Porphyromonas gingivalis, a Long-Range Pathogen: Systemic Impact and Therapeutic Implications



{kind=link}
Abstract
:1. Introduction
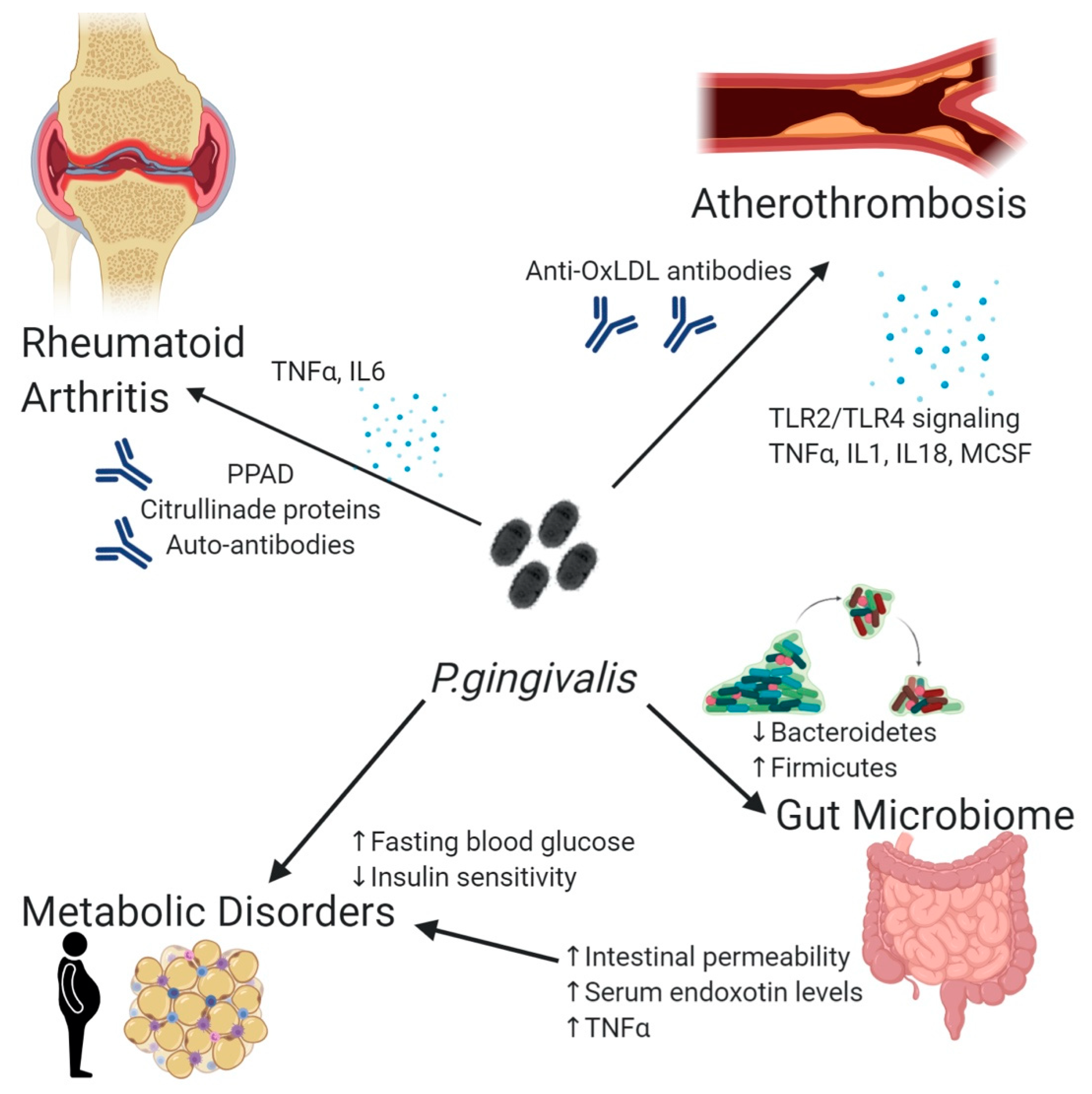
2. Distant Dissemination of P. gingivalis
3. Pathogenicity of P. gingivalis: The Case of Atherothrombosis
4. P. gingivalis and Rheumatoid Arthritis
5. Impact of P. gingivalis on Obesity, Metabolism, and Gut Microbiota Homeostasis
6. Clinical Implications
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Caton, J.G.; Armitage, G.; Berglundh, T.; Chapple, I.L.C.; Jepsen, S.; Kornman, K.S.; Mealey, B.L.; Papapanou, P.N.; Sanz, M.; Tonetti, M.S. A new classification scheme for periodontal and peri-implant diseases and conditions—Introduction and key changes from the 1999 classification. J. Periodontol. 2018, 89, S1–S8. [Google Scholar] [CrossRef] [PubMed]
- Kinane, D.F.; Stathopoulou, P.G.; Papapanou, P.N. Periodontal diseases. Nat. Rev. Dis. Primer 2017, 3, 17038. [Google Scholar] [CrossRef] [PubMed]
- Kassebaum, N.J.; Bernabe, E.; Dahiya, M.; Bhandari, B.; Murray, C.J.L.; Marcenes, W. Global Burden of Severe Periodontitis in 1990-2010: A Systematic Review and Meta-regression. J. Dent. Res. 2014, 93, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, L.; Saub, R.; Baharuddin, N.A.; Safii, S.H.; Gopal Krishna, V.; Bartold, P.M.; Vaithilingam, R.D. Impact of Severe Chronic Periodontitis on Oral Health-related Quality of Life. Oral Health Prev. Dent. 2019, 17, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Linden, G.J.; Lyons, A.; Scannapieco, F.A. Periodontal systemic associations: Review of the evidence. J. Clin. Periodontol. 2013, 40, 8–19. [Google Scholar] [CrossRef]
- Keijser, B.J.F.; Zaura, E.; Huse, S.M.; van der Vossen, J.M.B.M.; Schuren, F.H.J.; Montijn, R.C.; ten Cate, J.M.; Crielaard, W. Pyrosequencing analysis of the oral microflora of healthy adults. J. Dent. Res. 2008, 87, 1016–1020. [Google Scholar] [CrossRef]
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The oral microbiota: Dynamic communities and host interactions. Nat. Rev. Microbiol. 2018, 16, 745–759. [Google Scholar] [CrossRef]
- Zenobia, C.; Hajishengallis, G. Porphyromonas gingivalis virulence factors involved in subversion of leukocytes and microbial dysbiosis. Virulence 2015, 6, 236–243. [Google Scholar] [CrossRef] [Green Version]
- Darveau, R.P. Periodontitis: A polymicrobial disruption of host homeostasis. Nat. Rev. Microbiol. 2010, 8, 481–490. [Google Scholar] [CrossRef]
- Zenobia, C.; Hasturk, H.; Nguyen, D.; Dyke, T.E.V.; Kantarci, A.; Darveau, R.P. Porphyromonas gingivalis Lipid A Phosphatase Activity Is Critical for Colonization and Increasing the Commensal Load in the Rabbit Ligature Model. Infect. Immun. 2014, 82, 650–659. [Google Scholar] [CrossRef] [Green Version]
- Hajishengallis, G.; Liang, S.; Payne, M.A.; Hashim, A.; Jotwani, R.; Eskan, M.A.; McIntosh, M.L.; Alsam, A.; Kirkwood, K.L.; Lambris, J.D.; et al. Low-Abundance Biofilm Species Orchestrates Inflammatory Periodontal Disease through the Commensal Microbiota and Complement. Cell Host Microbe 2011, 10, 497–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Awady, A.; de Sousa Rabelo, M.; Meghil, M.M.; Rajendran, M.; Elashiry, M.; Stadler, A.F.; Foz, A.M.; Susin, C.; Romito, G.A.; Arce, R.M.; et al. Polymicrobial synergy within oral biofilm promotes invasion of dendritic cells and survival of consortia members. Npj Biofilms Microbiomes 2019, 5, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendrickson, E.L.; Wang, T.; Dickinson, B.C.; Whitmore, S.E.; Wright, C.J.; Lamont, R.J.; Hackett, M. Proteomics of Streptococcus gordonii within a model developing oral microbial community. BMC Microbiol. 2012, 12, 211. [Google Scholar] [CrossRef] [Green Version]
- Ahn, S.H.; Song, J.-E.; Kim, S.; Cho, S.-H.; Lim, Y.K.; Kook, J.-K.; Kook, M.-S.; Lee, T.-H. NOX1/2 activation in human gingival fibroblasts by Fusobacterium nucleatum facilitates attachment of Porphyromonas gingivalis. Arch. Microbiol. 2016, 198, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Olsen, I. Porphyromonas gingivalis and its CRISPR-Cas system. J. Oral Microbiol. 2019, 11, 1638196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cugini, C.; Klepac-Ceraj, V.; Rackaityte, E.; Riggs, J.E.; Davey, M.E. Porphyromonas gingivalis: Keeping the pathos out of the biont. J. Oral Microbiol. 2013, 5, 19804. [Google Scholar] [CrossRef]
- Bugueno, I.M.; Batool, F.; Korah, L.; Benkirane-Jessel, N.; Huck, O. Porphyromonas gingivalis Differentially Modulates Apoptosome Apoptotic Peptidase Activating Factor 1 in Epithelial Cells and Fibroblasts. AJPA 2018, 188, 404–416. [Google Scholar] [CrossRef] [Green Version]
- Bugueno, I.M.; Batool, F.; Keller, L.; Kuchler-Bopp, S.; Benkirane-Jessel, N.; Huck, O. Porphyromonas gingivalis bypasses epithelial barrier and modulates fibroblastic inflammatory response in an in vitro 3D spheroid model. Sci. Rep. 2018, 8, 30. [Google Scholar] [CrossRef] [Green Version]
- Huck, O.; Elkaim, R.; Davideau, J.-L.; Tenenbaum, H. Porphyromonas gingivalis-impaired innate immune response via NLRP3 proteolysis in endothelial cells. Innate Immun. 2014, 21, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Ding, L.; Liang, D.; Luo, L. Porphyromonas gingivalis inhibits M2 activation of macrophages by suppressing α-ketoglutarate production in mice. Mol. Oral Microbiol. 2018, 33, 388–395. [Google Scholar] [CrossRef]
- Sochalska, M.; Potempa, J. Manipulation of Neutrophils by Porphyromonas gingivalis in the Development of Periodontitis. Front. Cell. Infect. Microbiol. 2017, 7, 197. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, H.; Hirano, T.; Whitmore, S.E.; Morisaki, I.; Amano, A.; Lamont, R.J. The Serine Phosphatase SerB of Porphyromonas gingivalis Suppresses IL-8 Production by Dephosphorylation of NF-κB RelA/p65. PLoS Pathog. 2013, 9, e1003326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maekawa, T.; Krauss, J.L.; Abe, T.; Jotwani, R.; Triantafilou, M.; Triantafilou, K.; Hashim, A.; Hoch, S.; Curtis, M.A.; Nussbaum, G.; et al. Porphyromonas gingivalis manipulates complement and TLR signaling to uncouple bacterial clearance from inflammation and promote dysbiosis. Cell Host Microbe 2014, 15, 768–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popadiak, K.; Potempa, J.; Riesbeck, K.; Blom, A.M. Biphasic Effect of Gingipains from Porphyromonas gingivalis on the Human Complement System. J. Immunol. 2007, 178, 7242–7250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, E.-C.; Choi, S.-Y.; Lee, Y.; Park, J.-W.; Hong, S.-H.; Lee, H.-J. Extracellular RNAs in periodontopathogenic outer membrane vesicles promote TNF-α production in human macrophages and cross the blood-brain barrier in mice. FASEB J. 2019. [Google Scholar] [CrossRef] [Green Version]
- Lapérine, O.; Cloitre, A.; Caillon, J.; Huck, O.; Bugueno, I.M.; Pilet, P.; Sourice, S.; Le Tilly, E.; Palmer, G.; Davideau, J.-L.; et al. Interleukin-33 and RANK-L Interplay in the Alveolar Bone Loss Associated to Periodontitis. PLoS ONE 2016, 11, e0168080-17. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Gupte, R.; Zelkha, S.; Amar, S. Receptor activator of nuclear factor kappa B ligand antagonists inhibit tissue inflammation and bone loss in experimental periodontitis. J. Clin. Periodontol. 2011, 38, 1029–1036. [Google Scholar] [CrossRef]
- Huck, O.; Saadi-Thiers, K.; Tenenbaum, H.; Davideau, J.-L.; Romagna, C.; Laurent, Y.; Cottin, Y.; Roul, J.G. Evaluating periodontal risk for patients at risk of or suffering from atherosclerosis: Recent biological hypotheses and therapeutic consequences. Arch. Cardiovasc. Dis. 2017, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Preshaw, P.M.; Alba, A.L.; Herrera, D.; Jepsen, S.; Konstantinidis, A.; Makrilakis, K.; Taylor, R. Periodontitis and diabetes: A two-way relationship. Diabetologia 2012, 55, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Nibali, L.; Tatarakis, N.; Needleman, I.; Tu, Y.-K.; D’Aiuto, F.; Rizzo, M.; Donos, N. Association between Metabolic Syndrome and Periodontitis: A Systematic Review and Meta-analysis. J. Clin. Endocrinol. Metab. 2013, 98, 913–920. [Google Scholar] [CrossRef] [Green Version]
- Chou, Y.-Y.; Lai, K.-L.; Chen, D.-Y.; Lin, C.-H.; Chen, H.-H. Rheumatoid Arthritis Risk Associated with Periodontitis Exposure: A Nationwide, Population-Based Cohort Study. PLoS ONE 2015, 10, e0139693. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, G.G.; Peres, M.A.; Mittinty, M.N.; Peres, K.G.; Do, L.G.; Horta, B.L.; Gigante, D.P.; Corrêa, M.B.; Demarco, F.F. Diet-Induced Overweight and Obesity and Periodontitis Risk: An Application of the Parametric G-Formula in the 1982 Pelotas Birth Cohort. Am. J. Epidemiol. 2017, 185, 442–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linden, G.J.; Linden, K.; Yarnell, J.; Evans, A.; Kee, F.; Patterson, C.C. All-cause mortality and periodontitis in 60–70-year-old men: A prospective cohort study. J. Clin. Periodontol. 2012, 39, 940–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikawa, M.; Yoshida, K.; Okamura, H.; Ochiai, K.; Takamura, H.; Fujiwara, N.; Ozaki, K. Oral Porphyromonas gingivalis Translocates to the Liver and Regulates Hepatic Glycogen Synthesis through the Akt/GSK-3β Signaling Pathway. Biochim. Biophys. Acta 2013, 1832, 2035–2043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, S.H.; Nam, Y.; Jung, H.; Kim, J.; Rim, Y.A.; Park, N.; Lee, K.; Choi, S.; Jang, Y.; Kim, Y.; et al. Interrupting Oral Infection of Porphyromons gingivalis with anti-FimA Antibody Attenuates Bacterial Dissemination to the Arthritic Joint and Improves Experimental Arthritis. Exp. Mol. Med. 2018, 50, 460. [Google Scholar] [CrossRef] [PubMed]
- Panza, F.; Lozupone, M.; Solfrizzi, V.; Watling, M.; Imbimbo, B.P. Time to test antibacterial therapy in Alzheimer’s disease. Brain 2019, 142, 2905–2929. [Google Scholar] [CrossRef]
- Rosenfeld, M.E.; Campbell, L.A. Pathogens and atherosclerosis: Update on the potential contribution of multiple infectiou organisms to the pathogenesis of atherosclerosis. Thromb. Haemost. 2011, 106, 858–867. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Messas, E.; Batista, E.L.; Levine, R.A.; Amar, S. Porphyromonas gingivalis infection accelerates the progression of atherosclerosis in a heterozygous apolipoprotein E-deficient murine model. Circulation 2002, 105, 861–867. [Google Scholar]
- Sessa, R.; Di Pietro, M.; Schiavoni, G.; Petrucca, A.; Cipriani, P.; Zagaglia, C.; Nicoletti, M.; Santino, I.; del Piano, M. Measurement of Chlamydia pneumoniae bacterial load in peripheral blood mononuclear cells may be helpful to assess the state of chlamydial infection in patients with carotid atherosclerotic disease. Atherosclerosis 2007, 195, 224–230. [Google Scholar] [CrossRef]
- Izadi, M.; Fazel, M.; Sharubandi, S.H.; Saadat, S.H.; Farahani, M.M.; Nasseri, M.H.; Dabiri, H.; SafiAryan, R.; Esfahani, A.A.; Ahmadi, A.; et al. Helicobacter Species in the Atherosclerotic Plaques of Patients with Coronary Artery Disease. Cardiovasc Pathol. 2012, 21, 307–311. [Google Scholar] [CrossRef]
- Elkaim, R.; Dahan, M.; Kocgozlu, L.; Werner, S.; Kanter, D.; Kretz, J.G.; Tenenbaum, H. Prevalence of periodontal pathoges in subgingival lesions, atherosclerotic plaques and healthy blood vessels: A preliminary study. J. Periodontal Res. 2008, 43, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Rangé, H.; Labreuche, J.; Louedec, L.; Rondeau, P.; Planesse, C.; Sebbag, U.; Bourdon, E.; Michel, J.-B.; Bouchard, P.; Meilhac, O. Periodontal bacteria in human carotid atherothrombosis as a potential trigger for neutrophil activation. Atherosclerosis 2014, 236, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Michel, R.; Cohen, J.; Decarlo, A.; Kozarov, E. Intracellular survival and vascular cell-to-cell transmission of Porphyromonas gingivalis. BMC Microbiol. 2008, 8, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, J.S.; Kim, S.; Boström, K.I.; Wang, C.-Y.; Kim, R.H.; Park, N.-H. Periodontitis-induced systemic inflammation exacerbates atherosclerosis partly via endothelial–mesenchymal transition in mice. Int. J. Oral Sci. 2019, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lalla, E.; Lamster, I.B.; Hofmann, M.A.; Bucciarelli, L.; Jerud, A.P.; Tucker, S.; Lu, Y.; Papapanou, P.N.; Schmidt, A.M. Oral Infection With a Periodontal Pathogen Accelerates Early Atherosclerosis in Apolipoprotein E–Null Mice. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1405–1411. [Google Scholar] [CrossRef] [PubMed]
- Saadi-Thiers, K.; Huck, O.; Simonis, P.; Tilly, P.; Fabre, J.-E.; Tenenbaum, H.; Davideau, J.-L. Periodontal and systemic responses in various mice models of experimental periodontitis: Respective roles of inflammation duration and Porphyromonas gingivalis infection. J. Periodontol. 2013, 84, 396–406. [Google Scholar] [CrossRef]
- Chi, H.; Messas, E.; Levine, R.A.; Graves, D.T.; Amar, S. Interleukin-1 Receptor Signaling Mediates Atherosclerosis Associated with Bacterial Exposure and/or a High-Fat Diet in a Murine Apolipoprotein E Heterozygote Model. Circulation 2004, 110, 1678–1685. [Google Scholar] [CrossRef] [Green Version]
- Chukkapalli, S.S.; Ambadapadi, S.; Varkoly, K.; Jiron, J.; Aguirre, J.I.; Bhattacharyya, I.; Morel, L.M.; Lucas, A.R.; Kesavalu, L. Impaired innate immune signaling due to combined Toll-like receptor 2 and 4 deficiency affects both periodontitis and atherosclerosis in response to polybacterial infection. Pathog. Dis. 2018, 76. [Google Scholar] [CrossRef]
- Huck, O.; Elkaim, R.; Davideau, J.-L.; Tenenbaum, H. Porphyromonas gingivalis and its lipopolysaccharide differentially regulate the expression of cathepsin B in endothelial cells. Mol. Oral Microbiol. 2012. [Google Scholar] [CrossRef]
- Madan, M.; Amar, S. Toll-like receptor-2 mediates diet and/or pathogen associated atherosclerosis: Proteomic findings. PLoS ONE 2008, 3, e3204. [Google Scholar] [CrossRef] [Green Version]
- Amar, S.; Wu, S.; Madan, M. Is Porphyromonas gingivalis cell invasion required for atherogenesis? Pharmacotherapeutic implications. J. Immunol. 2009, 182, 1584–1592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, D.; Li, H.; Lei, L.; Chen, C.; Yan, F. Nonsurgical periodontal treatment reduced aortic inflammation in ApoE−/− mice with periodontitis. SpringerPlus 2016, 5, 940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhi, R.; Wang, C.; Nissinen, A.E.; Kankaanpää, J.; Bloigu, R.; Paju, S.; Mäntylä, P.; Buhlin, K.; Sinisalo, J.; Pussinen, P.J.; et al. Salivary IgA to MAA-LDL and Oral Pathogens Are Linked to Coronary Disease. J. Dent. Res. 2019, 98, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Kyrklund, M.; Kummu, O.; Kankaanpää, J.; Akhi, R.; Nissinen, A.; Turunen, S.P.; Pussinen, P.; Wang, C.; Hörkkö, S. Immunization with gingipain A hemagglutinin domain of Porphyromonas gingivalis induces IgM antibodies binding to malondialdehyde-acetaldehyde modified low-density lipoprotein. PLoS ONE 2018, 13, e0191216. [Google Scholar] [CrossRef] [Green Version]
- Bugueno, I.M.; Khelif, Y.; Seelam, N.; Morand, D.-N.; Tenenbaum, H.; Davideau, J.-L.; Huck, O. Porphyromonas gingivalis Differentially Modulates Cell Death Profile in Ox-LDL and TNF-α Pre-Treated Endothelial Cells. PLoS ONE 2016, 11, e0154590-18. [Google Scholar] [CrossRef]
- O’Connor, C.M.; Dunne, M.W.; Pfeffer, M.A.; Muhlestein, J.B.; Yao, L.; Gupta, S.; Benner, R.J.; Fisher, M.R.; Cook, T.D. Investigators in the WIZARD Study. Azithromycin for the Secondary Prevention of Coronary Heart Disease Events: The WIZARD Study: A Randomized Controlled Trial. JAMA 2003, 290, 1459–1466. [Google Scholar] [CrossRef] [Green Version]
- Grayston, J.T.; Kronmal, R.A.; Jackson, L.A.; Parisi, A.F.; Muhlestein, J.B.; Cohen, J.D.; Rogers, W.J.; Crouse, J.R.; Borrowdale, S.L.; Schron, E.; et al. ACES Investigators. Azithromycin for the Secondary Prevention of Coronary Events. N. Engl. J. Med. 2005, 352, 1637–1645. [Google Scholar] [CrossRef] [Green Version]
- Cannon, C.P.; Braunwald, E.; McCabe, C.H.; Grayston, J.T.; Muhlestein, B.; Giugliano, R.P.; Cairns, R.; Skene, A.M. Antibiotic treatment of Chlamydia pneumoniae after acute coronary syndrome. N. Engl. J. Med. 2005, 352, 1646–1654. [Google Scholar] [CrossRef] [Green Version]
- Jespersen, C.M.; Als-Nielsen, B.; Damgaard, M.; Hansen, J.F.; Hansen, S.; Helø, O.H.; Hildebrandt, P.; Hilden, J.; Jensen, G.B.; Kastrup, J.; et al. Randomised placebo controlled multicentre trial to assess short term clarithromycin for patients with stable coronary heart disease: CLARICOR trial. BMJ 2006, 332, 22–27. [Google Scholar] [CrossRef] [Green Version]
- Joensen, J.B.; Juul, S.; Henneberg, E.; Thomsen, G.; Ostergaard, L.; Lindholt, J.S. Can long-term antibiotic treatment prevent progression of peripheral arterial occlusive disease? A large, randomized, double-blinded, placebo-controlled trial. Atherosclerosis 2008, 196, 937–942. [Google Scholar] [CrossRef]
- Campbell, L.A.; Rosenfeld, M.E. Infection and Atherosclerosis Development. Arch. Med. Res. 2015, 46, 339–350. [Google Scholar] [CrossRef] [Green Version]
- Grassi, W.; Angelis, R.D.; Lamanna, G.; Cervini, C. The clinical features of rheumatoid arthritis. Eur. J. Radiol. 1998, 7, S18–S24. [Google Scholar] [CrossRef]
- Mantle, A. The Etiology of Rheumatism Considered from a Bacterial Point of View. Br. Med. J. 1887, 1, 1381–1384. [Google Scholar] [CrossRef]
- Li, S.; Yu, Y.; Yue, Y.; Zhang, Z.; Su, K. Microbial Infection and Rheumatoid Arthritis. J. Clin. Cell. Immunol. 2013, 4. [Google Scholar] [CrossRef]
- Bingham, C.O., 3rd; Moni, M. Periodontal disease and rheumatoid arthritis: The evidence accumulates for complex pathobiologic interactions. Curr. Opin. Rheumatol. 2013, 25, 345–353. [Google Scholar] [CrossRef] [Green Version]
- Rutger, G.P. Rheumatoid arthritis and periodontitis—Inflammatory and infectious connections. Review of the literature. J. Oral Microbiol. 2012, 4. [Google Scholar] [CrossRef]
- Chukkapalli, S.; Rivera-Kweh, M.; Gehlot, P.; Velsko, I.; Bhattacharyya, I.; Calise, S.J.; Satoh, M.; Chan, E.K.L.; Holoshitz, J.; Kesavalu, L. Periodontal bacterial colonization in synovial tissues exacerbates collagen-induced arthritis in B10.RIII mice. Arthritis Res. Ther. 2016, 18, 161. [Google Scholar] [CrossRef] [Green Version]
- Okumura, G.; Kondo, N.; Sato, K.; Yamazaki, K.; Ohshima, H.; Kawashima, H.; Ogose, A.; Endo, N. Experimental arthritis and Porphyromonas gingivalis administration synergistically decrease bone regeneration in femoral cortical defects. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Huck, O.; You, J.; Han, X.; Cai, B.; Panek, J.; Amar, S. Reduction of Articular and Systemic Inflammation by Kava-241 in Porphyromonas gingivalis-induced Arthritis Murine Model. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, M.; Yamashita, K.; Yoshie, H.; Hara, K. Detection of, and anti-collagen antibody produced by, CD5-positive B cells in inflamed gingival tissues. J. Periodontal Res. 1992, 27, 489–498. [Google Scholar] [CrossRef]
- Harre, U.; Georgess, D.; Bang, H.; Bozec, A.; Axmann, R.; Ossipova, E.; Jakobsson, P.-J.; Baum, W.; Nimmerjahn, F.; Szarka, E.; et al. Induction of osteoclastogenesis and bone loss by human autoantibodies against citrullinated vimentin. J. Clin. Investig. 2012, 122, 1791–1802. [Google Scholar] [CrossRef]
- Elkaim, R.; Bugueno-Valdebenito, I.M.; Benkirane-Jessel, N.; Tenenbaum, H. Porphyromonas gingivalis and its LPS differentially regulate the expression of peptidyl arginine deiminases in human chondrocytes. Innate Immun. 2017, 23, 468–475. [Google Scholar] [CrossRef]
- Maresz, K.J.; Hellvard, A.; Sroka, A.; Adamowicz, K.; Bielecka, E.; Koziel, J.; Gawron, K.; Mizgalska, D.; Marcinska, K.A.; Benedyk, M.; et al. Porphyromonas gingivalis facilitates the development and progression of destructive arthritis through its unique bacterial peptidylarginine deiminase (PAD). PLoS Pathog. 2013, 9, e1003627. [Google Scholar] [CrossRef]
- Larsen, D.N.; Mikkelsen, C.E.; Kierkegaard, M.; Bereta, G.P.; Nowakowska, Z.; Kaczmarek, J.Z.; Potempa, J.; Højrup, P. Citrullinome of Porphyromonas gingivalis Outer Membrane Vesicles: Confident Identification of Citrullinated Peptides. Mol. Cell. Proteom. 2020, 19, 167–180. [Google Scholar] [CrossRef]
- Casiano-Colón, A.; Marquis, R.E. Role of the arginine deiminase system in protecting oral bacteria and an enzymatic basis for acid tolerance. Appl. Environ. Microbiol. 1988, 54, 1318–1324. [Google Scholar] [CrossRef] [Green Version]
- Niederman, R.; Brunkhorst, B.; Smith, S.; Weinreb, R.N.; Ryder, M.I. Ammonia as a potential mediator of adult human periodontal infection: Inhibition of neutrophil function. Arch. Oral Biol. 1990, 35, S205–S209. [Google Scholar] [CrossRef]
- Dahlen, G.; Basic, A.; Bylund, J. Importance of Virulence Factors for the Persistence of Oral Bacteria in the Inflamed Gingival Crevice and in the Pathogenesis of Periodontal Disease. J. Clin. Med. 2019, 8, 1339. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.; Jung, S.M.; Rim, Y.A.; Park, N.; Nam, Y.; Lee, J.; Park, S.-H.; Ju, J.H. Arthritic role of Porphyromonas gingivalis in collagen-induced arthritis mice. PLoS ONE 2017, 12, e0188698. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.-Y.; Chen, S.-Y.; Siao, S.-H.; Chang, J.T.; Xue, T.-Y.; Lee, Y.-H.; Jan, M.-S.; Tsay, G.J.; Zouali, M. Targeting a cysteine protease from a pathobiont alleviates experimental arthritis. Arthritis Res. Ther. 2020, 22, 114. [Google Scholar] [CrossRef]
- Kharlamova, N.; Jiang, X.; Sherina, N.; Potempa, B.; Israelsson, L.; Quirke, A.M.; Eriksson, K.; Yucel-Lindberg, T.; Venables, P.J.; Potempa, J.; et al. Antibodies to Porphyromonas gingivalis Indicate Interaction between Oral Infection, Smoking, and Risk Genes in Rheumatoid Arthritis Etiology. Arthritis Rheumatol. 2016, 68, 604–613. [Google Scholar] [CrossRef] [Green Version]
- Carmona, L.; Cross, M.; Williams, B.; Lassere, M.; March, L. Rheumatoid arthritis. Best Pract. Res. Clin. Rheumatol. 2010, 24, 733–745. [Google Scholar] [CrossRef] [PubMed]
- Kononoff, A.; Elfving, P.; Pussinen, P.; Hörkkö, S.; Kautiainen, H.; Arstila, L.; Laasonen, L.; Savolainen, E.; Niinisalo, H.; Rutanen, J.; et al. Association of rheumatoid arthritis disease activity and antibodies to periodontal bacteria with serum lipoprotein profile in drug naive patients. Ann. Med. 2020, 52, 32–42. [Google Scholar] [CrossRef]
- de Molon, R.S.; Rossa, C., Jr.; Thurlings, R.M.; Cirelli, J.A.; Koenders, M.I. Linkage of Periodontitis and Rheumatoid Arthritis: Current Evidence and Potential Biological Interactions. Int. J. Mol. Sci. 2019, 20, 4541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Białowąs, K.; Radwan-Oczko, M.; Duś-Ilnicka, I.; Korman, L.; Świerkot, J. Periodontal disease and influence of periodontal treatment on disease activity in patients with rheumatoid arthritis and spondyloarthritis. Rheumatol. Int. 2020, 40, 455–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, A.; Rohde, J.F.; Raymond, K.; Heitmann, B.L. The Association between Periodontal Disease and Overweight and Obesity: A Systematic Review. J. Periodontol. 2015, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Le Sage, F.; Meilhac, O.; Gonthier, M.-P. Porphyromonas gingivalis lipopolysaccharide induces pro-inflammatory adipokine secretion and oxidative stress by regulating Toll-like receptor-mediated signaling pathways and redox enzymes in adipocytes. Mol. Cell. Endocrinol. 2017, 446, 102–110. [Google Scholar] [CrossRef]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [Green Version]
- Richard, G.; Trivedi, N.; Belta, C.; Amar, S. Partial restoration of macrophage alteration from diet-induced obesity in response to Porphyromonas gingivalis infection. PLoS ONE 2013, 8, e70320. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D. Human gut microbiome: Hopes, threats and promises. Gut 2018, 67, 1716–1725. [Google Scholar] [CrossRef]
- Tilg, H.; Moschen, A.R. Microbiota and diabetes: An evolving relationship. Gut 2014, 63, 1513–1521. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lin, S.; Vanhoutte, P.M.; Woo, C.W.; Xu, A. Akkermansia Muciniphila Protects against Atherosclerosis by Preventing Metabolic Endotoxemia-Induced Inflammation in Apoe-/- Mice. Circulation 2016, 133, 2434–2446. [Google Scholar] [CrossRef] [Green Version]
- Baquero, F.; Nombela, C. The microbiome as a human organ. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2012, 18, 2–4. [Google Scholar] [CrossRef] [Green Version]
- Kundu, P.; Blacher, E.; Elinav, E.; Pettersson, S. Our Gut Microbiome: The Evolving Inner. Self. Cell 2017, 171, 1481–1493. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, A.M.L.; Sørensen, C.E.; Proctor, G.B.; Carpenter, G.H.; Ekström, J. Salivary secretion in health and disease. J. Oral Rehabil. 2018, 45, 730–746. [Google Scholar] [CrossRef] [PubMed]
- Saygun, I.; Nizam, N.; Keskiner, I.; Bal, V.; Kubar, A.; Açıkel, C.; Serdar, M.; Slots, J. Salivary infectious agents and periodontal disease status. J. Periodontal Res. 2011, 46, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Arimatsu, K.; Yamada, H.; Miyazawa, H.; Minagawa, T.; Nakajima, M.; Ryder, M.I.; Gotoh, K.; Motooka, D.; Nakamura, S.; Iida, T.; et al. Oral pathobiont induces systemic inflammation and metabolic changes associated with alteration of gut microbiota. Sci. Rep. 2014, 4, 4828. [Google Scholar] [CrossRef] [Green Version]
- Lourenςo, T.G.B.; Spencer, S.J.; Alm, E.J.; Colombo, A.P. Defining the gut microbiota in individuals with periodontal diseases: An exploratory study. J. Oral Microbiol. 2018, 10, 1487741. [Google Scholar] [CrossRef] [Green Version]
- Kato, T.; Yamazaki, K.; Nakajima, M.; Date, Y.; Kikuchi, J.; Hase, K.; Ohno, H.; Yamazaki, K.; D’Orazio, S.E.F. Oral Administration of Porphyromonas gingivalis Alters the Gut Microbiome and Serum Metabolome. mSphere 2018, 3, e00460-18. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, M.; Arimatsu, K.; Kato, T.; Matsuda, Y.; Minagawa, T.; Takahashi, N.; Ohno, H.; Yamazaki, K. Oral Administration of P. gingivalis Induces Dysbiosis of Gut Microbiota and Impaired Barrier Function Leading to Dissemination of Enterobacteria to the Liver. PLoS ONE 2015, 10, e0134234. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, Y.; Kato, T.; Takahashi, N.; Nakajima, M.; Arimatsu, K.; Minagawa, T.; Sato, K.; Ohno, H.; Yamazaki, K. Ligature-induced periodontitis in mice induces elevated levels of circulating interleukin-6 but shows only weak effects on adipose and liver tissues. J. Periodontal. Res. 2016, 51, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.L.; Vieira-Silva, S.; Liston, A.; Raes, J. How informative is the mouse for human gut microbiota research? Dis. Model. Mech. 2015, 8, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segata, N.; Haake, S.K.; Mannon, P.; Lemon, K.P.; Waldron, L.; Gevers, D.; Huttenhower, C.; Izard, J. Composition of the adult digestive tract bacterial microbiome based on seven mouth surfaces, tonsils, throat and stool samples. Genome Biol. 2012, 13, R42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z. The metabolic syndrome. Lancet 2005, 365, 1415–1428. [Google Scholar] [CrossRef]
- Cani, P.D.; Osto, M.; Geurts, L.; Everard, A. Involvement of gut microbiota in the development of low-grade inflammation and type 2 diabetes associated with obesity. Gut Microbes 2012, 3, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef] [Green Version]
- Amar, S.; Zhou, Q.; Shaik-Dasthagirisaheb, Y.; Leeman, S. Diet-induced obesity in mice causes changes in immune responses and bone loss manifested by bacterial challenge. Proc. Natl. Acad. Sci. USA 2007, 104, 20466–20471. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Leeman, S.E.; Amar, S. Signaling mechanisms in the restoration of impaired immune function due to diet-induced obesity. Proc. Natl. Acad. Sci. USA 2011, 108, 2867–2872. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Leeman, S.E.; Amar, S. Signaling mechanisms involved in altered function of macrophages from diet-induced obese mice affect immune responses. Proc. Natl. Acad. Sci. USA 2009, 106, 10740–10745. [Google Scholar] [CrossRef] [Green Version]
- Blasco-Baque, V.; Garidou, L.; Pomié, C.; Escoula, Q.; Loubieres, P.; Le Gall-David, S.; Lemaitre, M.; Nicolas, S.; Klopp, P.; Waget, A.; et al. Periodontitis induced by Porphyromonas gingivalis drives periodontal microbiota dysbiosis and insulin resistance via an impaired adaptive immune response. Gut 2017, 66, 872–885. [Google Scholar] [CrossRef] [Green Version]
- Grossi, S.G.; Skrepcinski, F.B.; DeCaro, T.; Robertson, D.C.; Ho, A.W.; Dunford, R.G.; Genco, R.J. Treatment of Periodontal Disease in Diabetics Reduces Glycated Hemoglobin. J. Periodontol. 1997, 68, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Kıran, M.; Arpak, N.; Ünsal, E.; Erdoğan, M.F. The effect of improved periodontal health on metabolic control in type 2 diabetes mellitus. J. Clin. Periodontol. 2005, 32, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Altay, U.; Gürgan, C.A.; Ağbaht, K. Changes in Inflammatory and Metabolic Parameters after Periodontal Treatment in Patients with and without Obesity. J. Periodontol. 2013, 84, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Lang, N.P.; Salvi, G.E.; Sculean, A. Nonsurgical therapy for teeth and implants-When and why? Periodontology 2019, 79, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.L.; Duarte, P.M.; Figueira, E.A.; Lemos, J.C.; Nobre, C.M.G.; Miranda, T.S.; de Vasconcelos Gurgel, B.C. Effects of a full-mouth disinfection protocol on the treatment of type-2 diabetic and non-diabetic subjects with mild-to-moderate periodontitis: One-year clinical outcomes. Clin. Oral Investig. 2019. [Google Scholar] [CrossRef]
- Harmouche, L.; Courval, A.; Mathieu, A.; Petit, C.; Huck, O.; Séverac, F.; Davideau, J.-L. Impact of tooth-related factors on photodynamic therapy effectiveness during active periodontal therapy: A 6-months split-mouth randomized clinical trial. Photodiagnosis Photodyn. Ther. 2019, 27, 167–172. [Google Scholar] [CrossRef]
- Mombelli, A.; Almaghlouth, A.; Cionca, N.; Courvoisier, D.S.; Giannopoulou, C. Differential benefits of amoxicillin-metronidazole in different phases of periodontal therapy in a randomized controlled crossover clinical trial. J. Periodontol. 2015, 86, 367–375. [Google Scholar] [CrossRef] [Green Version]
- Leininger, M.; Tenenbaum, H.; Davideau, J.-L. Modified periodontal risk assessment score: Long-term predictive value of treatment outcomes. Retrosp. Study 2010, 37, 427–435. [Google Scholar] [CrossRef]
- Petit, C.; Schmeltz, S.; Burgy, A.; Tenenbaum, H.; Huck, O.; Davideau, J.-L. Risk factors associated with long-term outcomes after active and supporting periodontal treatments: Impact of various compliance definitions on tooth loss. Clin. Oral Investig. 2019, 62, 218–219. [Google Scholar] [CrossRef]
- Huck, O.; Mulhall, H.; Rubin, G.; Kizelnik, Z.; Iyer, R.; Perpich, J.D.; Haque, N.; Cani, P.D.; de Vos, W.M.; Amar, S. Akkermansia muciniphila reduces Porphyromonas gingivalis-induced inflammation and periodontal bone destruction. J. Clin. Periodontol. 2019. [Google Scholar] [CrossRef]
- Martin-Cabezas, R.; Davideau, J.-L.; Tenenbaum, H.; Huck, O. Clinical efficacy of probiotic as an adjunctive therapy to non-surgical periodontal treatment of chronic periodontitis: A systematic review and meta-analysis. J. Clin. Periodontol. 2016, 43. [Google Scholar] [CrossRef] [PubMed]
- Geraldo, B.M.C.; Batalha, M.N.; Milhan, N.V.M.; Rossoni, R.D.; Scorzoni, L.; Anbinder, A.L. Heat-killed Lactobacillus reuteri and cell-free culture supernatant have similar effects to viable probiotics during interaction with Porphyromonas gingivalis. J. Periodontal Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Liu, Y.; Song, Y.; Gao, Y.; Zhao, F.; Luo, Y.; Qian, F.; Mu, G.; Tuo, Y. The ameliorative effect of Lactobacillus plantarum-12 on DSS-induced murine colitis. Food Funct. 2020. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L.; et al. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat. Med. 2017, 23, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Everard, A.; Plovier, H.; Rastelli, M.; Van Hul, M.; de Wouters d’Oplinter, A.; Geurts, L.; Druart, C.; Robine, S.; Delzenne, N.M.; Muccioli, G.G.; et al. Intestinal epithelial N-acylphosphatidylethanolamine phospholipase D links dietary fat to metabolic adaptations in obesity and steatosis. Nat. Commun. 2019, 10, 457. [Google Scholar] [CrossRef]
- Zhai, R.; Xue, X.; Zhang, L.; Yang, X.; Zhao, L.; Zhang, C. Strain-Specific Anti-inflammatory Properties of Two Akkermansia muciniphila Strains on Chronic Colitis in Mice. Front. Cell. Infect. Microbiol. 2019, 9, 239. [Google Scholar] [CrossRef]
- Cani, P.D.; de Vos, W.M. Next-Generation Beneficial Microbes: The Case of Akkermansia muciniphila. Front. Microbiol. 2017, 8, 1765. [Google Scholar] [CrossRef]
- Ottman, N.; Reunanen, J.; Meijerink, M.; Pietilä, T.E.; Kainulainen, V.; Klievink, J.; Huuskonen, L.; Aalvink, S.; Skurnik, M.; Boeren, S.; et al. Pili-like proteins of Akkermansia muciniphila modulate host immune responses and gut barrier function. PLoS ONE 2017, 12, e0173004. [Google Scholar] [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mulhall, H.; Huck, O.; Amar, S. Porphyromonas gingivalis, a Long-Range Pathogen: Systemic Impact and Therapeutic Implications. Microorganisms 2020, 8, 869. https://doi.org/10.3390/microorganisms8060869
Mulhall H, Huck O, Amar S. Porphyromonas gingivalis, a Long-Range Pathogen: Systemic Impact and Therapeutic Implications. Microorganisms. 2020; 8(6):869. https://doi.org/10.3390/microorganisms8060869
Chicago/Turabian StyleMulhall, Hannah, Olivier Huck, and Salomon Amar. 2020. "Porphyromonas gingivalis, a Long-Range Pathogen: Systemic Impact and Therapeutic Implications" Microorganisms 8, no. 6: 869. https://doi.org/10.3390/microorganisms8060869
APA StyleMulhall, H., Huck, O., & Amar, S. (2020). Porphyromonas gingivalis, a Long-Range Pathogen: Systemic Impact and Therapeutic Implications. Microorganisms, 8(6), 869. https://doi.org/10.3390/microorganisms8060869