Anti-Biofilm Activity of a Low Weight Proteinaceous Molecule from the Marine Bacterium Pseudoalteromonas sp. IIIA004 against Marine Bacteria and Human Pathogen Biofilms

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Preparation of the Pseudoalteromonas sp. IIIA004 Supernatant (SNIIIA004)
2.3. Anti-Biofilm Assays
2.4. Antibacterial Assays
2.5. Microbial Adhesion to Solvent (MATS) Assays
2.6. Physico-Chemical Characterization of the Active Compound(s)
2.7. Biosurfactant Assay
2.8. Purification of the SNIIIA004 Anti-Biofilm Compound
2.9. Statistical Analyses
3. Results
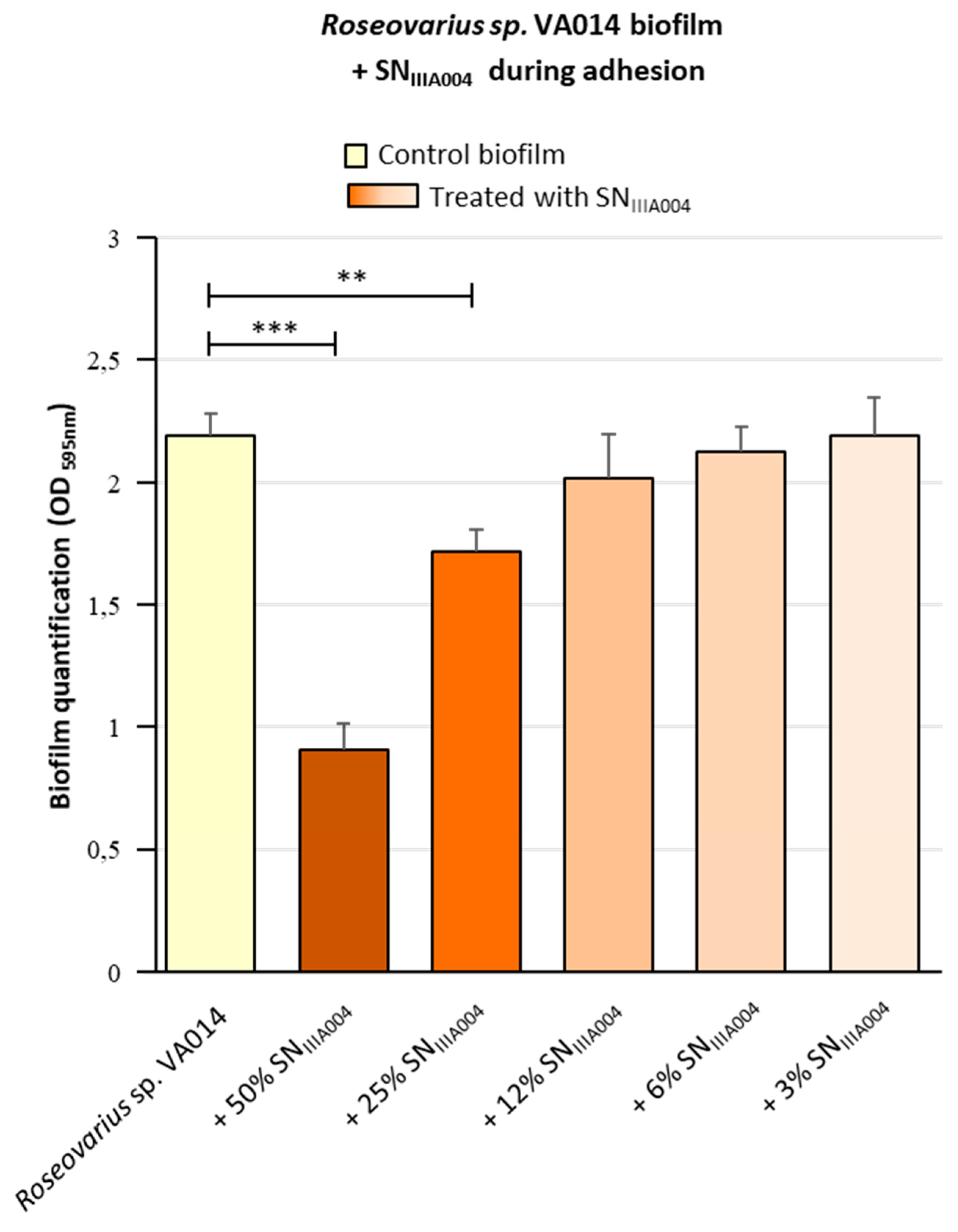
3.1. Pseudoalteromonas sp. IIIA004 Exoproducts Inhibit Biofilm Formation by Roseovarius sp. VA014
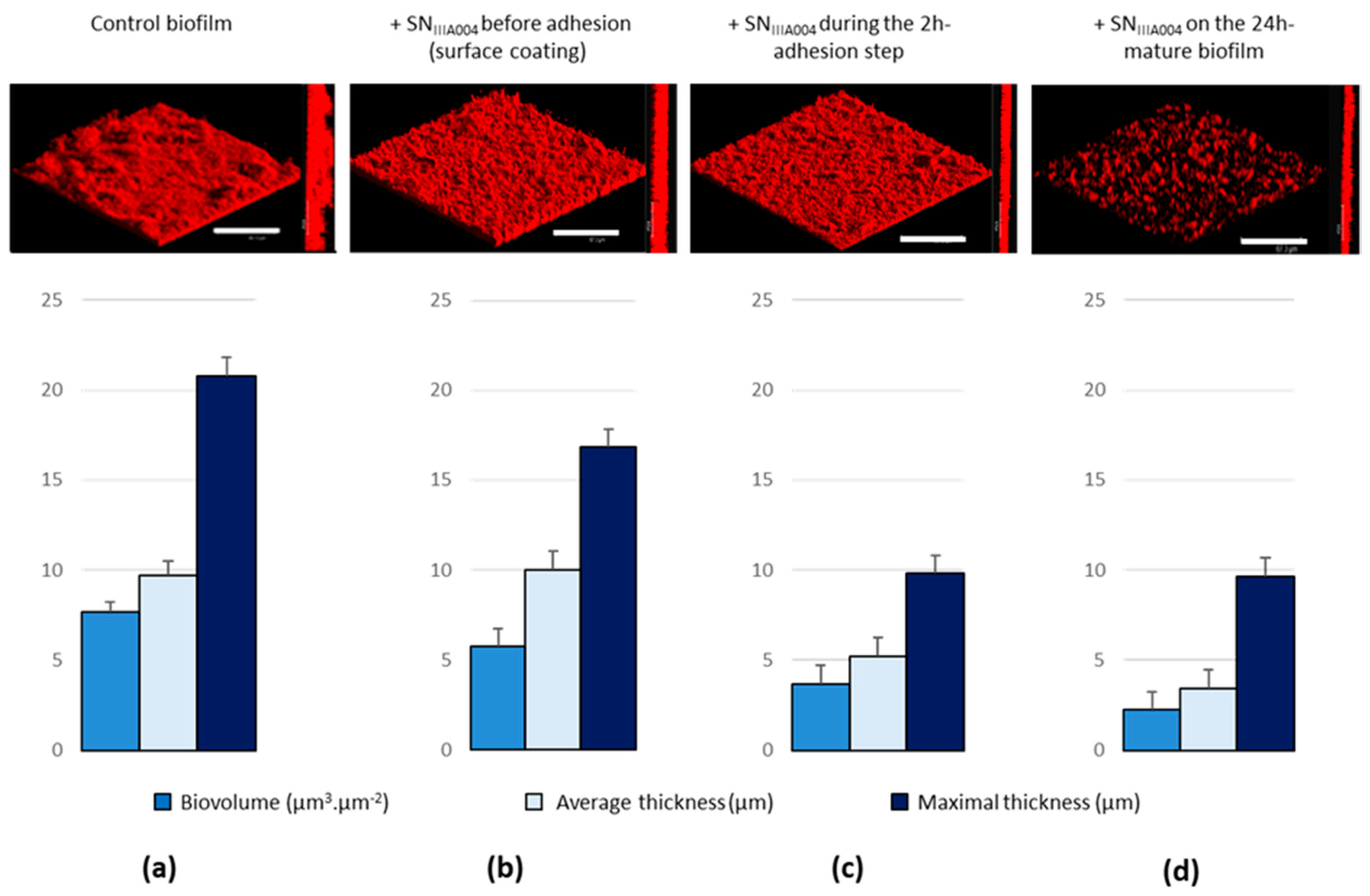
3.2. SNIIIA004 Disrupts the Established Bacterial Biofilm
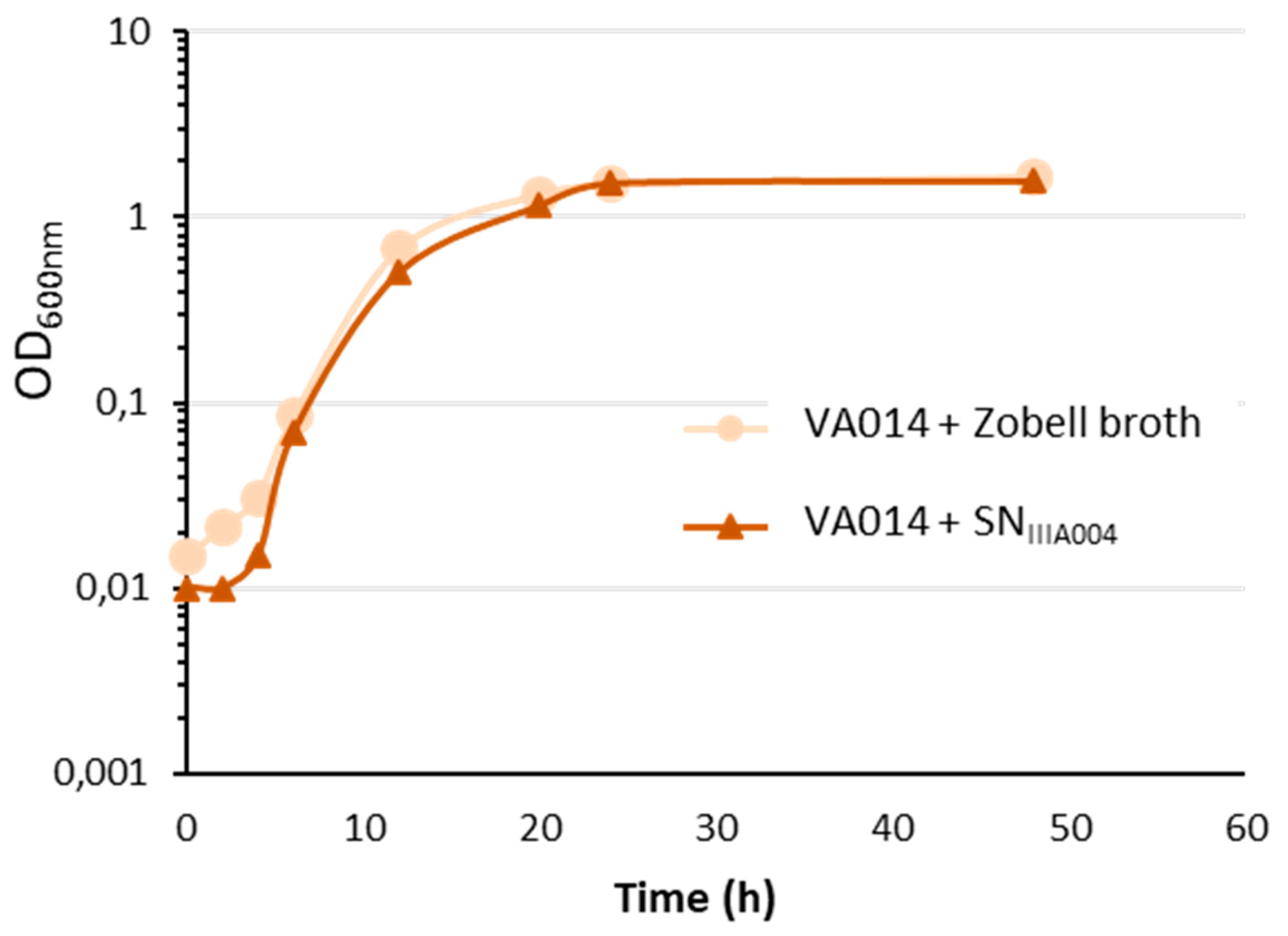
3.3. SNIIIA004 is Devoid of Bactericidal Activity against Free-Living Cells
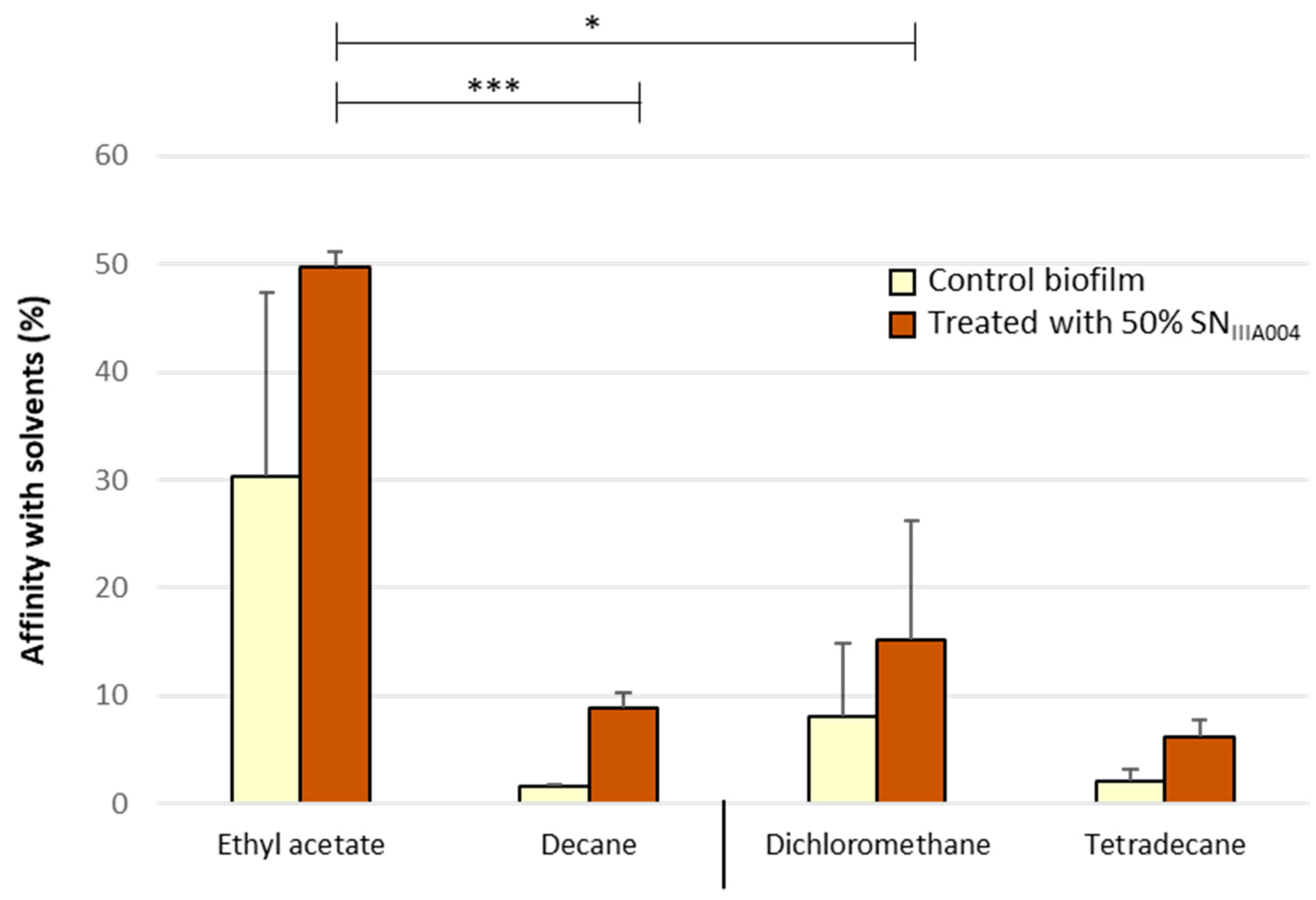
3.4. SNIIIA004 Does Not Change the Hydrophilic and Acid Character of the Roseovarius sp. VA014 Cell Surface
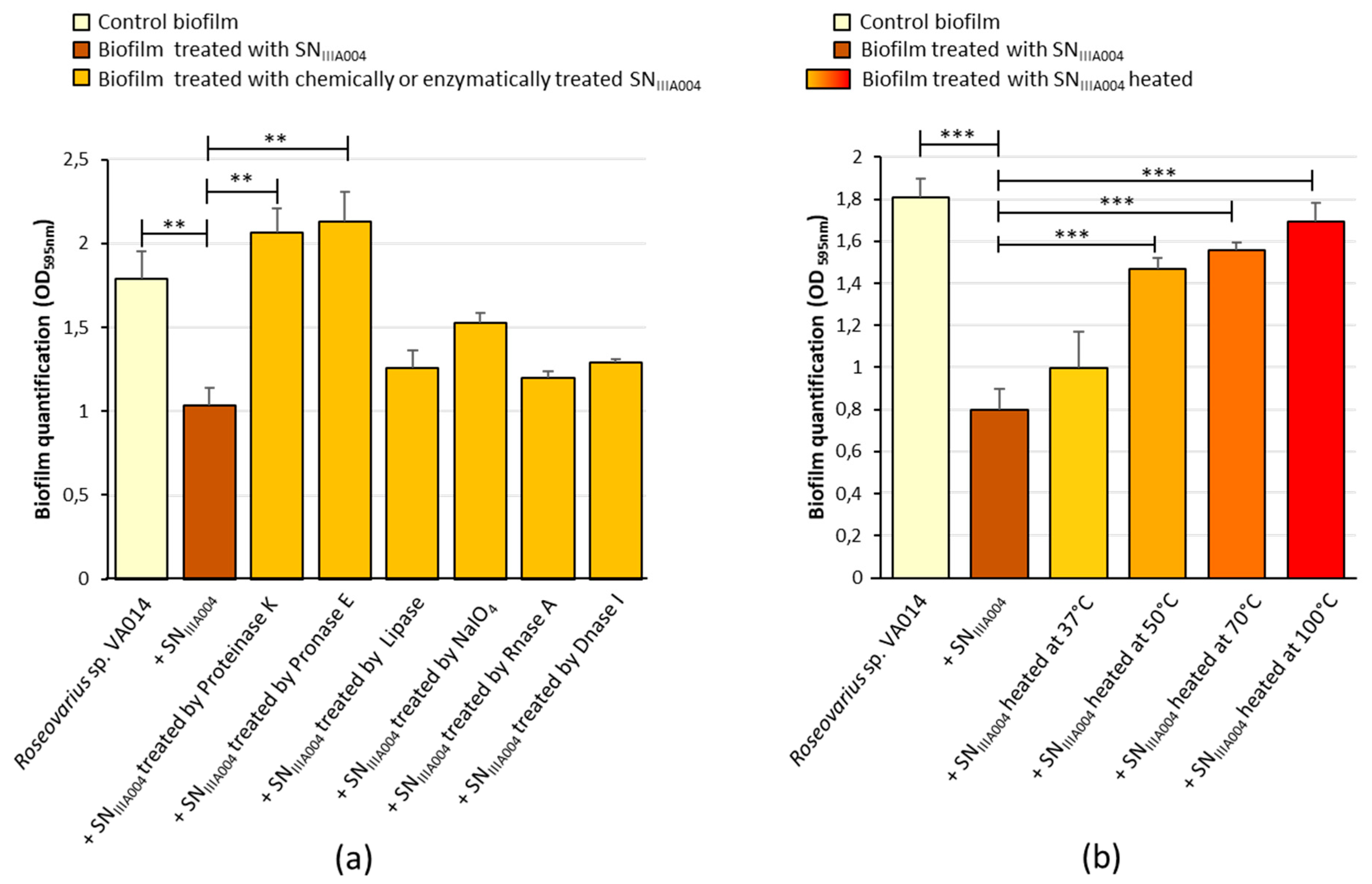
3.5. Physico-Chemical Characteristics of SNIIIA004 Anti-Biofilm Compounds
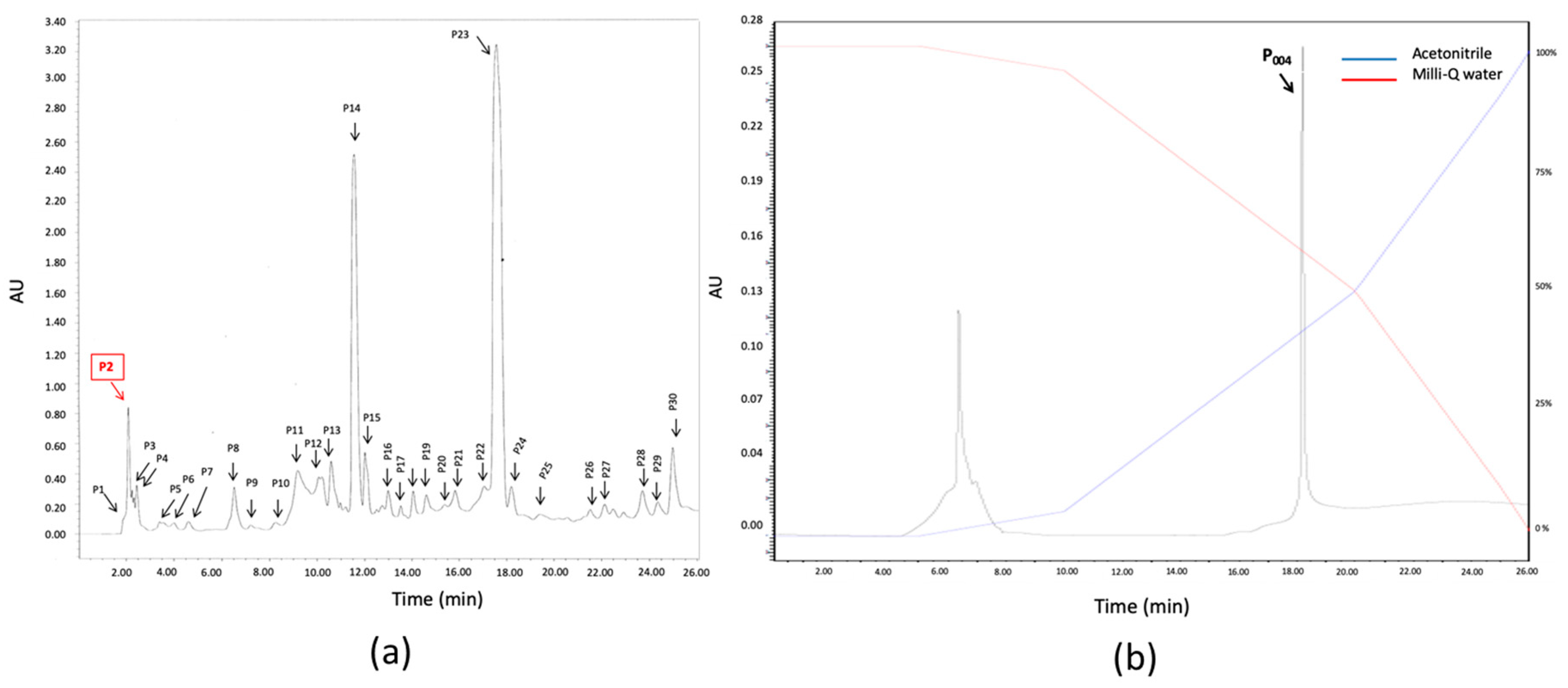
3.6. Isolation and Purification of the Anti-Biofilm Molecule
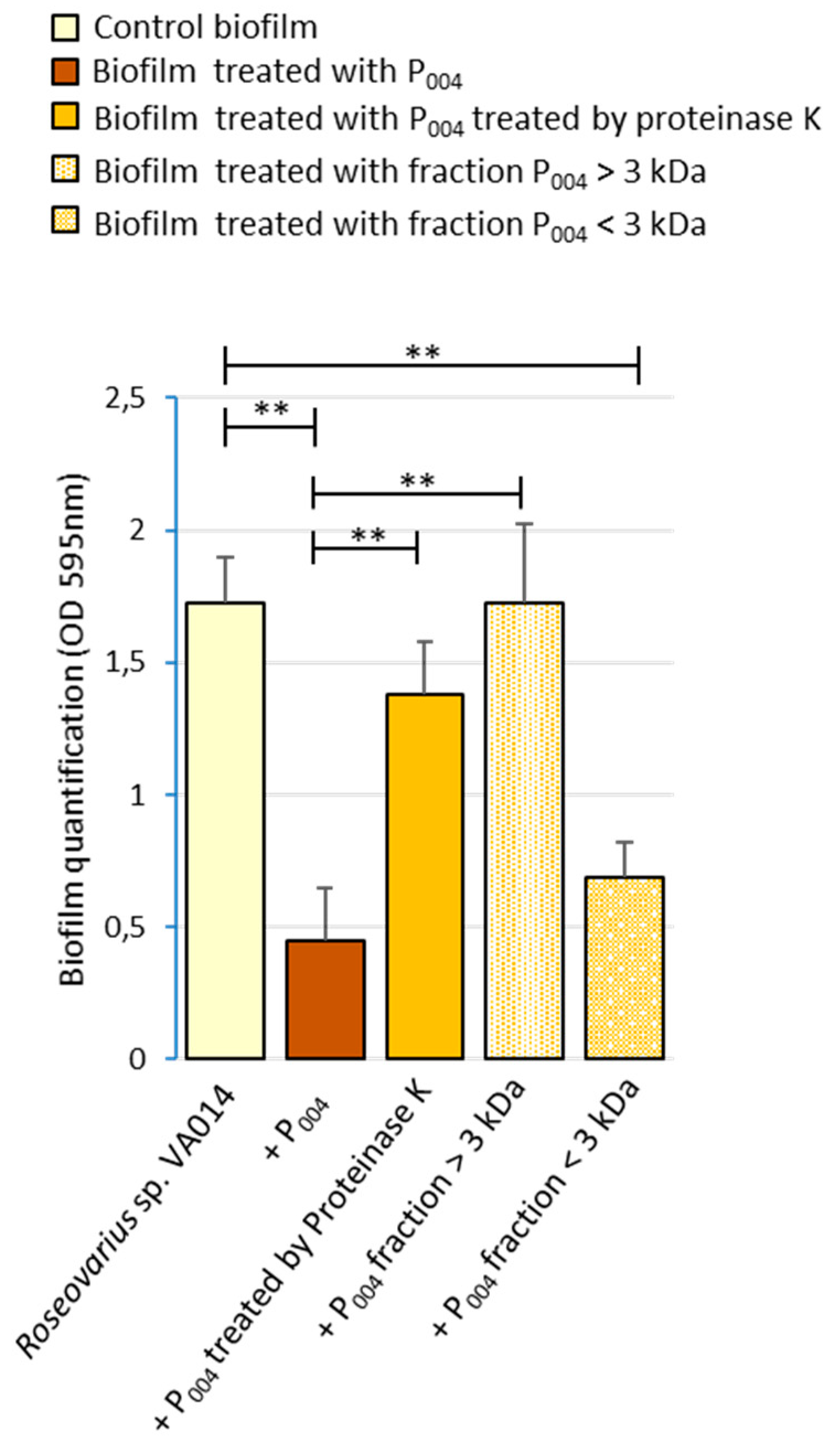
3.7. Spectrum of Action of the P004 Anti-Biofilm Molecule Purified from SNIIIA004
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Wang, Y.; Liu, L.; He, J. Biofilms: The Microbial “Protective Clothing” in Extreme Environments. Int. J. Mol. Sci. 2019, 20, 3423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francolini, I.; Donelli, G. Prevention of biofilm-based medical-device related infections. FEMS Immunol. Med. Microbiol. 2010, 59, 227–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lequette, Y.; Boels, G.; Clarisse, M.; Faille, C. Using enzymes to remove biofilms of bacterial isolates sampled in the food-industry. Biofouling 2010, 26, 421–431. [Google Scholar] [CrossRef]
- Subhadra, B.; Kim, D.H.; Woo, K.; Surendran, S.; Choi, C.H. Control of Biofilm Formation in Healthcare: Recent Advances Exploiting Quorum-Sensing Interference Strategies and Multidrug Efflux Pump Inhibitors. Materials 2018, 11, 1676. [Google Scholar] [CrossRef] [Green Version]
- Shikuma, N.J.; Hadfield, M.G. Marine biofilms on submerged surfaces are a reservoir for Escherichia coli and Vibrio cholera. Biofouling 2010, 26, 39–46. [Google Scholar] [CrossRef]
- Lanneluc, I.; Langumier, M.; Sabot, R.; Jeannin, M.; Refait, P.; Sablé, S. On the bacterial communities associated with the corrosion product layer during the early stages of marine corrosion of carbon steel. Int. Biodeterior. Biodegrad. 2015, 99, 55–65. [Google Scholar] [CrossRef]
- Schultz, M.P.; Bendick, J.A.; Holm, E.R.; Hertel, W.M. Economic impact of biofouling on a naval surface ship. Biofouling 2011, 27, 87–98. [Google Scholar] [CrossRef]
- Beloin, C.; Renard, S.; Ghigo, J.M.; Lebeaux, D. Novel approaches to combat bacterial biofilms. Curr. Opin. Pharm. 2014, 18, 61–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Norcy, T.; Niemann, H.; Proksch, P.; Linossier, I.; Vallée-Réhel, K.; Hellio, C.; Faÿ, F. Anti-Biofilm Effect of Biodegradable Coatings Based on Hemibastadin Derivative in Marine Environment. Int. J. Mol. Sci. 2017, 18, 1520. [Google Scholar] [CrossRef] [PubMed]
- Buommino, E.; Scognamiglio, M.; Donnarumma, G.; Fiorentino, A.; D’Abrosca, B. Recent advances in natural product-based anti-biofilm approaches to control infections. Mini. Rev. Med. Chem. 2014, 14, 1169–1182. [Google Scholar] [CrossRef]
- Kaplan, J.B. Biofilm dispersal: Mechanisms, clinical implications and potential therapeutic uses. J. Dent. Res. 2010, 89, 205–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daboor, S.M.; Raudonis, R.; Cohen, A.; Rohde, J.R.; Cheng, Z. Marine bacteria, a source for alginolytic enzyme to disrupt Pseudomonas aeruginosa biofilms. Mar. Drugs 2019, 17, 307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobretsov, S.; Dahms, H.U.; Qian, P.Y. Inhibition of biofouling by marine microorganisms and their metabolites. Biofouling 2006, 22, 43–54. [Google Scholar] [CrossRef]
- Estrela, A.B.; Heck, M.G.; Abraham, W.R. Novel approaches to control biofilm infections. Curr. Med. Chem. 2009, 16, 1512–1530. [Google Scholar] [CrossRef]
- Klein, G.L.; Soum-Soutera, E.; Guede, Z.; Bazire, A.; Compere, C.; Dufour, A. The anti-biofilm activity secreted by a marine Pseudoalteromonas strain. Biofouling 2011, 27, 931–940. [Google Scholar] [CrossRef]
- Dobretsov, S.; Abed, R.M.M.; Teplitski, M. Mini-review: Inhibition of biofouling by marine microorganisms. Biofouling 2013, 29, 423–441. [Google Scholar] [CrossRef]
- Papa, R.; Parrilli, E.; Sannino, F.; Barbato, G.; Tutino, M.L.; Artini, M.; Selan, L. Anti-biofilm activity of the Antarctic marine bacterium Pseudoalteromonas haloplanktis TAC125. Res. Microbiol. 2013, 164, 450–456. [Google Scholar] [CrossRef]
- Parrilli, E.; Ricciardelli, A.; Casillo, A.; Sannino, F.; Papa, R.; Tilotta, M.; Artini, M.; Selan, L.; Corsaro, M.M.; Tutino, M.L. Large-scale biofilm cultivation of Antarctic bacterium Pseudoalteromonas haloplanktis TAC125 for physiologic studies and drug discovery. Extremophiles 2016, 20, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.L.; Wu, Z.H.; Wang, Y.; Wang, C.Y.; Xu, Y. Mini-Review: Antifouling Natural Products from Marine Microorganisms and Their Synthetic Analogs. Mar. Drugs 2017, 15, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desriac, F.; Rodrigues, S.; Doghri, I.; Sablé, S.; Lanneluc, I.; Fleury, Y.; Bazire, A.; Dufour, A. Antimicrobial and antibiofilm molecules produced by marine bacteria. In Blue Biotechnology: Production and Use of Marine Molecules; Wiley: Hoboken, NJ, USA, 2018; ISBN 978-3-527-34138-2. [Google Scholar]
- Holmstrom, C.; Kjelleberg, S. Marine Pseudoalteromonas species are associated with higher organisms and produce active extracellular agents. Fems Microbiol. Ecol. 1999, 30, 285–293. [Google Scholar] [CrossRef]
- Bowman, J.P. Bioactive compound synthetic capacity and ecological significance of marine bacterial genus Pseudoalteromonas. Mar. Drugs 2007, 5, 220–241. [Google Scholar] [CrossRef]
- Grasland, B.; Mitalane, J.; Briandet, R.; Quemener, E.; Meylheuc, T.; Linossier, I.; Vallee-Rehel, K.; Haras, D. Bacterial Biofilm in Seawater: Cell Surface Properties of Early-attached Marine Bacteria. Biofouling 2003, 19, 307–313. [Google Scholar] [CrossRef]
- Pradier, C.M.; Rubio, C.; Poleunis, C.; Bertrand, P.; Marcus, P.; Compère, C. Surface characterization of three marine bacterial strains by Fourier transform IR, X-ray photoelectron spectroscopy, and time-of-flight secondary-ion mass spectrometry, correlation with adhesion on stainless steel surfaces. J. Phys. Chem. 2005, 109, 9540–9549. [Google Scholar] [CrossRef]
- Dheilly, A.; Soum-Soutera, E.; Klein, G.L.; Bazire, A.; Compere, C.; Haras, D.; Dufour, A. Anti-biofilm activity of the marine bacterium Pseudoalteromonas sp. strain 3J6. Appl. Environ. Microbiol. 2010, 76, 3452–3461. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, S.; Paillard, C.; Dufour, A.; Bazire, A. Antibiofilm activity of the marine bacterium Pseudoalteromonas sp. 3J6 against Vibrio tapetis, the causative agent of Brown Ring Disease. Probiotics Antimicrob. Proteins 2015, 27, 45–51. [Google Scholar] [CrossRef]
- Casillo, A.; Papa, R.; Ricciardelli, A.; Sannino, F.; Ziaco, M.; Tilotta, M.; Selan, L.; Marino, G.; Corsaro, M.M.; Tutino, M.L.; et al. Anti-Biofilm Activity of a Long-Chain Fatty Aldehyde from Antarctic Pseudoalteromonas haloplanktis TAC125 against Staphylococcus epidermidis Biofilm. Front. Cell Infect. Microbiol. 2017, 7, 46. [Google Scholar] [CrossRef] [Green Version]
- Sulieman, F.; Ahmad, A.; Usup, G.; Kuang, L.C. Diketopiperazine from marine bacterium Pseudoalteromonas ruthenica KLPp3. J. Biol. Res. 2018, 91, 7197. [Google Scholar] [CrossRef] [Green Version]
- Doghri, I.; Rodrigues, S.; Bazire, A.; Dufour, A.; Akbar, D.; Sopena, V.; Sablé, S.; Lanneluc, I. Marine bacteria from the French Atlantic coast displaying high forming-biofilm abilities and different biofilm 3D architectures. BMC Microbiol. 2015, 15, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, H.; Chen, R.; Wang, L.; Shao, S.; Dai, L.; Ye, Y.; Guo, L.; Huang, G.; Klotz, M.G. Molecular characterization of putative biocorroding microbiota with a novel niche detection of Epsilon- and Zetaproteobacteria in Pacific Ocean coastal seawaters. Environ. Microbiol. 2011, 13, 3059–3074. [Google Scholar] [CrossRef] [PubMed]
- Malone, C.L.; Boles, B.R.; Lauderdale, K.J.; Thoendel, M.; Kavanaugh, J.S.; Horswill, A.R. Fluorescent reporters for Staphylococcus aureus. J. Microbiol. Methods 2009, 77, 251–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, D.; Memmi, G.; Hernandez, D.; Bard, J.; Beaume, M.; Gill, S.; Francois, P.; Cheung, A.L. Whole-genome sequencing of Staphylococcus aureus strain RN4220, a key laboratory strain used in virulence research, identifies mutations that affect not only virulence factors but also the fitness of the strain. J. Bacteriol. 2011, 193, 2332–2335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klausen, M.; Heydorn, A.; Ragas, P.; Lambertsen, L.; Aaes-Jørgensen, A.; Molin, S.; Tolker-Nielsen, T. Biofilm formation by Pseudomonas aeruginosa wild type, flagella and type IV pili mutants. Mol. Microbiol. 2003, 48, 1511–1524. [Google Scholar] [CrossRef]
- He, J.; Baldini, R.L.; Déziel, E.; Saucier, M.; Zhang, Q.; Liberati, N.T.; Lee, D.; Urbach, J.; Goodman, H.M.; Rahme, L.G. The broad host range pathogen Pseudomonas aeruginosa strain PA14 carries two pathogenicity islands harboring plant and animal virulence genes. Proc. Natl. Acad. Sci. USA 2004, 101, 2530–2535. [Google Scholar] [CrossRef] [Green Version]
- Bridier, A.; Le Coq, D.; Dubois-Brissonnet, F.; Thomas, V.; Aymerich, S.; Briandet, R. The spatial architecture of Bacillus subtilis biofilms deciphered using a surface-associated model and in situ imaging. PLoS ONE 2011, 6, e16177. [Google Scholar] [CrossRef] [Green Version]
- Sheppard, A.E.; Poehlein, A.; Rosenstiel, P.; Liesegang, H.; Schulenburg, H. Complete Genome Sequence of Bacillus thuringiensis Strain 407 Cry-. Genome Announc. 2013, 1, e00158-12. [Google Scholar] [CrossRef] [Green Version]
- Heydorn, A.; Nielsen, A.T.; Hentzer, M.; Sternberg, C.; Givskov, M.; Ersbøll, B.K.; Molin, S. Quantification of biofilm structures by the novel computer program COMSTAT. Microbiol. 2000, 146, 2395–2407. [Google Scholar] [CrossRef] [Green Version]
- Sablé, S.; Pons, A.-M.; Gendron-Gaillard, S.; Cottenceau, G. Antibacterial activity evaluation of microcin J25 against diarrheagenic Escherichia coli. Appl. Environ. Microbiol. 2000, 66, 4595–4597. [Google Scholar] [CrossRef] [Green Version]
- Bellon-Fontaine, M.N.; Rault, J.; van Oss, C.J. Microbial adhesion to solvents: A novel method to determine the electron-donor/electron-acceptor or Lewis acid–base properties of microbial cells. Colloids Surf. B Biointerfaces 1996, 7, 47–53. [Google Scholar] [CrossRef]
- Mack, D.; Nedelmann, M.; Krokotsch, A.; Schwarzkopf, A.; Heesemann, J.; Laufs, R. Characterization of transposon mutants of biofilm-producing Staphylococcus epidermidis impaired in the accumulative phase of biofilm production: Genetic identification of a hexosamine-containing polysaccharide intercellular adhesin. Infect. Immun. 1994, 62, 3244–3253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mack, D.; Fischer, W.; Krokotsch, A.; Leopold, K.; Hartmann, R.; Egge, H.; Laufs, R. The intercellular adhesin involved in biofilm accumulation of Staphylococcus epidermidis is a linear beta-1,6-linked glucosaminoglycan: Purification and structural analysis. J. Bacteriol. 1996, 178, 175–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babor, K.; Kaláč, V.; Tihlárik, K. Periodate oxidation of saccharides. III. Comparison of the methods for determining the consumption of sodium periodate and the amount of formic acid formed. Chem. Zvesti. 1973, 27, 676–680. [Google Scholar]
- Tugrul, T.; Cansunar, E. Detecting surfactant-producing microorganisms by the drop-collapse test. World J. Microb. Biot. 2005, 21, 851–853. [Google Scholar] [CrossRef]
- Rendueles, O.; Kaplan, J.B.; Ghigo, J.M. Antibiofilm polysaccharides. Environ. Microbiol. 2013, 15, 334–346. [Google Scholar] [CrossRef]
- Wu, S.; Liu, G.; Jin, W.; Xiu, P.; Sun, C. Antibiofilm and anti-infection of a marine bacterial exopolysaccharide against Pseudomonas aeruginosa. Front. Microbiol. 2016, 7, 102. [Google Scholar] [CrossRef] [Green Version]
- Costerton, J.W.; Stewart, P.S. Battling biofilms the war is against bacterial colonies that cause some of the most tenacious infections known. The weapon is knowledge of the enemy’s communication system. Sci. Am. 2001, 285, 74–81. [Google Scholar] [CrossRef]
- Zhang, A.; Mu, H.; Zhang, W.; Cui, G.; Zhu, J.; Duan, J. Chitosan Coupling Makes Microbial Biofilms Susceptible to Antibiotics. Sci. Rep. 2013, 3, 3364. [Google Scholar] [CrossRef] [Green Version]
- Meira Ribeiro, S.; de la Fuente-Núñez, C.; Baquir, B.; Faria-Junior, C.; Franco, O.L.; Hancock, R.E.W. Antibiofilm peptides increase the susceptibility of carbapenemase-producing Klebsiella pneumoniae clinical isolates to ß-lactam antibiotics. Antimicrob. Agents Chemother. 2015, 59, 3906–3912. [Google Scholar] [CrossRef] [Green Version]
- Mai-Prochnow, A.; Evans, F.; Dalisay-Saludes, D.; Stelzer, S.; Egan, S.; James, S.; Webb, J.S.; Kjelleberg, S. Biofilm development and cell death in the marine bacterium Pseudoalteromonas tunicata. Appl. Environ. Microbiol. 2004, 70, 3232–3238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supardy, N.A.; Ibrahim, D.; Mat Nor, S.R.; Noordin, W.N.M. Bioactive Compounds of Pseudoalteromonas sp. IBRL PD4.8 Inhibit Growth of Fouling Bacteria and Attenuate Biofilms of Vibrio alginolyticus FB3. Pol. J. Microbiol. 2019, 68, 21–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, P.; Li, J.; Han, F.; Duan, G.; Lu, X.; Gu, Y.; Yu, W. Anti-biofilm Activity of an exopolysaccharide from Marine Bacterium Vibrio sp. QY101. PLoS ONE 2011, 6, e18514. [Google Scholar] [CrossRef] [Green Version]
- Das, P.; Mukherjee, S.; Sen, R. Antiadhesive action of a marine microbial surfactant. Colloids Surf. B Biointerfaces 2009, 71, 183–186. [Google Scholar] [CrossRef]
- Kim, Y.; Oh, S.; Kim, S.H. Released exopolysaccharide (r-EPS) produced from probiotic bacteria reduce biofilm formation of enterohemorrhagic Escherichia coli O157:H7. Biochem. Biophys Res. Commun. 2009, 379, 324–329. [Google Scholar] [CrossRef]
- Zinger-Yosovich, K.D.; Gilboa-Garber, N. Blocking of Pseudomonas aeruginosa and Ralstonia solanacearum lectins by plant and microbial branched polysaccharides used as food additives. J. Agric. Food Chem. 2009, 57, 6908–6913. [Google Scholar] [CrossRef]
- Chaves-López, C.; Usai, D.; Donadu, M.G.; Serio, A.; González-Mina, R.T.; Simeoni, M.C.; Molicotti, P.; Zanetti, S.; Pinna, A.; Paparella, A. Potential of Borojoa patinoi Cuatrecasas water extract to inhibit nosocomial antibiotic resistant bacteria and cancer cell proliferation in vitro. Food Funct. 2018, 23, 2725–2734. [Google Scholar] [CrossRef]
- Dong, Y.H.; Gusti, A.R.; Zhang, Q.; Xu, J.L.; Zhang, L.H. Identification of quorum-quenching N-acyl homoserine lactonases from Bacillus species. Appl. Environ. Microbiol. 2002, 68, 1754–1759. [Google Scholar] [CrossRef] [Green Version]
- Romero, M.; Diggle, S.P.; Heeb, S.; Camara, M.; Otero, A. Quorum quenching activity in Anabaena sp. PCC7120: Identification of AiiC, a novel AHL-acylase. Fems. Microbiol. Lett. 2008, 280, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Hymes, S.R.; Randis, T.M.; Sun, T.Y.; Ratner, A.J. DNase inhibits Gardnerella vaginalis biofilms in vitro and in vivo. J. Infect. Dis. 2013, 207, 1491–1497. [Google Scholar] [CrossRef] [Green Version]
- Kalpana, B.J.; Aarthy, S.; Pandian, S.K. Anti-biofilm activity of alpha-amylase from Bacillus subtilis S8-18 against biofilm forming human bacterial pathogens. Appl. Biochem. Biotechnol. 2012, 167, 1778–1794. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.B.; Ragunath, C.; Velliyagounder, K.; Fine, D.H.; Ramasubbu, N. Enzymatic detachment of Staphylococcus epidermidis biofilms. Antimicrob. Agents Chemother. 2004, 48, 2633–2636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brian-Jaisson, F.; Molmeret, M.; Fahs, A.; Guentas-Dombrowsky, L.; Culioli, G.; Blache, Y.; Cérantola, S.; Ortalo-Magné, A. Characterization and anti-biofilm activity of extracellular polymeric substances produced by the marine biofilm-forming bacterium Pseudoalteromonas ulvae strain TC14. Biofouling 2016, 32, 547–560. [Google Scholar] [CrossRef] [PubMed]
- Jouault, A.; Gobet, A.; Simon, M.; Portier, E.; Perennou, M.; Corre, E.; Gaillard, F.; Vallenet, D.; Michel, G.; Fleury, Y.; et al. Alterocin, an antibiofilm protein secreted by Pseudoalteromonas sp. strain 3J6. Appl. Environ. Microbiol. 2020, 86, e00893-20. [Google Scholar] [CrossRef]
- Pletzer, D.; Hancock, R.E. Antibiofilm Peptides: Potential as Broad-Spectrum Agents. J. Bacteriol. 2016, 198, 2572–2578. [Google Scholar] [CrossRef] [Green Version]
- Yasir, M.; Willcox, M.D.P.; Dutta, D. Action of Antimicrobial Peptides against Bacterial Biofilms. Materials 2018, 11, 2468. [Google Scholar] [CrossRef] [Green Version]
- Offret, C.; Desriac, F.; Le Chevalier, P.; Mounier, J.; Jégou, C.; Fleury, Y. Spotlight on Antimicrobial Metabolites from the Marine Bacteria Pseudoalteromonas: Chemodiversity and Ecological Significance. Mar. Drugs 2016, 14, 129. [Google Scholar] [CrossRef] [Green Version]
- James, S.G.; Holmstrom, C.; Kjelleberg, S. Purification and characterization of a novel antibacterial protein from the marine bacterium D2. Appl. Environ. Microbiol. 1996, 62, 2783–2788. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Reference and/or Source | Culture Conditions |
|---|---|---|
| Non Marine Strains | ||
| Staphylococcus aureus AH478 | [34] | Luria-Bertani 37 °C |
| S. aureus ATCC27217 | ATCC | |
| S. aureus RN4220 | [35] | |
| Pseudomonas aeruginosa PAO1 | [36] | |
| P. aeruginosa PA14 | [37] | |
| Yersiniaenterocolitica CIP106.676 | CIP | |
| Bacillus subtilis ND Food | [38] | |
| Bacillus thuringiensis 407 | [39] | |
| Marine Strains | ||
| Paracoccus sp. 4M6 | [26]/Morbihan Gulf, France | Zobell 22 °C |
| Micrococcus luteus | LBCM | |
| Zobellia galactanivorans | LBCM | |
| Cellulophaga lytica DSM2039 | DSMZ | |
| Cellulophaga lytica DSM2040 | DSMZ | |
| Vibrio lentus CIP107166T | CIP | |
| Vibrio anguillarum CIP6336T | CIP | |
| Vibrio sp. D01 | [27]/Bay of Brest, France | |
| Pseudoalteromonas sp. IIIA004 | [33]/Atlantic harbor, France | |
| Roseovarius sp. VA014 | [9]/Atlantic harbor, France | |
| Roseobacter sp. IV 3009 | [32]/Intertidal mudflat, France | |
| Shewanella sp. IV 3014 | [32]/Intertidal mudflat, France | |
| Flavobacterium sp. II2003 | [32]/Intertidal mudflat, France | |
| Tenacibaculum sp. II2021 | [32]/Intertidal mudflat, France | |
| P004 (10 µg) | |
|---|---|
| Strain | Anti-biofilm Assays a |
| High inhibition level b | |
| Roseovarius sp. VA014 | 71.4 ± 2.2 |
| Staphylococcus aureus AH478 | 71.4 ± 2.2 |
| Staphylococcus aureus RN4220 | 62.2 ± 1.3 |
| Yersiniaenterocolitica CIP106.676 | 81.5 ± 5.8 |
| Paracoccus sp. 4M6 | 77.7 ± 1.9 |
| Pseudomonas aeruginosa PAO1 | 80.9 ± 2.4 |
| Micrococcus luteus | 71.7 ± 8.3 |
| Mild Inhibition Level b | |
| Staphylococcus aureus ATCC27217 | 45.1 ± 5.3 |
| Zobellia galactanivorans | 42.7 ± 0.8 |
| Tenacibaculum sp. II2021 | 40.2 ± 5.2 |
| Flavobacterium sp. II2003 | 32.5 ± 1.7 |
| Cellulophaga lytica DSM2039 | 26.3 ± 3.8 |
| Cellulophaga lytica DSM2040 | 19.8 ± 2.1 |
| Bacillus thuringiensis 407 | 17.9 ± 1.8 |
| Bacillus subtilis ND Food | 12.3 ± 0.9 |
| Non Sensitive Strains b | |
| Pseudomonas aeruginosa PA14 | −0.38 ± 0.02 |
| Shewanella sp. IV3014 | −1.9 ± 0.2 |
| Roseobacter sp. IV3009 | −21.1 ± 1.1 |
| Vibrio anguillarum CIP6336T | 5.2 ± 0.7 |
| Vibrio lentus CIP107166T | 5.8 ± 0.4 |
| Vibrio sp. D01 | −12.1 ± 0.2 |
| Producing Bacterium | Source | Target Biofilms | Active Compounds | Action | References |
|---|---|---|---|---|---|
| Pseudoalteromonas ulvae TC14 | Mediterranean Sea, Bay of Toulon, France | Persicivirga (Nonlabens) mediterranea, Shewanella sp., Alteromonas genovensis, Pseudoalteromonas sp. | PS I and/or PS II (exo polysaccharides) | Inhibition of biofilm formation | [64] |
| Pseudoalteromonas haloplanktis TAC125 | Antarctic sea water | Staphylococcus epidermidis | Pentadecanal | - Inhibition of initial attachment - Modulation of the AI-2 quorum sensing system | [20,21,30] |
| Pseudoalteromonas sp. 3J6 | Glass slides immersed in the Morbihan gulf, France | Vibrio sp., Pseudomonas aeruginosa, Escherichia coli, Salmonella enterica, Colwellia sp., Algibacter sp., Micrococcus sp., Paracoccus sp. | 13-kDa protein (Alterocin) | - Inhibition of initial attachment (Vibrio tapetis) - Inhibition of biofilm formation | [18,26,28,29,65] |
| Pseudoalteromonas sp. D41 | Teflon coupon immersed in the Bay of Brest, France | Pseudoalteromonas sp., Paracoccus sp. | Proteinaceous molecule | Inhibition of biofilm formation | [18,27] |
| Pseudoalteromonasruthenica KLPp3 | Marine crab in Pulau Perhentian, Malaysia | Vibrio alginolyticus, Serratia marcescens | Cyclic peptide of the diketopiperazine family | Inhibition of initial attachment and biofilm formation | [31] |
| Pseudoalteromonas sp. 1400 | Sea water of Queensland Beach, Canada | Pseudomonas aeruginosa | 23-kDa alginate lyase (AlyP1400) | Disruption of the established biofilms | [15] |
| Pseudoalteromonas tunicata | Tunicates | Pseudoalteromonas tunicata | 190-kDa autotoxic protein (AlpP) | Killing and detachment of the biofilm from the substratum | [52] |
| Pseudoalteromonas sp. IBRL PD4.8 | Green macroalgae (Caulerpa racemose) Port Dickson, Malaysia | Vibrio alginolyticus | Crude extracts | - Inhibition of the initial and pre-formed biofilms - Antibacterial activity against fouling bacteria | [53] |
| Pseudoalteromonas sp. IIIA004 | Corroded carbon steel coupons immersed in La Rochelle harbor, Atlantic coast, France | Roseovarius sp., Staphylococcus aureus, Yersinia enterocolitica, Paracoccus sp., Pseudomonas aeruginosa PAO1, Micrococcus luteus, Flavobacterium sp., Tenacibaculum sp., Cellulophaga lytica | - Proteinaceous molecule P004 - Culture supernatant | - Inhibition of the biofilm formation without killing the bacteria or inhibiting their growth - Disruption of the Roseovarius sp. mature biofilms | This study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doghri, I.; Portier, E.; Desriac, F.; Zhao, J.M.; Bazire, A.; Dufour, A.; Rochette, V.; Sablé, S.; Lanneluc, I. Anti-Biofilm Activity of a Low Weight Proteinaceous Molecule from the Marine Bacterium Pseudoalteromonas sp. IIIA004 against Marine Bacteria and Human Pathogen Biofilms. Microorganisms 2020, 8, 1295. https://doi.org/10.3390/microorganisms8091295
Doghri I, Portier E, Desriac F, Zhao JM, Bazire A, Dufour A, Rochette V, Sablé S, Lanneluc I. Anti-Biofilm Activity of a Low Weight Proteinaceous Molecule from the Marine Bacterium Pseudoalteromonas sp. IIIA004 against Marine Bacteria and Human Pathogen Biofilms. Microorganisms. 2020; 8(9):1295. https://doi.org/10.3390/microorganisms8091295
Chicago/Turabian StyleDoghri, Ibtissem, Emilie Portier, Florie Desriac, Jean Michel Zhao, Alexis Bazire, Alain Dufour, Vincent Rochette, Sophie Sablé, and Isabelle Lanneluc. 2020. "Anti-Biofilm Activity of a Low Weight Proteinaceous Molecule from the Marine Bacterium Pseudoalteromonas sp. IIIA004 against Marine Bacteria and Human Pathogen Biofilms" Microorganisms 8, no. 9: 1295. https://doi.org/10.3390/microorganisms8091295
APA StyleDoghri, I., Portier, E., Desriac, F., Zhao, J. M., Bazire, A., Dufour, A., Rochette, V., Sablé, S., & Lanneluc, I. (2020). Anti-Biofilm Activity of a Low Weight Proteinaceous Molecule from the Marine Bacterium Pseudoalteromonas sp. IIIA004 against Marine Bacteria and Human Pathogen Biofilms. Microorganisms, 8(9), 1295. https://doi.org/10.3390/microorganisms8091295