Limited Response of Indigenous Microbes to Water and Nutrient Pulses in High-Elevation Atacama Soils: Implications for the Cold–Dry Limits of Life on Earth


Abstract
:1. Introduction
2. Materials and Methods
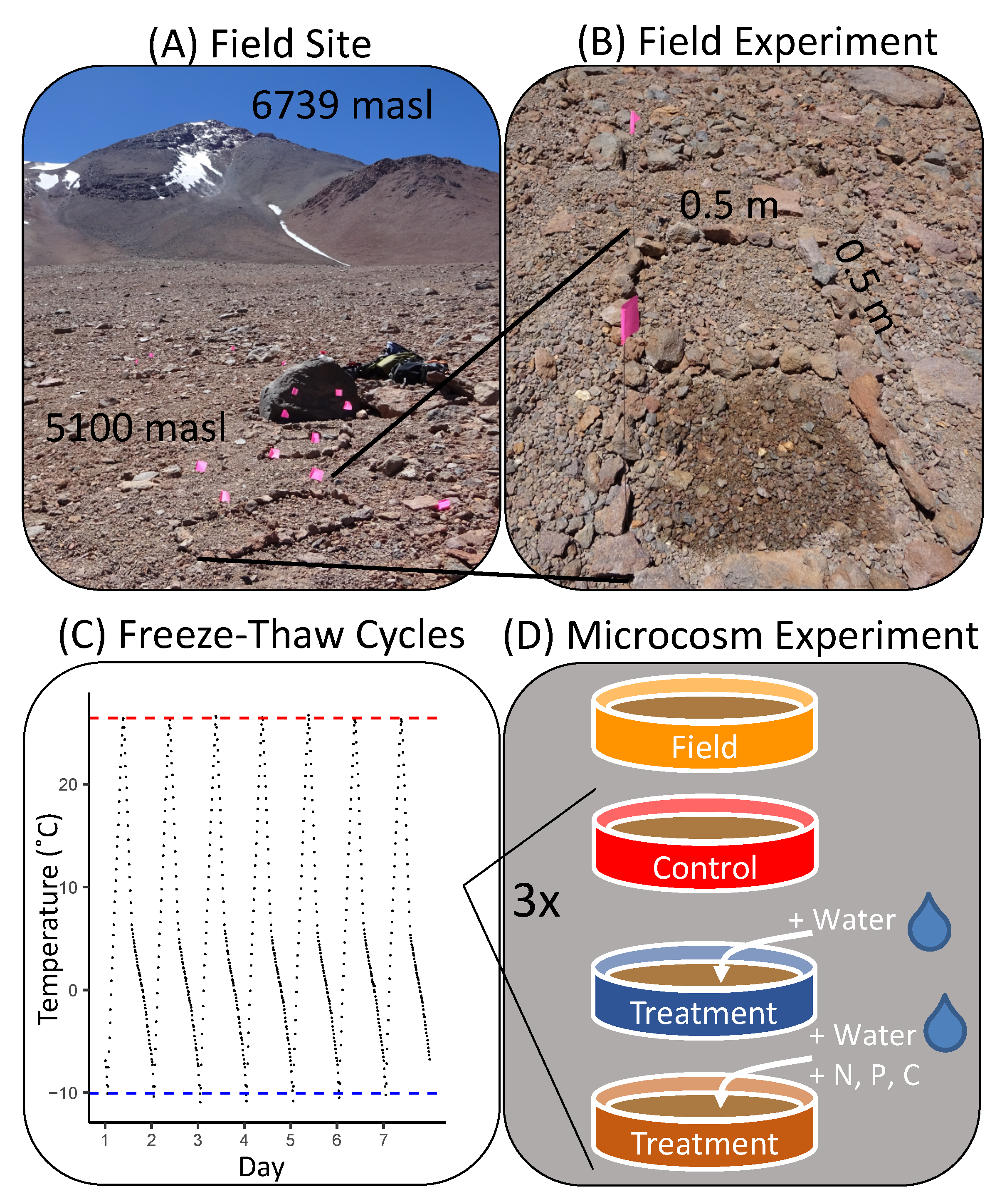
2.1. Soil Microcosm Manipulation
2.2. Field Experiment
2.3. DNA Extraction and Sequencing
2.4. Plate Counts
2.5. Data Processing and Statistical Analysis
3. Results
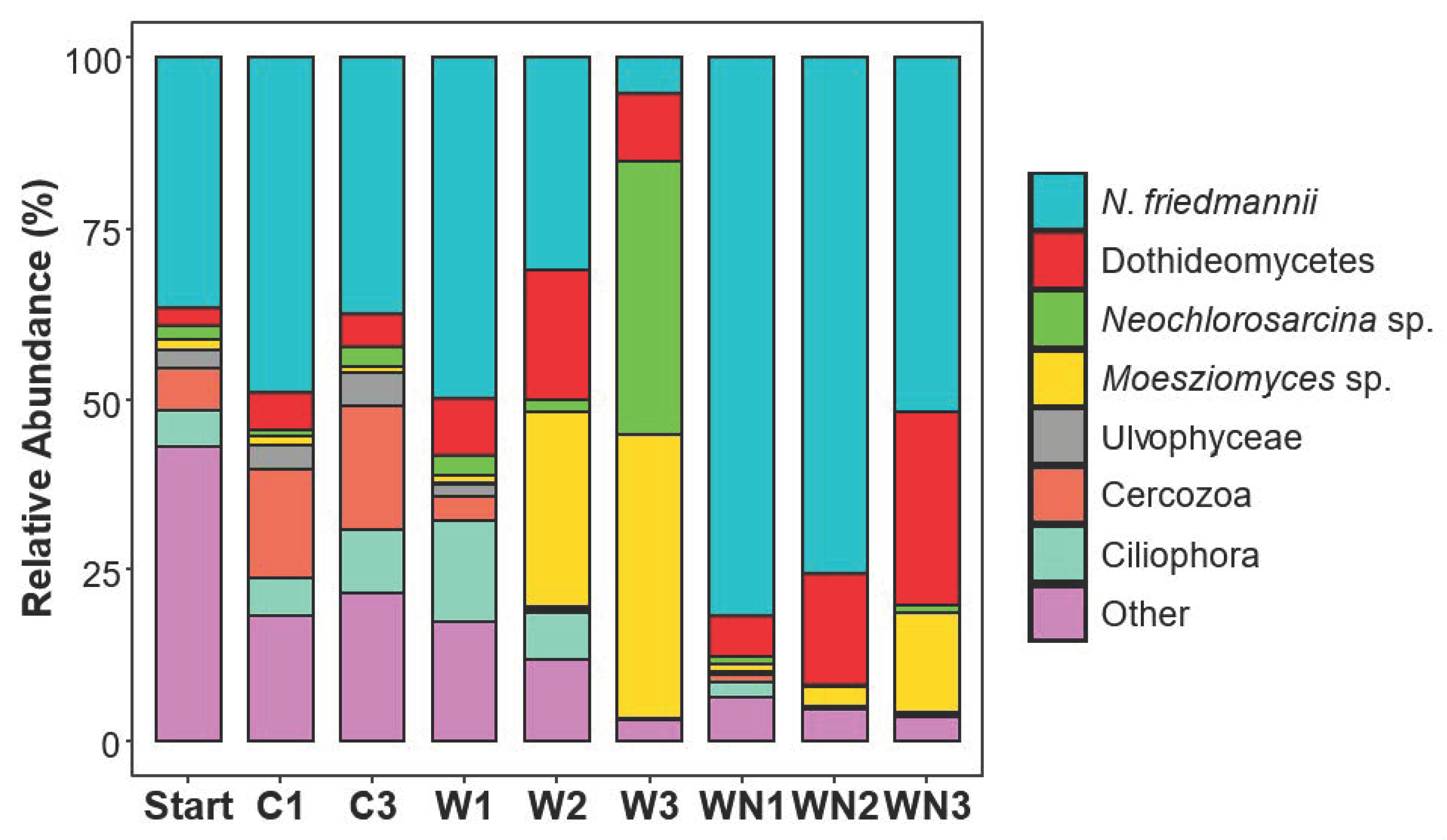
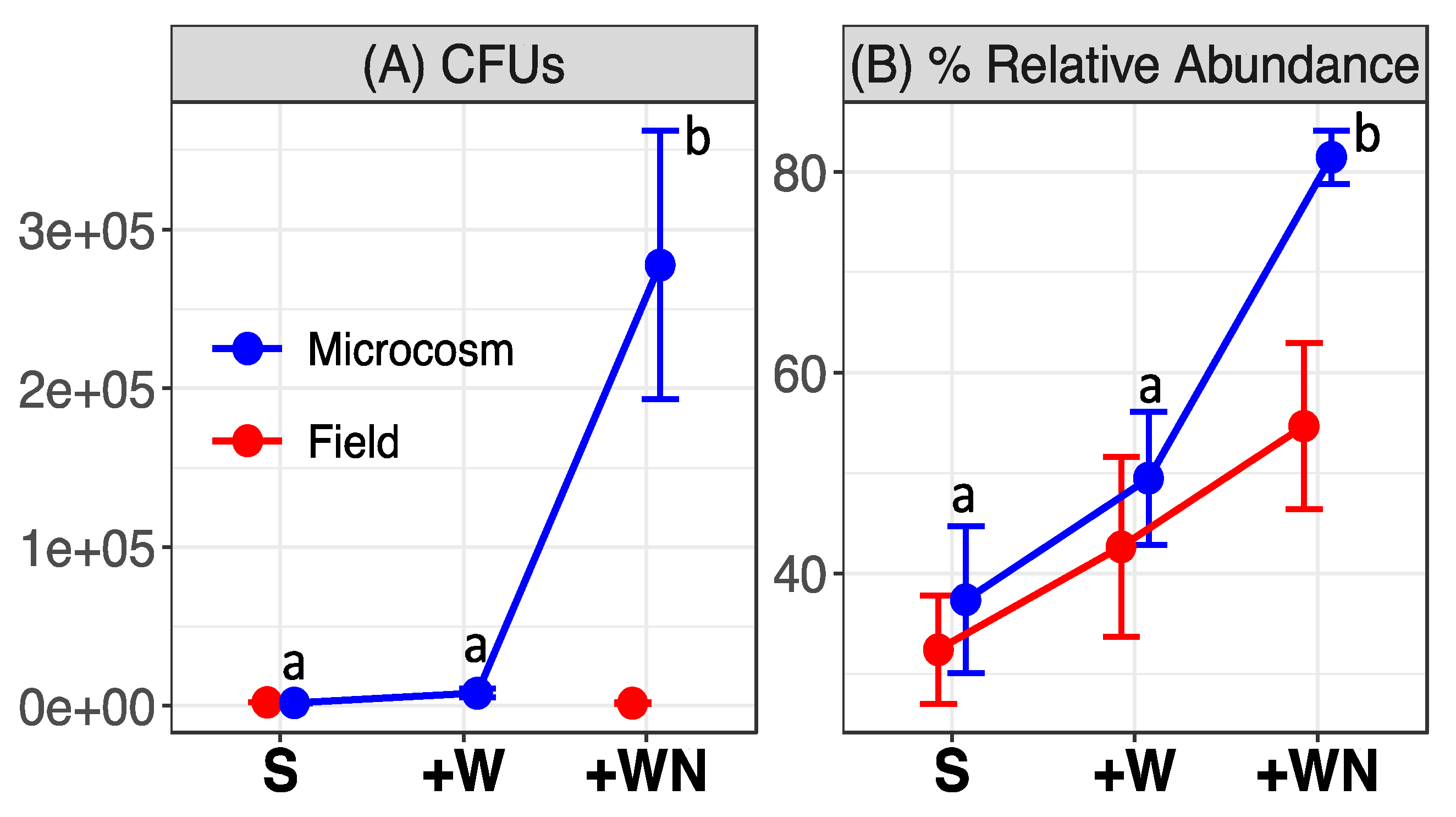
3.1. Naganishia friedmannii Response
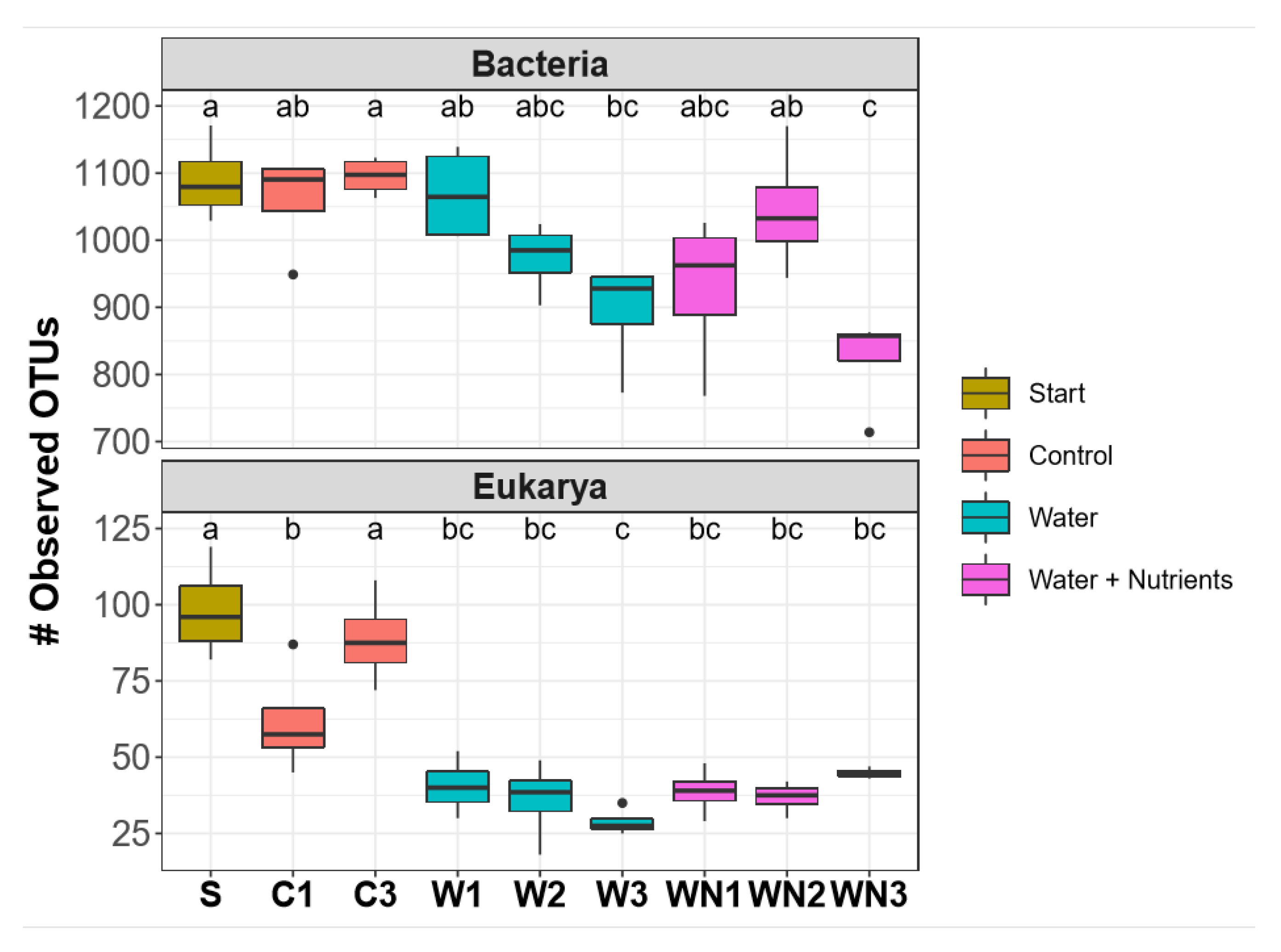
3.2. Alpha and Beta Diversity Patterns of Soil Microcosms
3.3. Taxonomic Shifts of Soil Microcosms
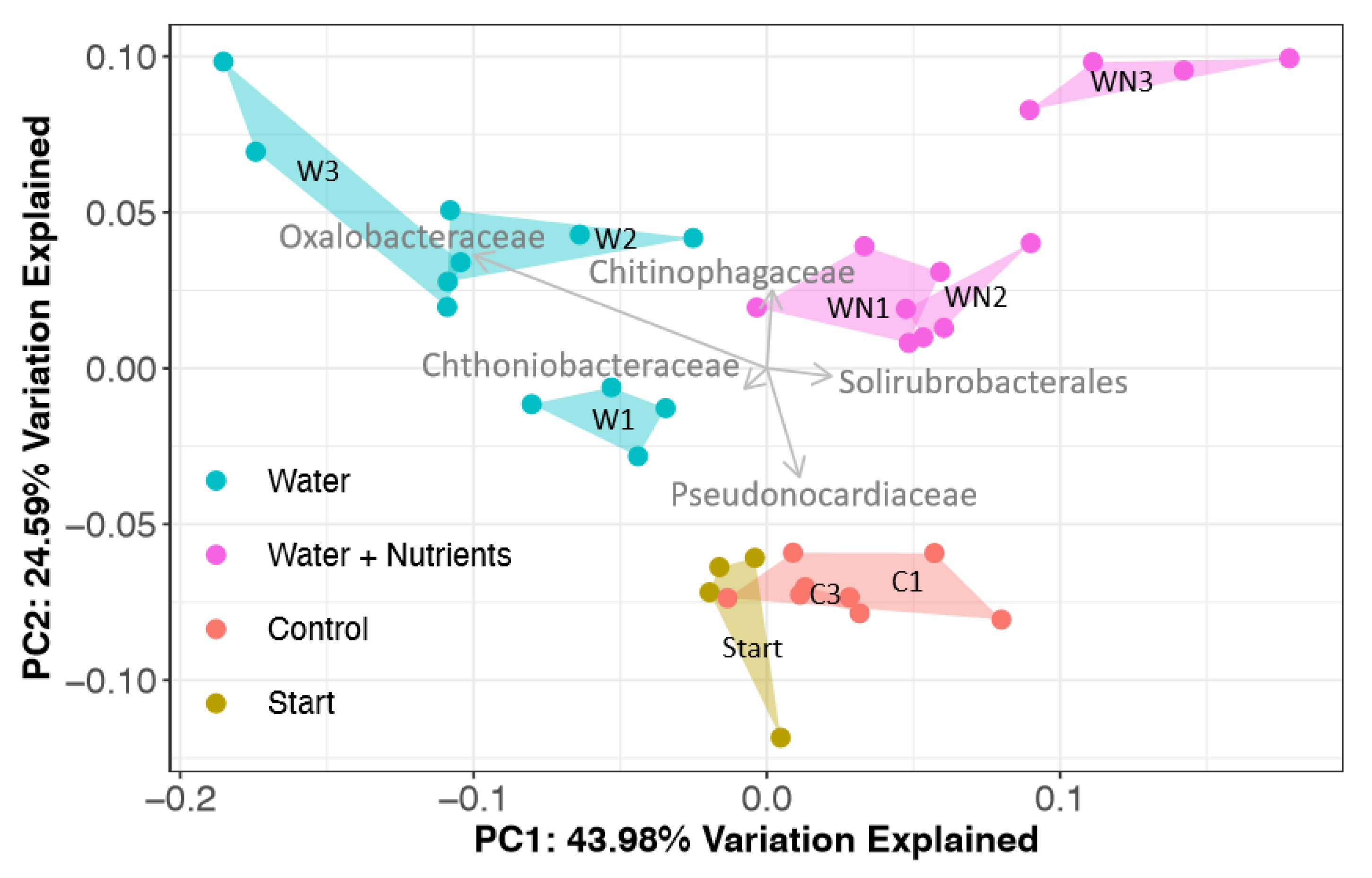
3.3.1. Bacterial Community Response
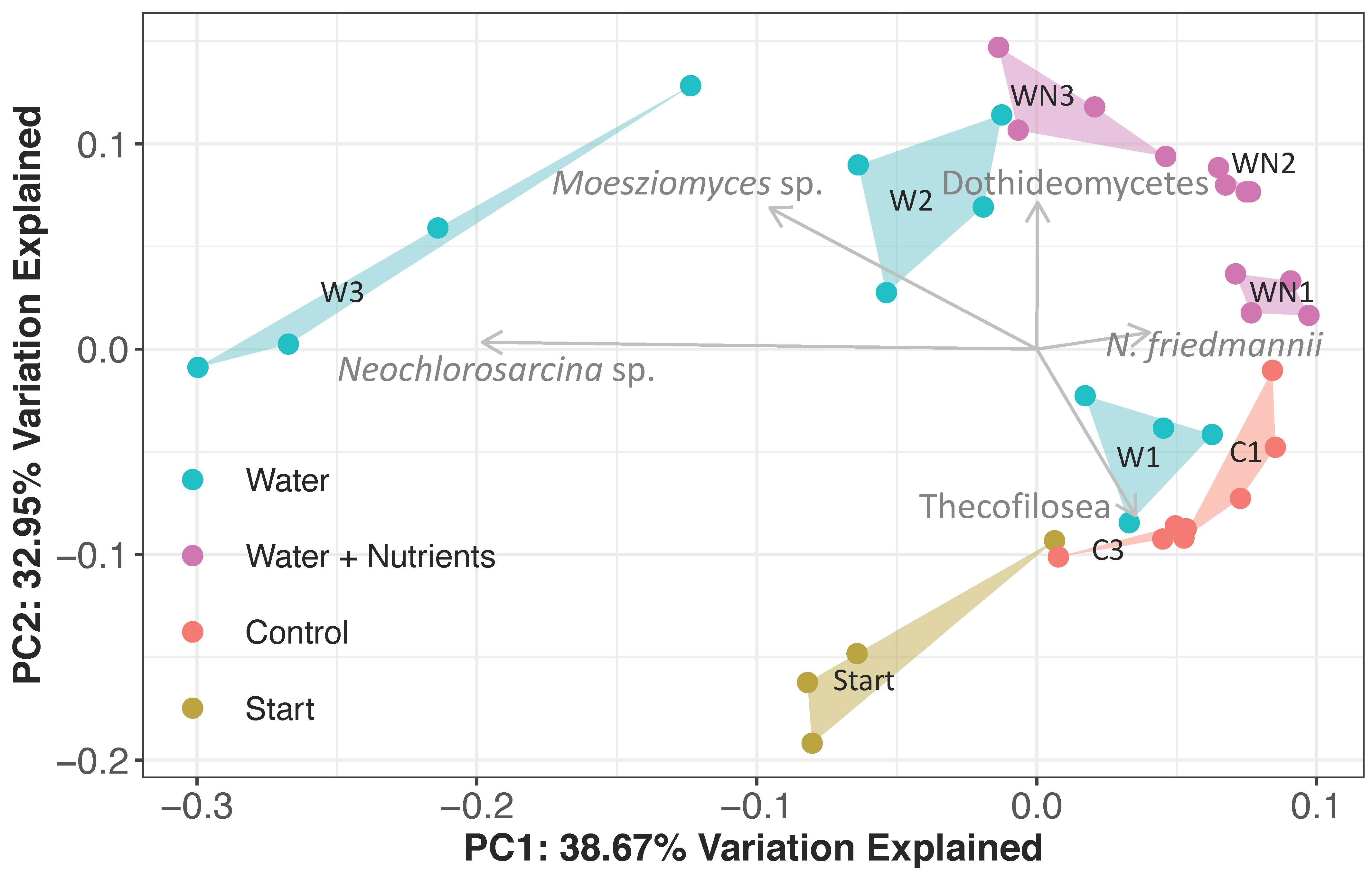
3.3.2. Eukaryotic Community Response
3.4. Environmental Community Response to Water and Nutrient Additions
4. Discussion
4.1. Naganishia friedmannii Response in Microcosms and in the Field
4.2. Bacterial Community Response in Soil Microcosms
4.2.1. Oxalobacteraceae
4.2.2. Bacterial Phototrophs
4.3. Eukaryotic Community Response in Soil Microcosms
4.3.1. Dothideomycetes
4.3.2. Moesziomyces sp.
4.3.3. Neochlorosarcina sp.
4.4. Astrobiological Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Costello, E.K.; Halloy, S.R.P.; Reed, S.C.; Sowell, P.; Schmidt, S.K. Fumarole-supported islands of biodiversity within a hyperarid, high-elevation landscape on socompa volcano, puna de atacama, andes. Appl. Environ. Microbiol. 2009, 75, 735–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, R.C.; King, A.J.; Farías, M.E.; Sowell, P.; Vitry, C.; Schmidt, S.K. The potential for microbial life in the highest-elevation (>6000 masl.) mineral soils of the Atacama region. J. Geophys. Res. Biogeosci. 2012, 117. [Google Scholar] [CrossRef]
- Vimercati, L.; Hamsher, S.; Schubert, Z.; Schmidt, S.K. Growth of high-elevation Cryptococcus sp. during extreme freeze–thaw cycles. Extremophiles 2016, 20, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.K.; Gendron, E.M.S.; Vincent, K.; Solon, A.J.; Sommers, P.; Schubert, Z.R.; Vimercati, L.; Porazinska, D.L.; Darcy, J.L.; Sowell, P. Life at extreme elevations on Atacama volcanoes: The closest thing to Mars on Earth? Antonie van Leeuwenhoek. 2018, 111, 1389–1401. [Google Scholar] [CrossRef]
- Cabrol, N.A.; Feister, U.; Häder, D.P.; Piazena, H.; Grin, E.A.; Klein, A. Record solar UV irradiance in the tropical Andes. Front. Environ. Sci. 2014, 2, 19. [Google Scholar] [CrossRef] [Green Version]
- Reinhard, J.; Stenzel, M. At 22,000 Feet Children of Inca Sacrifice Found Frozen in Time. Natl. Geogr. Mag. 1999, 196, 36–55. [Google Scholar]
- Reinhard, J.; Ceruti, M.C. Inca rituals and sacred mountains: A study of the world’s highest archaeological sites. Cotsen Inst. Archaeol. 2010, 67, 280. [Google Scholar]
- Wilson, A.S.; Brown, E.L.; Villa, C.; Lynnerup, N.; Healey, A.; Ceruti, M.C.; Reinhard, J.; Previgliano, C.H.; Araoz, F.A.; Diez, J.G.; et al. Archaeological, radiological, and biological evidence offer insight into Inca child sacrifice. Proc. Natl. Acad. Sci. USA 2013, 110, 13322–13327. [Google Scholar] [CrossRef]
- Solon, A.J.; Vimercati, L.; Darcy, J.L.; Arán, P.; Porazinska, D.; Dorador, C.; Farías, M.E.; Schmidt, S.K. Microbial Communities of High-Elevation Fumaroles, Penitentes, and Dry Tephra “Soils” of the Puna de Atacama Volcanic Zone. Microb. Ecol. 2018, 76, 340–351. [Google Scholar] [CrossRef]
- Vimercati, L.; Solon, A.J.; Krinsky, A.; Arán, P.; Porazinska, D.L.; Darcy, J.L.; Dorador, C.; Schmidt, S.K. Nieves penitentes are a new habitat for snow algae in one of the most extreme high-elevation environments on Earth. Arctic Antarct. Alp. Res. 2019, 51, 190–200. [Google Scholar] [CrossRef]
- Lynch, R.C.; Darcy, J.L.; Kane, N.C.; Nemergut, D.R.; Schmidt, S.K. Metagenomic evidence for metabolism of trace atmospheric gases by high-elevation desert actinobacteria. Front. Microbiol. 2014, 5, 698. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.M. The biological terraforming of Mars: Planetary ecosynthesis as ecological succession on a global scale. Astrobiology 2004, 4, 168–195. [Google Scholar] [CrossRef] [PubMed]
- Boynton, W.V.; Ming, D.W.; Kounaves, S.P.; Young, S.M.M.; Arvidson, R.E.; Hecht, M.H.; Hoffman, J.; Niles, P.B.; Hamara, D.K.; Quinn, R.C.; et al. Evidence for calcium carbonate at the mars phoenix landing site. Science 2009, 325, 61–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carini, P.; Marsden, P.J.; Leff, J.W.; Morgan, E.E.; Strickland, M.S.; Fierer, N. Relic DNA is abundant in soil and obscures estimates of soil microbial diversity. Nat. Microbiol. 2016, 2, 16242. [Google Scholar] [CrossRef] [PubMed]
- Borin, S.; Ventura, S.; Tambone, F.; Mapelli, F.; Schubotz, F.; Brusetti, L.; Scaglia, B.; D’Acqui, L.P.; Solheim, B.; Turicchia, S.; et al. Rock weathering creates oases of life in a High Arctic desert. Environ. Microbiol. 2010, 12, 293–303. [Google Scholar] [CrossRef]
- Cary, S.C.; McDonald, I.R.; Barrett, J.E.; Cowan, D.A. On the rocks: The microbiology of Antarctic Dry Valley soils. Nat. Rev. Microbiol. 2010, 8, 129–138. [Google Scholar] [CrossRef]
- Schmidt, S.K.; Lynch, R.C.; King, A.J.; Karki, D.; Robeson, M.S.; Nagy, L.; Williams, M.W.; Mitter, M.S.; Freeman, K.R. Phylogeography of microbial phototrophs in the dry valleys of the high Himalayas and Antarctica. Proc. R. Soc. B Biol. Sci. 2011, 278, 702–708. [Google Scholar] [CrossRef]
- Knelman, J.E.; Schmidt, S.K.; Lynch, R.C.; Darcy, J.L.; Castle, S.C.; Cleveland, C.C.; Nemergut, D.R. Nutrient addition dramatically accelerates microbial community succession. PLoS ONE 2014, 9, e102609. [Google Scholar] [CrossRef]
- Darcy, J.L.; Schmidt, S.K.; Knelman, J.E.; Cleveland, C.C.; Castle, S.C.; Nemergut, D.R. Phosphorus, not nitrogen, limits plants and microbial primary producers following glacial retreat. Sci. Adv. 2018, 4, eaaq0942. [Google Scholar] [CrossRef] [Green Version]
- Van Horn, D.J.; Van Horn, M.L.; Barrett, J.E.; Gooseff, M.N.; Altrichter, A.E.; Geyer, K.M.; Zeglin, L.H.; Takacs-Vesbach, C.D. Factors Controlling Soil Microbial Biomass and Bacterial Diversity and Community Composition in a Cold Desert Ecosystem: Role of Geographic Scale. PLoS ONE 2013, 8, e66103. [Google Scholar] [CrossRef]
- Van Horn, D.J.; Okie, J.G.; Buelow, H.N.; Gooseff, M.N.; Barrett, J.E.; Takacs-Vesbach, C.D. Soil microbial responses to increased moisture and organic resources along a salinity gradient in a polar desert. Appl. Environ. Microbiol. 2014, 80, 3034–3043. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, S.K.; Nemergut, D.R.; Miller, A.E.; Freeman, K.R.; King, A.J.; Seimon, A. Microbial activity and diversity during extreme freeze-thaw cycles in periglacial soils, 5400 m elevation, Cordillera Vilcanota, Perú. Extremophiles 2009, 13, 807–816. [Google Scholar] [CrossRef]
- Yergeau, E.; Kowalchuk, G.A. Responses of Antarctic soil microbial communities and associated functions to temperature and freeze-thaw cycle frequency. Environ. Microbiol. 2008, 10, 2223–2235. [Google Scholar] [CrossRef]
- Schmidt, D.; Schmidt, D. Das Extremklima der Nordchilenischen Hochatacama Unter Besonderer Berücksichtigung der Höhengradienten; Techn. Univ.: München, Germany, 1999. [Google Scholar]
- Schubert, Z. Dew Formation and Water Availability at High Elevation in the Atacama Desert, Chile. Undergraduate Honors Thesis, University of Colorado at Boulder, Boulder, CO, USA, 2014. [Google Scholar]
- Schmidt, S.K.; Vimercati, L.; Darcy, J.L.; Arán, P.; Gendron, E.M.S.; Solon, A.J.; Porazinska, D.; Dorador, C. A Naganishia in high places: Functioning populations or dormant cells from the atmosphere? Mycology 2017, 8, 153–163. [Google Scholar] [CrossRef]
- Schmidt, S.K.; Nemergut, D.R.; Todd, B.T.; Lynch, R.C.; Darcy, J.L.; Cleveland, C.C.; King, A.J. A simple method for determining limiting nutrients for photosynthetic crusts. Plant Ecol. Divers. 2012, 5, 513–519. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pẽa, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Darcy, J.L.; Schmidt, S.K. Nutrient limitation of microbial phototrophs on a debris-covered glacier. Soil Biol. Biochem. 2016, 95, 156–163. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a Chimera-Checked 16S rRNA Gene Database and Workbench Compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Bittinger, K.; Bushman, F.D.; Desantis, T.Z.; Andersen, G.L.; Knight, R. PyNAST: A flexible tool for aligning sequences to a template alignment. Bioinformatics 2010, 26, 266–267. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing large minimum evolution trees with profiles instead of a distance matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-6, 2019.
- Hervé, M. RVAideMemoire: Testing and Plotting Procedures for Biostatistics. R Package Version 0.9-73. 2019. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2019. [Google Scholar]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Nakada, T.; Tsuchida, Y.; Tomita, M. Improved taxon sampling and multigene phylogeny of unicellular chlamydomonads closely related to the colonial volvocalean lineage Tetrabaenaceae-Goniaceae-Volvocaceae (Volvocales, Chlorophyceae). Mol. Phylogenet. Evol. 2019, 130, 1–8. [Google Scholar] [CrossRef]
- Buzzini, P.; Branda, E.; Goretti, M.; Turchetti, B. Psychrophilic yeasts from worldwide glacial habitats: Diversity, adaptation strategies and biotechnological potential. FEMS Microbiol. Ecol. 2012, 82, 217–241. [Google Scholar] [CrossRef]
- Sannino, C.; Tasselli, G.; Filippucci, S.; Turchetti, B.; Buzzini, P. Yeasts in nonpolar cold habitats. In Yeasts in Natural Ecosystems: Diversity; Springer: Cham, Switzerland, 2017; ISBN 9783319626833. [Google Scholar]
- Khroustalyova, G.; Giovannitti, G.; Severini, D.; Scherbaka, R.; Turchetti, B.; Buzzini, P.; Rapoport, A. Anhydrobiosis in yeasts: Psychrotolerant yeasts are highly resistant to dehydration. Yeast 2019, 36, 375–379. [Google Scholar] [CrossRef]
- Pulschen, A.A.; de Araujo, G.G.; Souza Ramos de Carvalho, A.C.; Cerini, M.F.; de Fonseca, L.M.; Galante, D.; Rodrigues, F. Survival of extremophilic yeasts in the stratospheric environment during balloon flights and in laboratory simulations. Appl. Environ. Microbiol. 2018, 84, e01942-18. [Google Scholar] [CrossRef]
- Wilhelm, L.; Singer, G.A.; Fasching, C.; Battin, T.J.; Besemer, K. Microbial biodiversity in glacier-fed streams. ISME J. 2013, 7, 1651–1660. [Google Scholar] [CrossRef]
- Bajerski, F.; Ganzert, L.; Mangelsdorf, K.; Lipski, A.; Busse, H.J.; Padur, L.; Wagner, D. Herbaspirillum psychrotolerans sp. nov., a member of the family Oxalobacteraceae from a glacier forefield. Int. J. Syst. Evol. Microbiol. 2013, 63, 3197–3203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.C.; Redzic, M.; Schinner, F.; Margesin, R. Glaciimonas immobilis gen. nov., sp. nov., a member of the family Oxalobacteraceae isolated from alpine glacier cryoconite. Int. J. Syst. Evol. Microbiol. 2011, 61, 2186–2190. [Google Scholar] [CrossRef]
- Vimercati, L.; Darcy, J.L.; Schmidt, S.K. The disappearing periglacial ecosystem atop Mt. Kilimanjaro supports both cosmopolitan and endemic microbial communities. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Baldani, J.I.; Rouws, L.; Cruz, L.M.; Olivares, F.L.; Schmid, M.; Hartmann, A. The family Oxalobacteraceae. In The Prokaryotes: Alphaproteobacteria and Betaproteobacteria; Springer: Heidelberg, Germany, 2014; ISBN 9783642301971. [Google Scholar]
- Cheptsov, V.S.; Vorobyova, E.A.; Manucharova, N.A.; Gorlenko, M.V.; Pavlov, A.K.; Vdovina, M.A.; Lomasov, V.N.; Bulat, S.A. 100 kGy gamma-affected microbial communities within the ancient Arctic permafrost under simulated Martian conditions. Extremophiles 2017, 21, 1057–1067. [Google Scholar] [CrossRef]
- Wierzchos, J.; DiRuggiero, J.; Vítek, P.; Artieda, O.; Souza-Egipsy, V.; Škaloud, P.; Tisza, M.J.; Davila, A.F.; Vílchez, C.; Garbayo, I.; et al. Adaptation strategies of endolithic chlorophototrophs to survive the hyperarid and extreme solar radiation environment of the Atacama Desert. Front. Microbiol. 2015, 6, 934. [Google Scholar] [CrossRef]
- Vincent, W.F. Cyanobacterial dominance in the polar regions. In The Ecology of Cyanobacteria; Potts, M., Whitton, B.A., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 321–340. [Google Scholar]
- Quesada, A.; Vincent, W.F. Cyanobacteria in the Cryosphere: Snow, Ice and Extreme Cold. In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Whitton, B., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 387–399. [Google Scholar]
- Selbmann, L.; Zucconi, L.; Isola, D.; Onofri, S. Rock black fungi: Excellence in the extremes, from the Antarctic to space. Curr. Genet. 2015, 61, 335–345. [Google Scholar] [CrossRef]
- Onofri, S.; Selbmann, L.; de Hoog, G.S.; Grube, M.; Barreca, D.; Ruisi, S.; Zucconi, L. Evolution and adaptation of fungi at boundaries of life. Adv. Space Res. 2007, 40, 1657–1664. [Google Scholar] [CrossRef]
- Onofri, S.; Barreca, D.; Selbmann, L.; Isola, D.; Rabbow, E.; Horneck, G.; de Vera, J.P.P.; Hatton, J.; Zucconi, L. Resistance of Antarctic black fungi and cryptoendolithic communities to simulated space and Martian conditions. Stud. Mycol. 2008, 61, 99–109. [Google Scholar] [CrossRef]
- Onofri, S.; De La Torre, R.; De Vera, J.P.; Ott, S.; Zucconi, L.; Selbmann, L.; Scalzi, G.; Venkateswaran, K.J.; Rabbow, E.; Sánchez Iñigo, F.J.; et al. Survival of rock-colonizing organisms after 1.5 years in outer space. Astrobiology 2012, 12, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Selbmann, L.; De Hoog, G.S.; Mazzaglia, A.; Friedmann, E.I.; Onofri, S. Fungi at the edge of life: Cryptoendolithic black fungi from Antarctic desert. Stud. Mycol. 2005, 51, 1–32. [Google Scholar]
- Ruibal, C.; Gueidan, C.; Selbmann, L.; Gorbushina, A.A.; Crous, P.W.; Groenewald, J.Z.; Muggia, L.; Grube, M.; Isola, D.; Schoch, C.L.; et al. Phylogeny of rock-inhabiting fungi related to Dothideomycetes. Stud. Mycol. 2009, 64, 123–133. [Google Scholar] [CrossRef]
- Selbmann, L.; De Hoog, G.S.; Zucconi, L.; Isola, D.; Onofri, S. Black yeasts in cold habitats. In Cold-Adapted Yeasts: Biodiversity, Adaptation Strategies and Biotechnological Significance; Springer: Berlin/Heidelberg, Germany, 2014; pp. 173–189. ISBN 9783642396816. [Google Scholar]
- Goto, S.; Sugiyama, J.; Iizuka, H. A taxonomic study of Antarctic yeasts. Mycologia 1969, 61, 748–774. [Google Scholar] [CrossRef] [PubMed]
- Perfumo, A.; Banat, I.M.; Marchant, R. Going Green and Cold: Biosurfactants from Low-Temperature Environments to Biotechnology Applications. Trends Biotechnol. 2018, 36, 277–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewert, M.; Deming, J.W. Sea ice microorganisms: Environmental constraints and extracellular responses. Biology 2013, 2, 603–628. [Google Scholar] [CrossRef] [PubMed]
- Belnap, J.; Weber, B.; Büdel, B. Biological Soil Crusts as an Organizing Principle in Drylands. In Biological Soil Crusts: An Organizing Principle in Drylands; Springer: Cham, Switzerland, 2016; pp. 3–13. [Google Scholar]
- Hanagata, N.; Dubinsky, Z. Secondary carotenoid accumulation in Scenedesmus komarekii (Chlorophyceae, Chlorophyta). J. Phycol. 1999, 35, 960–966. [Google Scholar] [CrossRef]
- Orosa, M.; Torres, E.; Fidalgo, P.; Abalde, J. Production and analysis of secondary carotenoids in green algae. J. Appl. Phycol. 2000, 12, 553–556. [Google Scholar] [CrossRef]
- Orosa, M.; Valero, J.F.; Herrero, C.; Abalde, J. Comparison of the accumulation of astaxanthin in Haematococcus pluvialis and other green microalgae under N-starvation and high light conditions. Biotechnol. Lett. 2001, 23, 1079–1085. [Google Scholar] [CrossRef] [Green Version]
- Pushkareva, E.; Johansen, J.R.; Elster, J. A review of the ecology, ecophysiology and biodiversity of microalgae in Arctic soil crusts. Polar Biol. 2016, 39, 2227–2240. [Google Scholar] [CrossRef]
- Palmer, R.J.; Friedmann, E.I. Water relations and photosynthesis in the cryptoendolithic microbial habitat of hot and cold deserts. Microb. Ecol. 1990, 19, 111–118. [Google Scholar] [CrossRef]
- King, A.J.; Freeman, K.R.; McCormick, K.F.; Lynch, R.C.; Lozupone, C.A.; Knight, R.; Schmidt, S.K. Biogeography and habitat modelling of high-alpine bacteria. Nat. Commun. 2010, 1, 53. [Google Scholar] [CrossRef] [Green Version]
- Fairén, A.G.; Davila, A.F.; Lim, D.; Bramall, N.; Bonaccorsi, R.; Zavaleta, J.; Uceda, E.R.; Stoker, C.; Wierzchos, J.; Dohm, J.M.; et al. Astrobiology through the ages of Mars: The study of terrestrial analogues to understand the habitability of Mars. Astrobiology 2010, 10, 821–843. [Google Scholar] [CrossRef]
- Schulze-Makuch, D.; Wagner, D.; Kounaves, S.P.; Mangelsdorf, K.; Devine, K.G.; De Vera, J.P.; Schmitt-Kopplin, P.; Grossart, H.P.; Parro, V.; Kaupenjohann, M.; et al. Transitory microbial habitat in the hyperarid Atacama Desert. Proc. Natl. Acad. Sci. USA 2018, 115, 2670–2675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Des Marais, D.J.; Nuth, J.A.; Allamandola, L.J.; Boss, A.P.; Farmer, J.D.; Hoehler, T.M.; Jakosky, B.M.; Meadows, V.S.; Pohorille, A.; Runnegar, B.; et al. The NASA Astrobiology Roadmap. Astrobiology 2008, 8, 715–730. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comparison | OTU ID | Phylum | Family | % | Cumulative |
|---|---|---|---|---|---|
| Contribution | % | ||||
| Start—Water | denovo6723 | Betaproteobacteria | Oxalobacteraceae | 7.08 | 7.08 |
| Mean = 35% | denovo7247 | Actinobacteria | Pseudonocardiaceae | 3.58 | 10.66 |
| denovo3383 | Actinobacteria | Solirubrobacterales | 2.82 | 13.48 | |
| Control—Water | denovo6723 | Betaproteobacteria | Oxalobacteraceae | 6.82 | 6.82 |
| Mean = 36% | denovo3383 | Actinobacteria | Solirubrobacterales | 3.87 | 10.69 |
| denovo7247 | Actinobacteria | Pseudonocardiaceae | 3.42 | 14.11 | |
| Start—Water + Nutrients | denovo7247 | Actinobacteria | Pseudonocardiaceae | 3.57 | 3.57 |
| Mean = 36% | denovo11840 | Verrucomicrobia | Chthoniobacteraceae | 3.00 | 6.56 |
| denovo8881 | Actinobacteria | Phycicoccus | 2.65 | 9.22 | |
| Control—Water + Nutrients | denovo7247 | Actinobacteria | Pseudonocardiaceae | 3.56 | 3.56 |
| Mean = 35% | denovo7213 | Bacteroidetes | Chitinophagaceae | 2.89 | 6.45 |
| denovo15971 | Bacteroidetes | Chitinophagaceae | 2.86 | 9.31 | |
| Water–Water + Nutrients | denovo6723 | Betaproteobacteria | Oxalobacteraceae | 7.01 | 7.01 |
| Mean = 37% | denovo3383 | Actinobacteria | Solirubrobacterales | 4.72 | 11.73 |
| denovo11840 | Verrucomicrobia | Chthoniobacteraceae | 2.55 | 14.28 |
| Comparison | OTU ID | Phylum | Closest Taxonomy | % | Cumulative |
|---|---|---|---|---|---|
| Contribution | % | ||||
| Start—Water | denovo259 | Basidiomycota | Moeszyomyces sp. | 16.76 | 16.76 |
| Mean = 68% | denovo1264 | Basidiomycota | Naganishia friedmannii | 15.27 | 32.03 |
| denovo251 | Chlorophyta | Neochlorosarcina sp. | 10.29 | 42.32 | |
| Control—Water | denovo259 | Basidiomycota | Moeszyomyces sp. | 18.29 | 18.29 |
| Mean = 63% | denovo1264 | Basidiomycota | Naganishia friedmannii | 17.16 | 35.45 |
| denovo251 | Chlorophyta | Neochlorosarcina sp. | 11.39 | 46.84 | |
| Start—Water + Nutrients | denovo1264 | Basidiomycota | Naganishia friedmannii | 29.33 | 29.33 |
| Mean = 59% | denovo1929 | Dothideomycetes | Pleosporales (order) | 9.90 | 39.23 |
| denovo264 | Chlorophyta | Chlamydomonas sp. | 6.11 | 45.34 | |
| Control—Water + Nutrients | denovo1264 | Basidiomycota | Naganishia friedmannii | 28.04 | 28.04 |
| Mean = 52% | denovo1929 | Dothideomycetes | Pleosporales (order) | 10.27 | 38.31 |
| denovo598 | Cercozoa | Thecophilosea sp. | 9.53 | 47.84 | |
| Water–Water + Nutrients | denovo1264 | Basidiomycota | Naganishia friedmannii | 36.05 | 36.05 |
| Mean = 59% | denovo259 | Basidiomycota | Moeszyomyces sp. | 17.79 | 53.84 |
| denovo251 | Chlorophyta | Neochlorosarcina sp. | 12.24 | 66.08 |
| Taxon | Organism Type | + Water | + Water + Nutrients | Increase Rate | Strategy | Mechanisms of Stress Resistance |
|---|---|---|---|---|---|---|
| N. friedmannii | Basidiomycete yeast | Neutral | Increase | Fast | r-selected | Dormancy |
| Moesziomyces sp. | Basidiomycete fungus | Increase | Increase | Slow | K-selected | Cold-active enzymes, biosurfactants |
| Dothideomycetes | Ascomycete fungus | Increase | Increase | Slow | K-selected | Extracellular polymeric substances, melanin, compatible solutes |
| Neochlorosarcina sp. | Alga | Increase | Neutral | Slow | K-selected | Extracellular polymeric substances, carotenoids, dormancy, motility |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vimercati, L.; Bueno de Mesquita, C.P.; Schmidt, S.K. Limited Response of Indigenous Microbes to Water and Nutrient Pulses in High-Elevation Atacama Soils: Implications for the Cold–Dry Limits of Life on Earth. Microorganisms 2020, 8, 1061. https://doi.org/10.3390/microorganisms8071061
Vimercati L, Bueno de Mesquita CP, Schmidt SK. Limited Response of Indigenous Microbes to Water and Nutrient Pulses in High-Elevation Atacama Soils: Implications for the Cold–Dry Limits of Life on Earth. Microorganisms. 2020; 8(7):1061. https://doi.org/10.3390/microorganisms8071061
Chicago/Turabian StyleVimercati, Lara, Clifton P. Bueno de Mesquita, and Steven K. Schmidt. 2020. "Limited Response of Indigenous Microbes to Water and Nutrient Pulses in High-Elevation Atacama Soils: Implications for the Cold–Dry Limits of Life on Earth" Microorganisms 8, no. 7: 1061. https://doi.org/10.3390/microorganisms8071061
APA StyleVimercati, L., Bueno de Mesquita, C. P., & Schmidt, S. K. (2020). Limited Response of Indigenous Microbes to Water and Nutrient Pulses in High-Elevation Atacama Soils: Implications for the Cold–Dry Limits of Life on Earth. Microorganisms, 8(7), 1061. https://doi.org/10.3390/microorganisms8071061