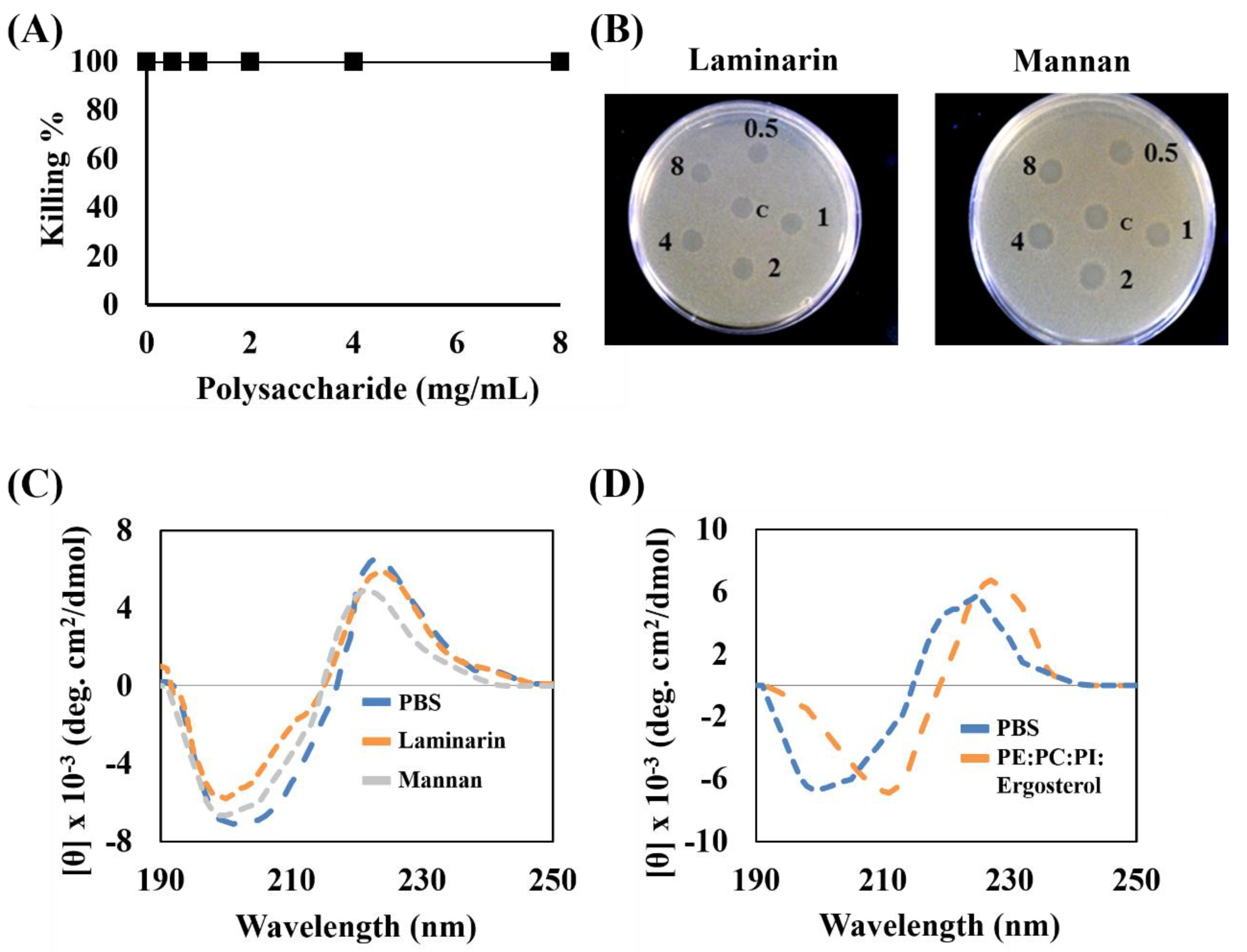
3.6. Interaction of Peptides with Fungal Cell Wall Components and Fungal Membranes
3.7. Non-Permeabilizing Action of the KWn Peptides in Fungal Membranes
3.8. Membrane Binding Action of Peptides
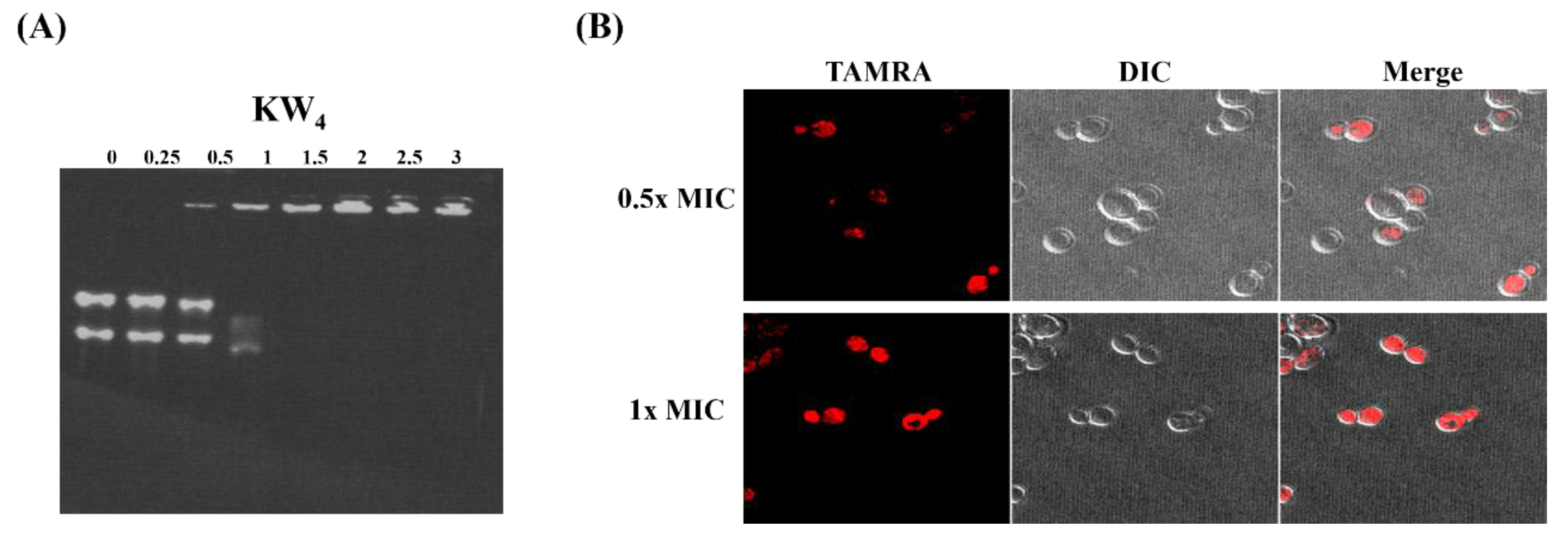
3.9. Nucleic Acid-Binding Properties of KW4 and Observation of Peptide Action by Confocal Microscopy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fisher, M.C.; Hawkins, N.J.; Sanglard, D.; Gurr, S.J. Worldwide emergence of resistance to antifungal drugs challenges human health and food security. Science 2018, 360, 739–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campoy, S.; Adrio, J.L. Antifungals. Biochem. Pharmacol. 2017, 133, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Nicola, A.M.; Albuquerque, P.; Paes, H.C.; Fernandes, L.; Costa, F.F.; Kioshima, E.S.; Abadio, A.K.R.; Bocca, A.L.; Felipe, M.S. Antifungal drugs: New insights in research & development. Pharmacol. Ther. 2019, 195, 21–38. [Google Scholar] [PubMed]
- Fuentefria, A.M.; Pippi, B.; Dalla Lana, D.F.; Donato, K.K.; de Andrade, S.F. Antifungals discovery: An insight into new strategies to combat antifungal resistance. Lett. Appl. Microbiol. 2018, 66, 2–13. [Google Scholar] [CrossRef] [Green Version]
- Lohse, M.B.; Gulati, M.; Johnson, A.D.; Nobile, C.J. Development and regulation of single-and multi-species candida albicans biofilms. Nat. Rev. Microbiol. 2018, 16, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Duncan, V.M.S.; O’Neil, D.A. Commercialization of antifungal peptides. Fungal Biol. Rev. 2013, 26, 156–165. [Google Scholar] [CrossRef]
- Ciociola, T.; Giovati, L.; Conti, S.; Magliani, W.; Santinoli, C.; Polonelli, L. Natural and synthetic peptides with antifungal activity. Future Med. Chem. 2016, 8, 1413–1433. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Haney, E.F.; Vogel, H.J. The expanding scope of antimicrobial peptide structures and their modes of action. Trends Biotechnol. 2011, 29, 464–472. [Google Scholar] [CrossRef]
- Hancock, R.E.W. Peptide antibiotics. Lancet 1997, 349, 418–422. [Google Scholar] [CrossRef]
- Hwang, P.M.; Vogel, H.J. Structure-function relationships of antimicrobial peptides. Biochem. Cell Biol. 1998, 76, 235–246. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.W.; Haney, E.F.; Gill, E.E. The immunology of host defense peptides: Beyond antimicrobial activity. Nat. Rev. Immunol. 2016, 16, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Haney, E.F.; Straus, S.K.; Hancock, R.E.W. Reassessing the host defense peptide landscape. Front. Chem. 2019, 7, 43–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hancock, R.E.; Sahl, H.G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Shai, Y.; Fox, J.; Caratsch, C.; Shih, Y.L.; Edwards, C.; Lazarovici, P. Sequencing and synthesis of pardaxin, a polypeptide from the red sea moses sole with ionophore activity. FEBS Lett. 1988, 242, 161–166. [Google Scholar] [CrossRef] [Green Version]
- Oren, Z.; Shai, Y. Selective lysis of bacteria but not mammalian cells by diastereomers of melittin: Structure−function study. Biochemistry 1997, 36, 1826–1835. [Google Scholar] [CrossRef] [PubMed]
- Johansson, J.; Gudmundsson, G.H.; Rottenberg, M.E.; Berndt, K.D.; Agerberth, B. Conformation-dependent antibacterial activity of the naturally occurring human peptide LL-37. J. Biol. Chem. 1998, 273, 3718–3724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, L.; Bai, X.; Luan, N.; Yao, H.; Zhang, Z.; Liu, W.; Chen, Y.; Yan, X.; Rong, M.; Lai, R.; et al. A designed tryptophan- and lysine/arginine-rich antimicrobial peptide with therapeutic potential for clinical antibiotic-resistant candida albicans vaginitis. J. Med. Chem. 2016, 59, 1791–1799. [Google Scholar] [CrossRef]
- Godballe, T.; Mojsoska, B.; Nielsen, H.M.; Jenssen, H. Antimicrobial activity of GN peptides and their mode of action. Biopolymers 2016, 106, 172–183. [Google Scholar] [CrossRef] [Green Version]
- Deslouches, B.; Hasek, M.L.; Craigo, J.K.; Steckbeck, J.D.; Montelaro, R.C. Comparative functional properties of engineered cationic antimicrobial peptides consisting exclusively of tryptophan and either lysine or arginine. J. Med. Microbiol. 2016, 65, 554–565. [Google Scholar] [CrossRef]
- Luong, H.X.; Kim, D.H.; Lee, B.J.; Kim, Y.W. Effects of lysine-to-arginine substitution on antimicrobial activity of cationic stapled heptapeptides. Arch. Pharm. Res. 2018, 41, 1092–1097. [Google Scholar] [CrossRef] [PubMed]
- Han, H.M.; Gopal, R.; Park, Y. Design and membrane-disruption mechanism of charge-enriched AMPs exhibiting cell selectivity, high-salt resistance, and anti-biofilm properties. Amino Acids 2016, 48, 505–522. [Google Scholar] [CrossRef] [PubMed]
- Koo, Y.S.; Kim, J.M.; Park, I.Y.; Yu, B.J.; Jang, S.A.; Kim, K.S.; Park, C.B.; Cho, J.H.; Kim, S.C. Structure–activity relations of parasin I, a histone H2A-derived antimicrobial peptide. Peptides 2008, 29, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Rothstein, D.M.; Spacciapoli, P.; Tran, L.T.; Xu, T.; Roberts, F.D.; Dalla Serra, M.; Buxton, D.K.; Oppenheim, F.G.; Friden, P. Anticandida activity is retained in P-113, a 12-amino-acid fragment of histatin 5. Antimicrob. Agents Chemother. 2001, 45, 1367–1373. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.F.; Soto, A.; Knoop, F.C.; Conlon, J.M. Antimicrobial peptides isolated from skin secretions of the diploid frog, Xenopus tropicalis (Pipidae). Biochim. Biophys. Acta 2001, 1550, 81–89. [Google Scholar] [CrossRef]
- Shen, W.-C.; Ryser, H. Conjugation of poly-L-lysine to albumin and horseradish peroxidase: A novel method of enhancing the cellular uptake of proteins. Proc. Natl. Acad. Sci. USA 1978, 75, 1872–1876. [Google Scholar] [CrossRef] [Green Version]
- Hyun, S.; Kim, H.J.; Lee, N.J.; Lee, K.H.; Lee, Y.; Ahn, D.R.; Kim, K.; Jeong, S.; Yu, J. Alpha-helical peptide containing N,N-dimethyl lysine residues displays low-nanomolar and highly specific binding to RRE RNA. J. Am. Chem. Soc. 2007, 129, 4514–4515. [Google Scholar] [CrossRef]
- Strøm, M.B.; Haug, B.E.; Rekdal, O.; Skar, M.L.; Stensen, W.; Svendsen, J.S. Important structural features of 15-residue lactoferricin derivatives and methods for improvement of antimicrobial activity. Biochem. Cell Biol. 2002, 80, 65–74. [Google Scholar] [CrossRef]
- Andrushchenko, V.V.; Vogel, H.J.; Prenner, E.J. Interactions of tryptophan-rich cathelicidin antimicrobial peptides with model membranes studied by differential scanning calorimetry. Biochim. Biophys. Acta 2007, 1768, 2447–2458. [Google Scholar] [CrossRef] [Green Version]
- Subbalakshmi, C.; Bikshapathy, E.; Sitaram, N.; Nagaraj, R. Antibacterial and hemolytic activities of single tryptophan analogs of indolicidin. Biochem. Biophys. Res. Commun. 2000, 274, 714–716. [Google Scholar] [CrossRef]
- Strøm, M.B.; Haug, B.E.; Skar, M.L.; Stensen, W.; Stiberg, T.; Svendsen, J.S. The pharmacophore of short cationic antibacterial peptides. J. Med. Chem. 2003, 46, 1567–1570. [Google Scholar] [CrossRef] [PubMed]
- Dathe, M.; Nikolenko, H.; Klose, J.; Bienert, M. Cyclization increases the antimicrobial activity and selectivity of arginine-and tryptophan-containing hexapeptides. Biochemistry 2004, 43, 9140–9150. [Google Scholar] [CrossRef] [PubMed]
- Houghten, R.A.; Pinilla, C.; Blondelle, S.E.; Appel, J.R.; Dooley, C.T.; Cuervo, J.H. Generation and use of synthetic peptide combinatorial libraries for basic research and drug discovery. Nature 1991, 354, 84–86. [Google Scholar] [CrossRef] [PubMed]
- Hunter, H.N.; Jing, W.; Schibli, D.J.; Trinh, T.; Park, I.Y.; Kim, S.C.; Vogel, H.J. The interactions of antimicrobial peptides derived from lysozyme with model membrane systems. Biochim. Biophys. Acta 2005, 1668, 175–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gopal, R.; Seo, C.H.; Song, P.I.; Park, Y. Effect of repetitive lysine–tryptophan motifs on the bactericidal activity of antimicrobial peptides. Amino Acids 2013, 44, 645–660. [Google Scholar] [CrossRef] [Green Version]
- Pouny, Y.; Shai, Y. Interaction of D-amino acid incorporated analogs of pardaxin with membranes. Biochemistry 1992, 31, 9482–9490. [Google Scholar] [CrossRef]
- Park, S.C.; Lee, J.R.; Shin, S.O.; Park, Y.; Lee, S.Y.; Hahm, K.S. Characterization of a heat-stable protein with antimicrobial activity from arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2007, 362, 562–567. [Google Scholar] [CrossRef]
- Jang, W.S.; Bajwa, J.S.; Sun, J.N.; Edgerton, M. Salivary histatin 5 internalization by translocation, but not endocytosis, is required for fungicidal activity in Candida albicans. Mol. Microbiol. 2010, 77, 354–370. [Google Scholar] [CrossRef] [Green Version]
- Veerman, E.C.; Valentijn-Benz, M.; Nazmi, K.; Ruissen, A.L.; Walgreen-Weterings, E.; van Marle, J.; Doust, A.B.; van’t Hof, W.; Bolscher, J.G.; Amerongen, A.V. Energy depletion protects Candida albicans against antimicrobial peptides by rigidifying its cell membrane. J. Biol. Chem. 2007, 282, 18831–18841. [Google Scholar] [CrossRef] [Green Version]
- Park, S.C.; Kim, J.Y.; Shin, S.O.; Jeong, C.Y.; Kim, M.H.; Shin, S.Y.; Cheong, G.W.; Park, Y.; Hahm, K.S. Investigation of toroidal pore and oligomerization by melittin using transmission electron microscopy. Biochem. Biophys. Res. Commun. 2006, 343, 222–228. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Sugishita, K.; Miyajima, K. Interactions of an antimicrobial peptide, magainin 2, with lipopolysaccharide-containing liposomes as a model for outer membranes of gram-negative bacteria. FEBS Lett. 1999, 449, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Kang, H.K.; Choi, M.C.; Chae, J.D.; Son, B.K.; Chong, Y.P.; Seo, C.H.; Park, Y. Antibacterial activity and mechanism of action of analogues derived from the antimicrobial peptide mBjAMP1 isolated from Branchiostoma japonicum. J. Antimicrob. Chemother. 2018, 73, 2054–2063. [Google Scholar] [CrossRef] [PubMed]
- Rajasekharan, S.K.; Kamalanathan, C.; Ravichandran, V.; Ray, A.K.; Satish, A.S.; Mohanvel, S.K. Mannich base limits Candida albicans virulence by inactivating Ras-cAMP-PKA pathway. Sci. Rep. 2018, 8, 14972. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.S.; Kim, H.K.; Lee, K.Y.; Kim, S.A.; Han, Y.S.; Lee, I.H. Antifungal activity of synthetic peptide derived from halocidin, antimicrobial peptide from the tunicate, Halocynthia aurantium. FEBS Lett. 2006, 580, 1490–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewan, P.C.; Anantharaman, A.; Chauhan, V.S.; Sahal, D. Antimicrobial action of prototypic amphipathic cationic decapeptides and their branched dimers. Biochemistry 2009, 48, 5642–5657. [Google Scholar] [CrossRef]
- Mao, D.; Wallace, B.A. Differential light scattering and absorption flattening optical effects are minimal in the circular dichroism spectra of small unilamellar vesicles. Biochemistry 1984, 23, 2667–2673. [Google Scholar] [CrossRef]
- Sweeney, T.; Collins, C.B.; Reilly, P.; Pierce, K.M.; Ryan, M.; O’Doherty, J.V. Effect of purified β-glucans derived from Laminaria digitata, Laminaria hyperborea and Saccharomyces cerevisiae on piglet performance, selected bacterial populations, volatile fatty acids and pro-inflammatory cytokines in the gastrointestinal tract of pigs. Br. J. Nutr. 2012, 108, 1226–1234. [Google Scholar] [CrossRef] [Green Version]
- Woody, R.W. Contributions of tryptophan side chains to the far-ultraviolet circular dichroism of proteins. Eur. Biophys. J. 1994, 23, 253–262. [Google Scholar] [CrossRef]
- Lee, W.; Lee, D.G. Fungicidal mechanisms of the antimicrobial peptide Bac8c. Biochim. Biophys. Acta 2015, 1848, 673–679. [Google Scholar] [CrossRef] [Green Version]
- Burstein, E.A.; Vedenkina, N.S.; Ivkova, M.N. Fluorescence and the location of tryptophan residues in protein molecules. Photochem. Photobiol. 1973, 18, 263–279. [Google Scholar] [CrossRef]
- Liu, Z.; Brady, A.; Young, A.; Rasimick, B.; Chen, K.; Zhou, C.; Kallenbach, N.R. Length effects in antimicrobial peptides of the (RW)n series. Antimicrob. Agents Chemother. 2007, 51, 597–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gopal, R.; Na, H.; Seo, C.H.; Park, Y. Antifungal activity of (KW)n or (RW)n peptide against Fusarium solani and Fusarium oxysporum. Int. J. Mol. Sci. 2012, 13, 15042–15053. [Google Scholar] [CrossRef] [PubMed]
- Deslouches, B.; Phadke, S.M.; Lazarevic, V.; Cascio, M.; Islam, K.; Montelaro, R.C.; Mietzner, T.A. De novo generation of cationic antimicrobial peptides: Influence of length and tryptophan substitution on antimicrobial activity. Antimicrob. Agents Chemother. 2005, 49, 316–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, S.H.; Wimley, W.C. Hydrophobic interactions of peptides with membrane interfaces. Biochim. Biocphys. Acta 1998, 1376, 339–352. [Google Scholar] [CrossRef] [Green Version]
- Makovitzki, A.; Shai, Y. pH-dependent antifungal lipopeptides and their plausible mode of action. Biochemistry 2005, 44, 9775–9784. [Google Scholar] [CrossRef]
- Krishnakumari, V.; Rangaraj, N.; Nagaraj, R. Antifungal activities of human beta-defensins HBD-1 to HBD-3 and their c-terminal analogs Phd1 to Phd3. Antimicrob. Agents Chemother. 2009, 53, 256–260. [Google Scholar] [CrossRef] [Green Version]
- Ibeas, J.I.; Lee, H.; Damsz, B.; Prasad, D.T.; Pardo, J.M.; Hasegawa, P.M.; Bressan, R.A.; Narasimhan, M.L. Fungal cell wall phosphomannans facilitate the toxic activity of a plant PR-5 protein. Plant J. 2000, 23, 375–383. [Google Scholar] [CrossRef]
- van der Weerden, N.L.; Hancock, R.E.; Anderson, M.A. Permeabilization of fungal hyphae by the plant defensin NaD1 occurs through a cell wall-dependent process. J. Biol. Chem. 2010, 285, 37513–37520. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Park, C.; Park, S.C.; Woo, E.R.; Park, Y.; Hahm, K.S.; Lee, D.G. Cell selectivity-membrane phospholipids relationship of the antimicrobial effects shown by pleurocidin enantiomeric peptides. J. Pept. Sci. 2009, 15, 601–606. [Google Scholar] [CrossRef]
- Mano, M.; Henriques, A.; Paiva, A.; Prieto, M.; Gavilanes, F.; Simões, S.; Pedroso de Lima, M.C. Cellular uptake of S413-PV peptide occurs upon conformational changes induced by peptide–membrane interactions. Biochim. Biophys. Acta 2006, 1758, 336–346. [Google Scholar] [CrossRef] [Green Version]
- Henriques, S.T.; Melo, M.N.; Castanho, M.A. Cell-penetrating peptides and antimicrobial peptides: How different are they? Biochem. J. 2006, 399, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mano, M.; Henriques, A.; Paiva, A.; Prieto, M.; Gavilanes, F.; Simões, S.; Pedroso de Lima, M.C. Interaction of S413-PV cell penetrating peptide with model membranes: Relevance to peptide translocation across biological membranes. J. Pept. Sci. 2007, 13, 301–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuzaki, K. Membrane permeabilization mechanisms. In Antimicrobial Peptides; Springer: Singapore, 2019; pp. 9–16. [Google Scholar]
- Lee, J.K.; Park, S.C.; Hahm, K.S.; Park, Y. Antimicrobial HPA3NT3 peptide analogs: Placement of aromatic rings and positive charges are key determinants for cell selectivity and mechanism of action. Biochim. Biophys. Acta 2013, 1828, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haney, E.F.; Petersen, A.P.; Lau, C.K.; Jing, W.; Storey, D.G.; Vogel, H.J. Mechanism of action of puroindoline derived tryptophan-rich antimicrobial peptides. Biochim. Biophys. Acta 2013, 1828, 1802–1813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernst, A.M.; Contreras, F.X.; Brügger, B.; Wieland, F. Determinants of specificity at the protein-lipid interface in membranes. FEBS Lett. 2010, 584, 1713–1720. [Google Scholar] [CrossRef] [Green Version]
- Muller, M.P.; Jiang, T.; Sun, C.; Lihan, M.; Pant, S.; Mahinthichaichan, P.; Trifan, A.; Tajkhorshid, E. Characterization of lipid-protein interactions and lipid-mediated modulation of membrane protein function through molecular simulation. Chem. Rev. 2019, 119, 6086–6161. [Google Scholar] [CrossRef]
- de Jesus, A.J.; Allen, T.W. The role of tryptophan side chains in membrane protein anchoring and hydrophobic mismatch. Biochim. Biophys. Acta 2013, 1828, 864–876. [Google Scholar] [CrossRef] [Green Version]
- Shagaghi, N.; Palombo, E.A.; Clayton, A.H.; Bhave, M. Archetypal tryptophan-rich antimicrobial peptides: Properties and applications. World J. Microbiol. Biotechnol. 2016, 32, 31–40. [Google Scholar] [CrossRef]
- Rezansoff, A.J.; Hunter, H.N.; Jing, W.; Park, I.Y.; Kim, S.C.; Vogel, H.J. Interactions of the antimicrobial peptide Ac-FRWWHR-NH(2) with model membrane systems and bacterial cells. J. Pept. Res. 2005, 65, 491–501. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Minimum Inhibitory Concentrations (µM) | ||||||
|---|---|---|---|---|---|---|
| Strain | KW2 | KW3 | KW4 | KW5 | Melittin | Fluconazole |
| C. albicans (KCTC 7270) | >128 | 32 | 8 | 8 | 4 | 16 |
| Resistant strains | ||||||
| C. albicans (CCARM 14001) | >128 | >128 | 32 | 8 | 16 | >128 |
| C. albicans (CCARM 14007) | >128 | >128 | 32 | 16 | 8 | >128 |
| C. albicans (CCARM 140020) | >128 | >128 | 32 | 8 | 8 | >128 |
| Cytotoxicty | ||||||
| Cell survival a (%) | 100 | 100 | 91 | 4 | 0 | - |
| Hemolysis b (%) | 0 | 0 | 8 | 71 | 100 | - |
| Peptide λmax Buffer (nm) | Blue Shift (nm) | KSV (M−1) a | ||||
|---|---|---|---|---|---|---|
| PE–PC–PI–Ergosterol | PC–CH–SM | Buffer | PE–PC–PI–Ergosterol | PC–CH–SM | ||
| KW3 | 353 | 6 | 1 | 15 | 2.8 | 5.7 |
| KW4 | 353 | 8 | 1 | 14 | 2.2 | 5.2 |
| KW5 | 351 | 10 | 6 | 11 | 2.0 | 2.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramamourthy, G.; Park, J.; Seo, C.; J. Vogel, H.; Park, Y. Antifungal and Antibiofilm Activities and the Mechanism of Action of Repeating Lysine-Tryptophan Peptides against Candida albicans. Microorganisms 2020, 8, 758. https://doi.org/10.3390/microorganisms8050758
Ramamourthy G, Park J, Seo C, J. Vogel H, Park Y. Antifungal and Antibiofilm Activities and the Mechanism of Action of Repeating Lysine-Tryptophan Peptides against Candida albicans. Microorganisms. 2020; 8(5):758. https://doi.org/10.3390/microorganisms8050758
Chicago/Turabian StyleRamamourthy, Gopal, Jonggwan Park, Changho Seo, Hans J. Vogel, and Yoonkyung Park. 2020. "Antifungal and Antibiofilm Activities and the Mechanism of Action of Repeating Lysine-Tryptophan Peptides against Candida albicans" Microorganisms 8, no. 5: 758. https://doi.org/10.3390/microorganisms8050758
APA StyleRamamourthy, G., Park, J., Seo, C., J. Vogel, H., & Park, Y. (2020). Antifungal and Antibiofilm Activities and the Mechanism of Action of Repeating Lysine-Tryptophan Peptides against Candida albicans. Microorganisms, 8(5), 758. https://doi.org/10.3390/microorganisms8050758