
DNA-Metabarcoding of Belowground Fungal Communities in Bare-Root Forest Nurseries: Focus on Different Tree Species

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Sampling
2.2. DNA Isolation, Amplification and Sequencing
2.3. Bioinformatics
2.4. Statistical Analysis
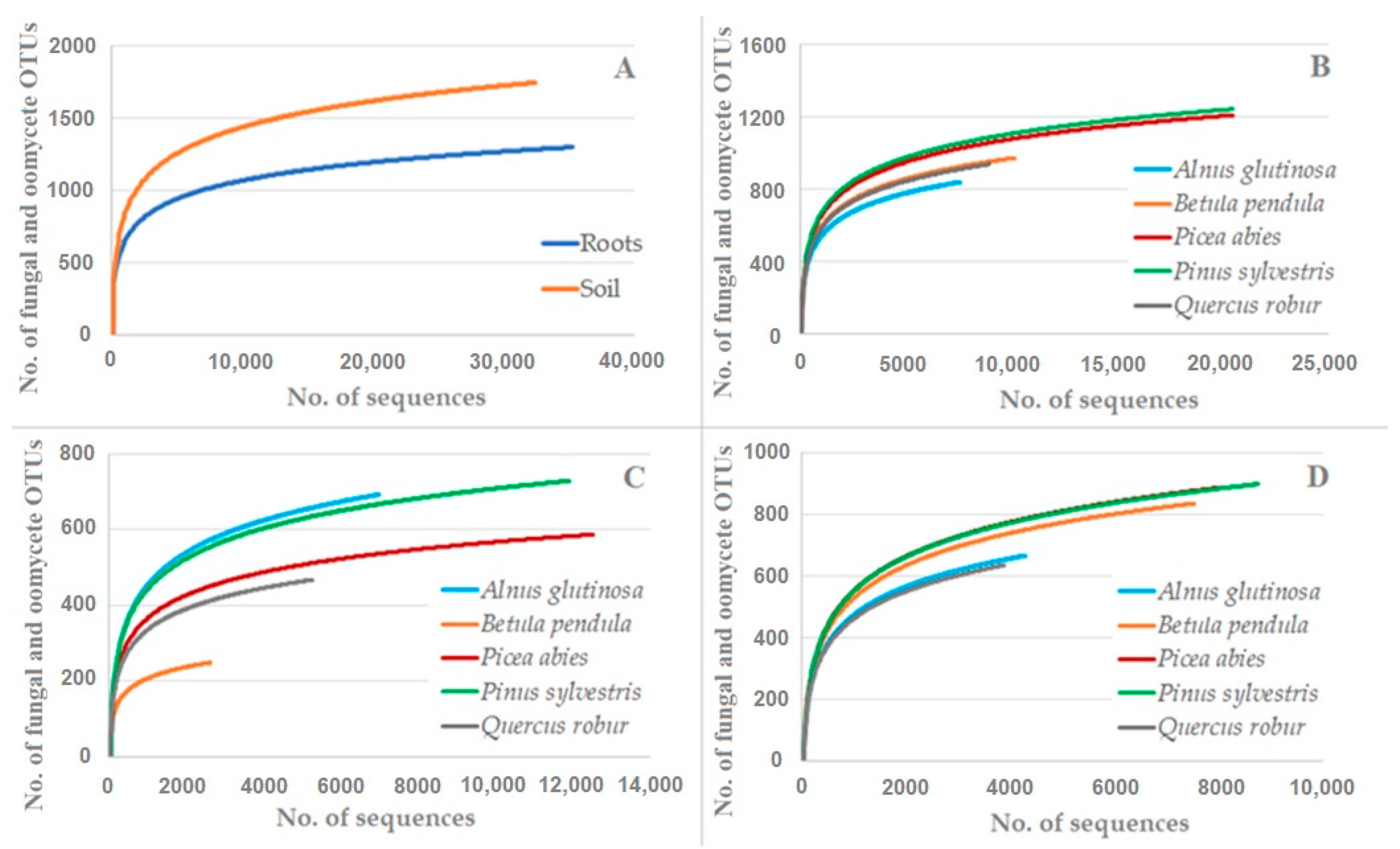
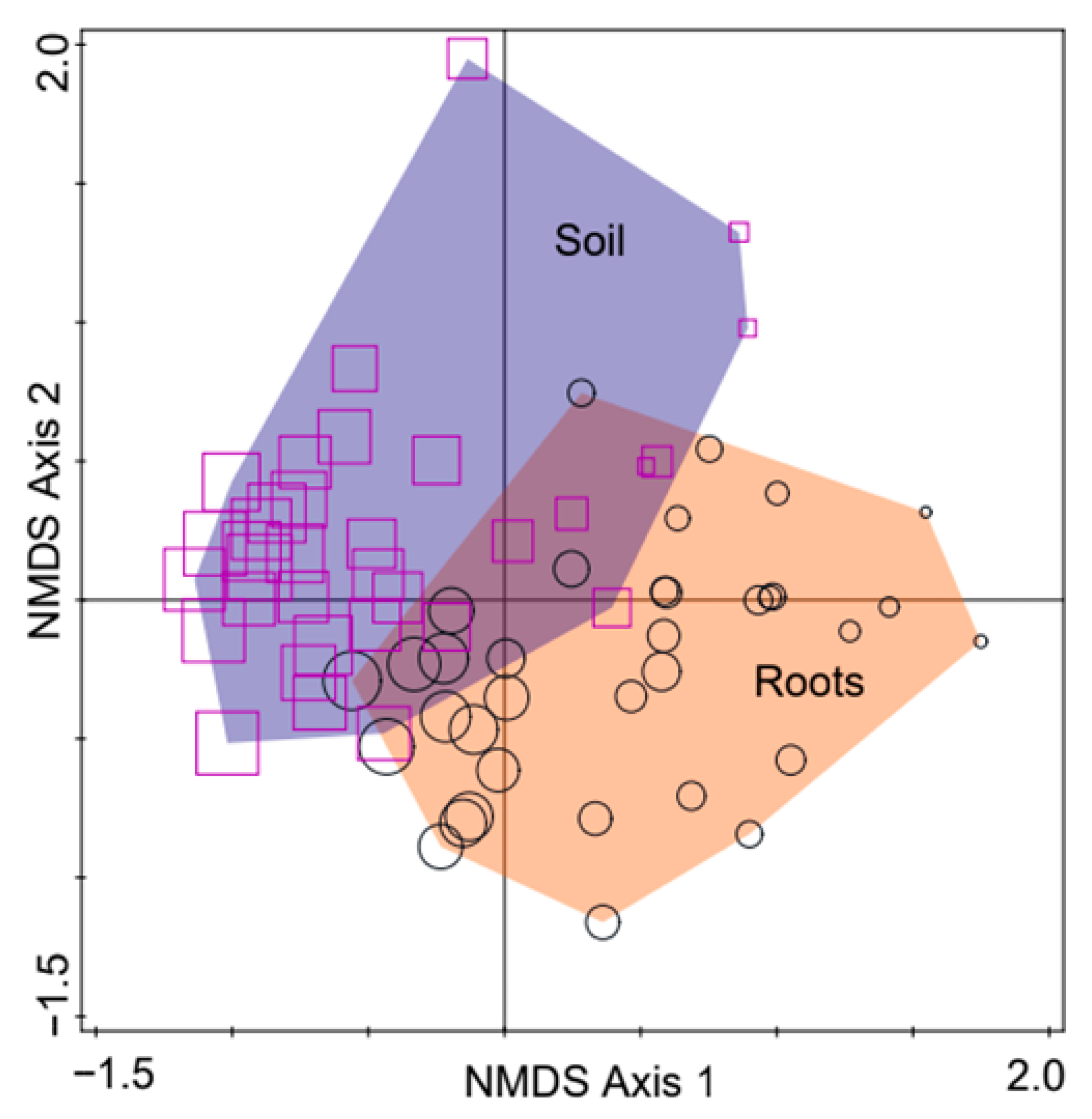
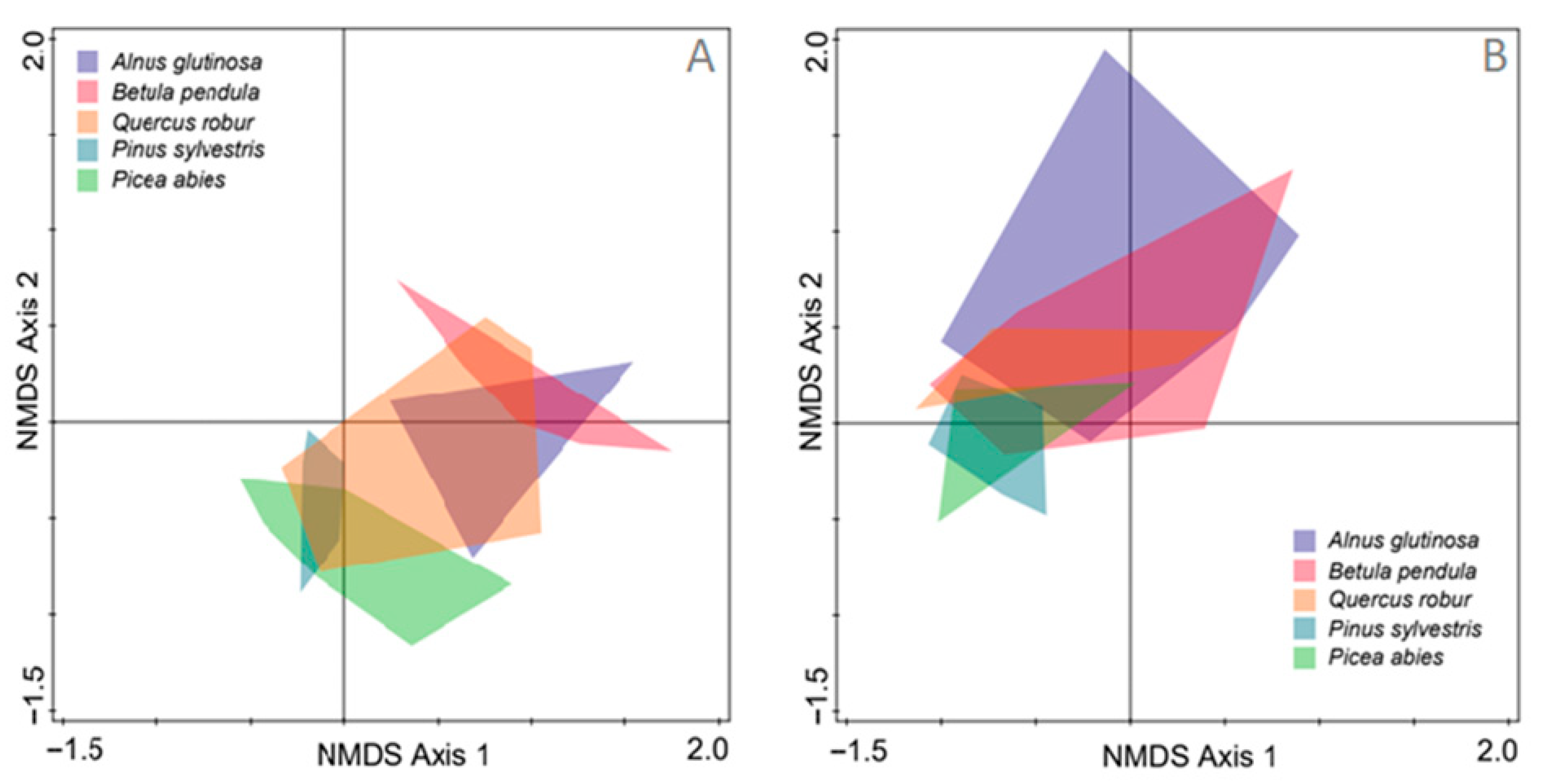
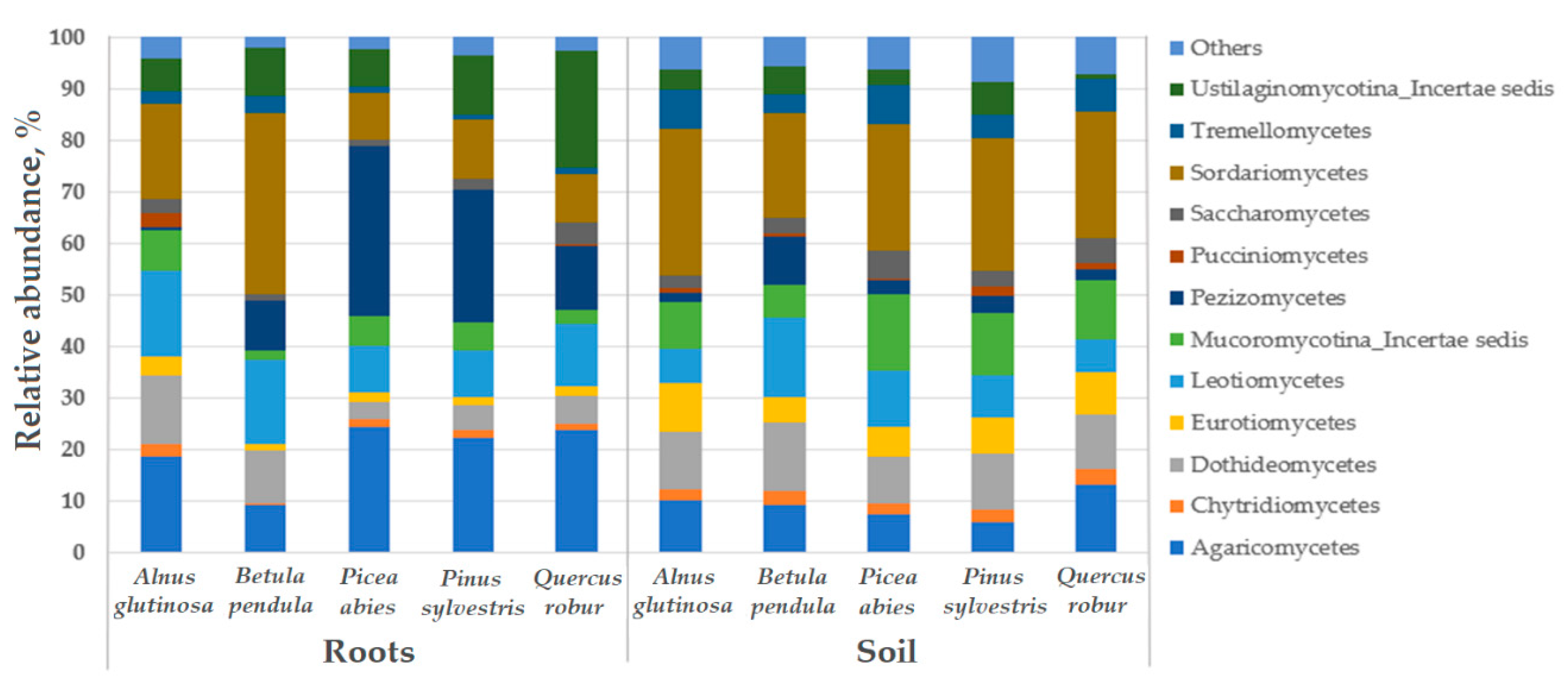
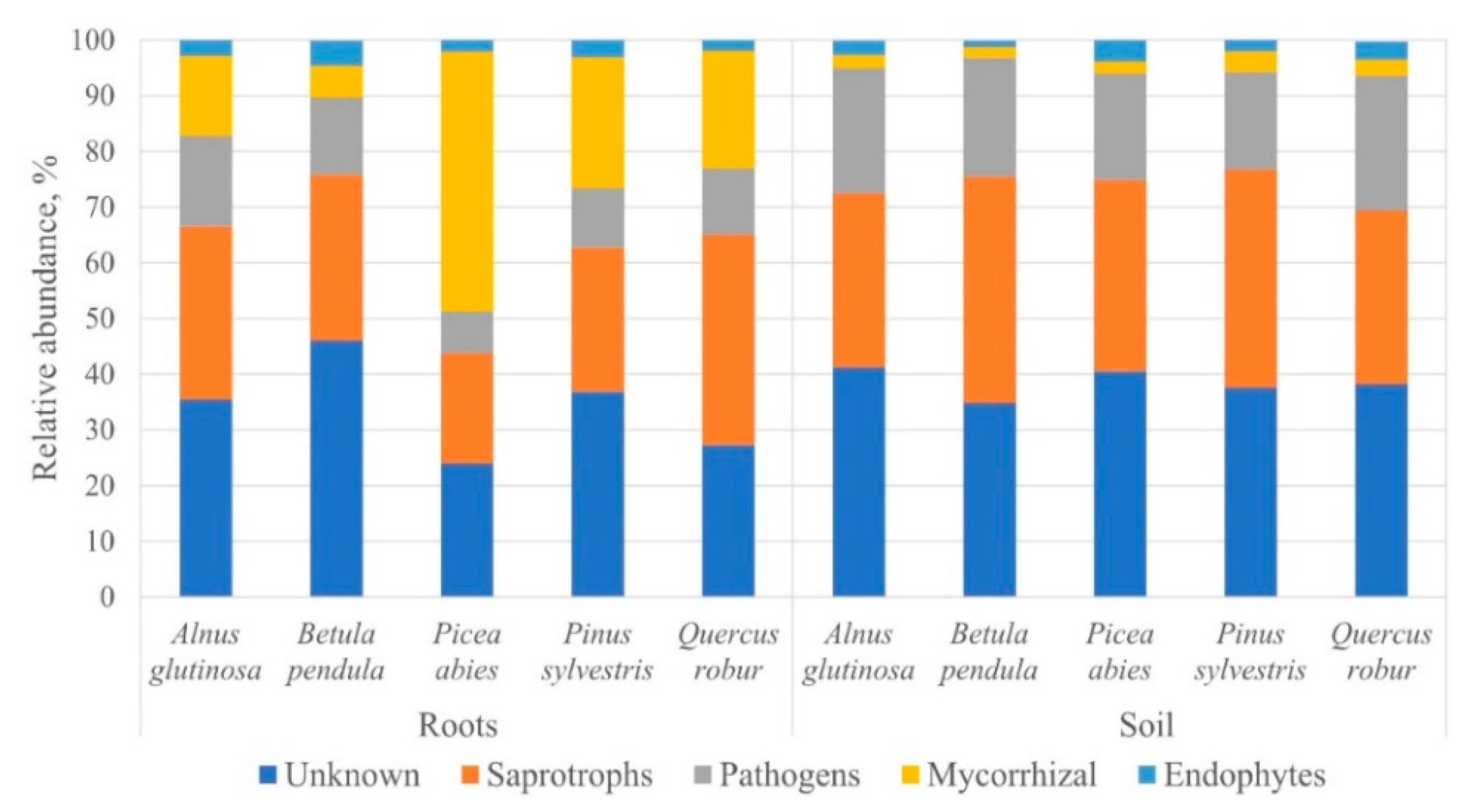
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Karjalainen, T.; Liski, J.; Pussinen, A.; Lapvetela Èinen, T. Sinks in the Kyoto Protocol and Considerations for Ongoing Work in the UNFCCC Work; The Nordic Ministry Council; 2000; in press. [Google Scholar]
- APAT—Auditory Processing Abilities Test. Nursery Production and Stand Establishment of Broad-Leaves to Promote Sustainable Forest Management; APAT—Auditory Processing Abilities Test: Roma, Italy, 2003. [Google Scholar]
- Bastin, J.F.; Finegold, Y.; Garcia, C.; Mollicone, D.; Rezende, M.; Routh, D.; Zohner, C.M.; Crowther, T.W. The global tree restoration potential. Science 2019, 365, 76–79. [Google Scholar] [CrossRef]
- Krasowski, M.J. Forests and Forest Plants—Volume III: Producing Planting Stocks in Forest Nurseries; Eolss Publishers Co.: Oxford, UK, 2012. [Google Scholar]
- Grossnickle, S.C.; MacDonald, J.E. Seedling Quality: History, Application and Plant Attributes. Forests 2018, 9, 283. [Google Scholar] [CrossRef] [Green Version]
- Menkis, A.; Vasiliauskas, R.; Taylor, A.F.S.; Stenlid, J.; Finlay, R. Fungal communities in mycorrhizal roots of conifer seedlings in forest nurseries under different cultivation systems, assessed by morphotyping, direct sequencing and mycelial isolation. Mycorrhiza 2005, 16, 33–41. [Google Scholar] [CrossRef]
- Menkis, A.; Vasiliauskas, R.; Taylor, A.F.S.; Stenström, E.; Stenlid, J.; Finlay, R. Fungi in decayed roots of conifer seedlings in forest nurseries, afforested clear-cuts and abandoned farmland. Plant. Pathol. 2006, 55, 117–129. [Google Scholar] [CrossRef]
- Menkis, A.; Vasiliauskas, R.; Taylor, A.F.S.; Stenlid, J.; Finlay, R. Afforestation of abandoned farmland with conifer seedlings inoculated with three ectomycorrhizal fungi—Impact on plant performance and ectomycorrhizal community. Mycorrhiza 2007, 17, 337–348. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant. Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef]
- Mendes, R.; Kruijt, M.; de Bruijn, I.; Dekkers, E.; van der Voort, M.; Schneider, J.H.M.; Piceno, Y.M.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A.; et al. Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef]
- Ritpitakphong, U.; Falquet, L.; Vimoltust, A.; Berger, A.; Metraux, J.P.; L’Haridon, F. The microbiome of the leaf surface of Arabidopsis protects against a fungal pathogen. New Phytol. 2016, 210, 1033–1043. [Google Scholar] [CrossRef] [Green Version]
- Van der Ent, S.; Van Hulten, M.; Pozo, M.J.; Czechowski, T.; Udvardi, M.K.; Pieterse, C.M.J.; Ton, J. Priming of plant innate immunity by rhizobacteria and β-aminobutyric acid: Differences and similarities in regulation. New Phytol. 2009, 183, 419–431. [Google Scholar] [CrossRef] [Green Version]
- Zamioudis, C.; Korteland, J.; Van Pelt, J.A.; van Hamersveld, M.; Dombrowski, N.; Bai, Y.; Hanson, J.; Van Verk, M.C.; Ling, H.Q.; Schulze-Lefert, P.; et al. Rhizobacterial volatiles and photosynthesis-related signals coordinate MYB72 expression in Arabidopsis roots during onset of induced systemic resistance and iron-deficiency responses. Plant. J. 2015, 84, 309–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Heijden, M.; Bruin, S.; Luckerhoff, L.; van Logtestijn, R.S.; Schlaeppi, K. A widespread plant-fungal-bacterial symbiosis promotes plant biodiversity, plant nutrition and seedling recruitment. ISME J. 2016, 10, 389–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolli, E.; Marasco, R.; Vigani, G.; Ettoumi, B.; Mapelli, F.; Deangelis, M.L.; Gandolfi, C.; Casati, E.; Previtali, F.; Gerbino, R.; et al. Improved plant resistance to drought is promoted by the root-associated microbiome as a water stress-dependent trait. Environ. Microbiol. 2015, 17, 316–331. [Google Scholar] [CrossRef] [PubMed]
- Haney, C.H.; Samuel, B.S.; Bush, J.; Ausubel, F.M. Associations with rhizosphere bacteria can confer an adaptive advantage to plants. Nat. Plants. 2015, 1, 15051. [Google Scholar] [CrossRef] [Green Version]
- Garbaye, J. Helper bacteria—A new dimension to the mycorrhizal symbiosis. New Phytol. 1994, 128, 197–210. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 2nd ed.; Academic Press: London, UK, 1997; p. 605. [Google Scholar]
- Zak, B. The role of mycorrhizae in root disease. Ann. Rev. Phytopathol. 1964, 2, 377–392. [Google Scholar] [CrossRef]
- Vaario, L.M.; Tervonen, A.; Haukioja, K.; Haukioja, M.; Pennanen, T.; Timonen, S. The effect of nursery substrate and fertilization on the growth and ectomycorrhizal status of containerized and out planted seedlings of Picea abies. Can. J. For. Res. 2009, 39, 64–75. [Google Scholar] [CrossRef]
- Kazantseva, O.; Bingham, M.; Simard, S.W.; Berch, S.M. Effects of growth medium, nutrients, water, and aeration on mycorrhization and biomass allocation of greenhouse-grown interior Douglas-fir seedlings. Mycorrhiza 2009, 20, 51–66. [Google Scholar] [CrossRef]
- Henry, C.; Raivoarisoa, J.; Razafimamonjy, A.; Ramanankierana, H.; Andrianaivomahefa, P.; Ducousso, M.; Selosse, M.A. Transfer to forest nurseries significantly affects mycorrhizal community composition of Asteropeia mcphersonii wildings. Mycorrhiza 2017, 27, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Raj, A.J.; Lal, S.B. Forestry Principles and Applications; Scientific Publishers: Rajasthan, India, 2013. [Google Scholar]
- Jung, T.; Orlikowski, L.; Henricot, B.; Abad-Campos, P.; Aday, A.G.; Aguín Casal, O.; Bakonyi, J.; Cacciola, S.O.; Cech, T.; Chavarriaga, D.; et al. Widespread Phytophthora infestations in European nurseries put forest, semi-natural and horticultural ecosystems at high risk of Phytophthora diseases. For. Path. 2016, 46, 134–163. [Google Scholar] [CrossRef] [Green Version]
- Peterson, G.W.; Smith, R.S., Jr. Forest Nursery Diseases in the United States; Agriculture Hand-Book No 470. Forest Service 1975. Library of Congress Catalog No. 74-600103; U.S. Department of Agriculture: Washington, DC, USA, 1975; p. 125.
- Brasier, C.M. The biosecurity threat to the UK and global environment from international trade in plants. Plant. Pathol. 2008, 57, 792–808. [Google Scholar] [CrossRef]
- Liebhold, A.M.; Brockerhoff, E.G.; Garrett, L.J.; Parke, J.L.; Britton, K.O. Live plant imports: The major pathway for forest insect and pathogen invasions of the US. Front. Ecol. Environ. 2012, 10, 135–143. [Google Scholar] [CrossRef]
- Yakabe, L.E.; Blomquist, C.L.; Thomas, S.L.; MacDonald, J.D. Identification and frequency of Phytophthora species associated with foliar diseases in California ornamental nurseries. Plant. Dis. 2009, 93, 883–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bienapfl, J.C.; Balci, Y. Movement of Phytophthora spp. in Maryland’s nursery trade. Plant. Dis. 2014, 98, 134–144. [Google Scholar] [CrossRef] [Green Version]
- Parke, J.L.; Knaus, B.J.; Fieland, V.J.; Lewis, C.; Grünwald, N.J. Phytophthora community structure analyses in Oregon nurseries inform systems approaches to disease management. Phytopathology 2014, 104, 1052–1062. [Google Scholar] [CrossRef] [Green Version]
- Reeser, P.W.; Sutton, W.; Hansen, E.M.; Goheen, E.M.; Fieland, V.J.; Grunwald, N.J. First report of Phytophthora occultans causing root and collar rot on Ceanothus, boxwood, rhododendron, and other hosts in horticultural nurseries in Oregon, USA. Plant. Dis. 2015, 99, 1282. [Google Scholar] [CrossRef]
- Rooney-Latham, S.; Blomquist, C.; Swiecki, T.; Bernhardt, E.; Frankel, S.J. First detection in the USA: New plant pathogen, Phytophthora tentaculata, in native plant nurseries and restoration sites in California. Native Plants J. 2015, 16, 23–25. [Google Scholar] [CrossRef]
- Rooney-Latham, S.; Blomquist, C.L.; Kosta, K.L.; Gou, Y.Y.; Woods, P.W. Phytophthora species are common on nursery stock grown for restoration and revegetation purposes in California. Plant. Dis. 2019, 103, 448–455. [Google Scholar] [CrossRef] [Green Version]
- Dobbelaere, S.; Vanderleyden, J.; Okon, Y. Plant growth-promoting effects of diazotrophs in the rhizosphere. Crit. Rev. Plant. Sci. 2003, 22, 107–149. [Google Scholar] [CrossRef]
- Duffy, B.; Keel, C.; Defago, G. Potential role of pathogen signaling in multitrophic plant-microbe interactions involved in disease protection. Appl. Environ. Microbiol. 2004, 70, 1836–1842. [Google Scholar] [CrossRef] [Green Version]
- Morgan, J.A.W.; Bending, G.D.; White, P.J. Biological costs and benefits to plant-microbe interactions in the rhizosphere. J. Exp. Bot. 2005, 56, 1729–1739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhart, K.O.; Callaway, R.M. Soil biota and invasive plants. New Phytol. 2006, 170, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Batten, K.M.; Scow, K.M.; Davies, K.F.; Harrison, S.P. Two invasive plants alter soil microbial community composition in serpentine grasslands. Biol. Invasions 2006, 8, 217–230. [Google Scholar] [CrossRef]
- Innes, L.; Hobbs, P.J.; Bardgett, R.D. The impacts of individual plant species on rhizosphere microbial communities in soils of different fertility. Biol. Fertil. Soils 2004, 40, 7–13. [Google Scholar] [CrossRef]
- Priha, O.; Grayston, S.J.; Pennanen, T.; Smolander, A. Microbial activities related to C and N cycling and microbial community structure in the rhizospheres of Pinus sylvestris, Picea abies and Betula pendula seedlings in an organic and mineral soil. FEMS Microbiol. Ecol. 1999, 30, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Kowalchuk, G.A.; Hol, W.H.G.; Van Veen, J.A. Rhizosphere fungal communities are influenced by Senecio jacobaea pyrrolizidine alkaloid content and composition. Soil Biol. Biochem. 2006, 38, 2852–2859. [Google Scholar] [CrossRef]
- Klavina, D.; Zaluma, A.; Pennanen, T.; Velmala, S.; Gaitnieks, T.; Gailis, A.; Menkis, A. Seed provenance impacts growth and ectomycorrhizal colonisation of Picea abies seedlings. Balt. For. 2015, 21, 184–191. [Google Scholar]
- Mougel, C.; Offre, P.; Ranjard, L.; Corberand, T.; Gamalero, E.; Robin, C.; Lemanceau, P. Dynamic of the genetic structure of bacterial and fungal communities at different developmental stages of Medicago truncatula Gaertn. cv. Jemalong line J5. New Phytol. 2006, 170, 165–175. [Google Scholar] [CrossRef]
- Weisskopf, L.; Tomasi, N.; Santelia, D.; Martinoia, E.; Langlade, N.B.; Tabacchi, R.; Abou-Mansour, E. Isoflavonoid exudation from white lupin roots is influenced by phosphate supply, root type and cluster-root stage. New Phytol. 2006, 171, 657–668. [Google Scholar] [CrossRef] [Green Version]
- Stenström, E.; Ndobe, N.E.; Jonsson, M.; Stenlid, J.; Menkis, A. Root associated fungi of healthy-looking Pinus sylvestris and Picea abies seedlings in Swedish forest nurseries. Scand. J. For. Res. 2014, 29, 12–21. [Google Scholar] [CrossRef]
- Bzdyk, R.M.; Olchowik, J.; Studnicki, M.; Oszako, T.; Sikora, K.; Szmidla, H.; Hilszczańska, D. The impact of effective microorganisms (em) and organic and mineral fertilizers on the growth and mycorrhizal colonization of Fagus sylvatica and Quercus robur seedlings in a bare-root nursery experiment. Forests 2018, 9, 597. [Google Scholar] [CrossRef] [Green Version]
- Beyer-Ericson, L.; Damm, E.; Unestam, T. An overview of root dieback and its causes in Swedish forest nurseries. Eur. J. For. Pathol. 1991, 21, 439. [Google Scholar] [CrossRef]
- Galaaen, R.; Venn, K. Pythium sylvaticum Campbell & Hendrix and other fungi associated with root dieback of 2-0 seedlings of Picea abies (L.) Karst. in Norway. Meddelerser Norsk Inst. Skogforsk. 1979, 34, 221–228. [Google Scholar]
- Venn, K. Rotavdoing hos bartreplanter i skogplanteskoler (Root dieback of coniferous seedlings in forest nurseries). In Norwegian, English Summary; Research Paper 3/85; Norwegian Forest Research Institute: Ås, Norway, 1985; Volume 11. [Google Scholar]
- Venn, K.; Sandvik, M.; Langerud, B. Nursery routines, growth media and pathogens affect growth and root dieback in Norway spruce seedlings. Meddelelser Norsk Inst. Skogforsk. 1986, 34, 314–328. [Google Scholar]
- Lilja, A.; Lilja, S.; Poteri, M.; Ziren, L. Conifer seedling root fungi and root dieback in Finnish nurseries. Scand. J. For. Res. 1992, 7, 547–556. [Google Scholar] [CrossRef]
- Lilja, A. The occurrence and pathogenicity of uni- and binucleate Rhizoctonia and Pythiaceae fungi among conifer seedlings in Finnish forest nurseries. Eur. J. Plant. Pathol. 1994, 24, 181–192. [Google Scholar] [CrossRef]
- Santini, A.; Ghelardini, L.; De Pace, C.; Desprez-Loustau, M.L.; Capretti, P.; Chandelier, A.; Cech, T.; Chira, D.; Diamandis, S.; Gaitniekis, T.; et al. Biogeographical patterns and determinants of invasion by forest pathogens in Europe. New Phytol. 2013, 197, 238–250. [Google Scholar] [CrossRef] [Green Version]
- Cram, M.M.; Hansen, E.M. Phytophthora Root Rot. In Forest Nursery Pests; Agriculture Handbook 680 Rev.; Cram, M.M., Frank, M.S., Mallams, K.M., Eds.; Department of Agriculture, Forest Service: Washington, DC, USA, 2012; pp. 126–128. [Google Scholar]
- Goheen, E.M.; Kanaskie, A.; Navarro, S.; Hansen, E. Sudden oak death management in Oregon tanoak forests. For. Phytophthoras 2017, 7, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Stenlid, J.; Oliva, J.; Boberg, J.B.; Hopkins, A.J.M. Emerging diseases in European forest ecosystems and responses in society. Forests 2011, 2, 486–504. [Google Scholar] [CrossRef]
- Vaičys, M. Miško dirvožemių klasifikacija. In Lietuvos Dirvožemiai [Forest Site Types. Lithuanian Soils]; Mokslas Publishers: Vilnius, Lithuania, 2001; pp. 1040–1043. (In Lithuanian) [Google Scholar]
- Marčiulynas, A.; Marčiulynienė, D.; Lynikienė, J.; Gedminas, A.; Vaičiukynė, M.; Menkis, A. Fungi and oomycetes in the irrigation water of forest nurseries. Forests 2020, 11, 459. [Google Scholar] [CrossRef] [Green Version]
- Cooke, D.E.L.; Drenth, A.; Duncan, J.M.; Wagels, G.; Brasier, C.M. A molecular phylogeny of Phytophthora and related oomycetes. Fungal Genet. Biol. 2000, 30, 17–32. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988; p. 192. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 3rd ed.; W.H. Freeman and Company: New York, NY, USA, 1995. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.; Boberg, J.; Ihrmark, K.; Stenstrom, E.; Stenlid, J. Do foliar fungal communities of Norway spruce shift along a tree species diversity gradient in mature European forests? Fungal Ecol. 2016, 23, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Menkis, A.; Burokienė, D.; Stenlid, J.; Stenström, E. High-throughput sequencing shows high fungal diversity and community segregation in the rhizospheres of container-grown conifer seedlings. Forests 2016, 7, 44. [Google Scholar] [CrossRef] [Green Version]
- Mickan, B.S.; Hart, M.M.; Solaiman, Z.M.; Jenkins, S.; Siddique, K.H.M.; Abbott, L.K. Molecular divergence of fungal communities in soil, roots and hyphae highlight the importance of sampling strategies. Rhizosphere 2017, 4, 104–111. [Google Scholar] [CrossRef]
- Pascale, A.; Proietti, S.; Pantelides, I.S.; Stringlis, I.A. Modulation of the root microbiome by plant molecules: The basis for targeted disease suppression and plant growth promotion. Front. Plant. Sci. 2020, 10, 1741. [Google Scholar] [CrossRef]
- Aponte, C.; García, L.V.; Marañón, T. Tree species effects on nutrient cycling and soil biota: A feedback mechanism favouring species coexistence. For. Ecol. Manage. 2013, 309, 36–46. [Google Scholar] [CrossRef] [Green Version]
- Klaubauf, S.; Inselsbacher, E.; Zechmeister-Boltenstern, S.; Wanek, W.; Gottsberger, R.; Strauss, J.; Gorfer, M. Molecular diversity of fungal communities in agricultural soils from Lower Austria. Fungal Divers. 2010, 44, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.M.; Chen, C.R.; Xu, Z.H.; Liu, Y.Q. Effects of single and mixed species forest ecosystems on diversity and function of soil microbial community in subtropical China. J. Soils Sediments 2012, 12, 228–240. [Google Scholar] [CrossRef]
- Han, L.L.; Wang, J.T.; Yang, S.H.; Chen, W.F.; Zhang, L.M.; He, J.Z. Temporal dynamics of fungal communities in soybean rhizosphere. J. Soils Sediments 2016, 17, 491–498. [Google Scholar] [CrossRef]
- Petrini, O. Fungal Endophytes of Tree Leaves. In Microbial Ecology of Leaves; Andrews, J.H., Hirano, S.S., Eds.; Springer: New York, NY, USA, 1991; pp. 179–197. [Google Scholar]
- King, B.C.; Waxman, K.D.; Nenni, N.V.; Walker, L.P.; Bergstrom, G.C.; Gibson, D.M. Arsenal of plant cell wall degrading enzymes reflects host preference among plant pathogenic fungi. Biotechnol. Biofuels 2011, 4, 1–14. [Google Scholar]
- Lang, C.; Seven, J.; Polle, A. Host preferences and differential contributions of deciduous tree species shape mycorrhizal species richness in a mixed Central European forest. Mycorrhiza 2011, 21, 297–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayerhofer, M.S.; Kernaghan, G.; Harper, K.A. The effects of fungal root endophytes on plant growth: A meta-analysis. Mycorrhiza 2013, 23, 119–128. [Google Scholar] [CrossRef]
- Austin, A.T.; Vivanco, L.; Gonzalez-Arzac, A.; Perez, L.I. There’s no place like home? An exploration of the mechanisms behind plant litter-decomposer affinity in terrestrial ecosystems. New Phytol. 2014, 204, 307–314. [Google Scholar] [CrossRef]
- Jumpponen, A.; Egerton-Warburton, L.M. Mycorrhizal fungi in successional environments—A community assembly model incorporating host plant, environmental and biotic filters. In The Fungal Community; Dighton, J., White, J.F., Oudemans, P., Eds.; CRC Press: New York, NY, USA, 2005; pp. 139–180. [Google Scholar]
- Ishida, T.A.; Nara, K.; Hogetsu, T. Host effects on ectomycorrhizal fungal communities: Insight from eight host species in mixed conifer–broadleaf forests. New Phytol. 2007, 174, 430–440. [Google Scholar] [CrossRef]
- Menkis, A.; Vasaitis, R. Fungi in roots of nursery grown Pinus sylvestris: Ectomycorrhizal colonisation, genetic diversity and spatial distribution. Microb Ecol 2011, 61, 52–63. [Google Scholar] [CrossRef]
- Vrålstad, T.; Myhre, E.; Schumacher, T. Molecular diversity and phylogenetic affinities of symbiotic root-associated ascomycetes of the Helotiales in burnt and metal polluted habitats. New Phytol. 2002, 155, 131–148. [Google Scholar] [CrossRef]
- Sugita, T.; Boekhout, T.; Velegraki, A.; Guillot, J.; Hađina, S.; Cabañes, F.J. Epidemiology of Malassezia-related skin diseases. In Malassezia and the Skin: Science and Clinical Practice, 1st ed.; Boekhout, T., Guého-Kellermann, E., Mayser, P., Velegraki, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 65–120. [Google Scholar]
- Lai, X.; Cao, L.; Tan, H.; Fang, S.; Huang, Y.; Zhou, S. Fungal communities from methane hydrate-bearing deep-sea marine sediments in South China Sea. ISME J. 2007, 1, 756–762. [Google Scholar] [CrossRef]
- Le Calvez, T.; Burgaud, G.; Mahe, S.; Barbier, G.; Vandenkoornhuyse, P. Fungal diversity in deep-sea hydrothermal ecosystems. Appl. Environ. Microbiol. 2009, 75, 6415–6421. [Google Scholar] [CrossRef] [Green Version]
- Amend, A.S.; Barshis, D.J.; Oliver, T.A. Coral-associated marine fungi form novel lineages and heterogeneous assemblages. ISME J. 2012, 6, 1291–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arenz, B.E.; Held, B.W.; Jurgens, J.A.; Farrell, R.L.; Blanchette, R.A. Fungal diversity in soils and historic wood from the Ross Sea Region of Antarctica. Soil Biol. Biochem. 2006, 38, 3057–3064. [Google Scholar] [CrossRef]
- Fell, J.W.; Scorzetti, G.; Connell, L.; Craig, S. Biodiversity of micro-eukaryotes in Antarctic Dry Valley soils with. Soil Biol. Biochem. 2006, 38, 3107–3119. [Google Scholar] [CrossRef]
- Renker, C.; Alphei, J.; Buscot, F. Soil nematodes associated with the mammal pathogenic fungal genus Malassezia (Basidiomycota: Ustilaginomycetes) in Central European forests. Biol. Fertil Soils 2003, 37, 70–72. [Google Scholar] [CrossRef]
- Roy, M.; Watthana, S.; Stier, A.; Richard, F.; Vessabutr, S.; Selosse, M.A. Two mycoheterotrophic orchids from Thailand tropical dipterocarpacean forests associate with a broad diversity of ectomycorrhizal fungi. BMC Biol. 2009, 7, 51. [Google Scholar] [CrossRef] [Green Version]
- Klavina, D.; Menkis, A.; Gaitnieks, T.; Pennanen, T.; Lazdiņš, A.; Velmala, S.; Vasaitis, R. Low impact of stump removal on mycorrhization and growth of replanted Picea abies: Data from three types of hemiboreal forest. Balt For. 2016, 22, 16–24. [Google Scholar]
- Renseigné, N.; Rudawska, M.; Leski, T. Mycorrhizal associations of nursery grown Scots pine (Pinus sylvestris L.) seedlings in Poland. Ann. For. Sci. 2006, 63, 715–723. [Google Scholar]
- Zwiazek, J.J.; Equiza, M.A.; Karst, J.; Senorans, J.; Wartenbe, M.; Calvo-Polanco, M. Role of urban ectomycorrhizal fungi in improving the tolerance of lodgepole pine (Pinus contorta) seedlings to salt stress. Mycorrhiza 2019, 29, 303–312. [Google Scholar] [CrossRef]
- Lazarevic, J.; Menkis, A. Fungi inhabiting fine roots of Pinus heldreichii in the Montenegrin montane forests. Symbiosis 2018, 74, 189–197. [Google Scholar] [CrossRef]
- Smaill, S.J.; Walbert, K. Fertilizer and fungicide use increases the abundance of less beneficial ectomycorrhizal species in a seedling nursery. Appl. Soil Ecol. 2013, 65, 60–64. [Google Scholar] [CrossRef]
- Jones, M.D.; Grenon, F.; Peat, H.; Fitzgerald, M.; Holt, L.; Philip, L.J.; Bradley, R. Differences in N-15 uptake amongst spruce seedlings colonized by three pioneer ectomycorrhizal fungi in the field. Fungal Ecol. 2009, 2, 110–120. [Google Scholar] [CrossRef]
- Chu-Chou, M.; Grace, L.J. Mycorrhizal fungi of radiata pine in different forests of the North and South Islands in New Zealand. Soil Biol. Biochem. 1988, 20, 883–886. [Google Scholar] [CrossRef]
- Menkis, A.; Lygis, V.; Burokienė, D.; Vasaitis, R. Establishment of ectomycorrhiza-inoculated Pinus sylvestris seedlings on coastal dunes following a forest fire. Balt For. 2012, 18, 33–40. [Google Scholar]
- James, R.L.; Dumroese, R.K. Investigations of Fusarium diseases within Inland Pacific Northwest forest nurseries. In Proceedings of the 53rd Western International Forest Disease Work Conference, Jackson, WY, USA, 26–29 August 2005; Guyon, J.C., Ed.; USDA Forest Service, Intermountain Region: Ogden, UT, USA, 2007; pp. 3–11. [Google Scholar]
- Adesemoye, A.; Eskalen, A.; Faber, B.; O’Connell, N. Current nowledge on Fusarium dry root rot of citrus. Citrograph 2011, 2, 29–33. [Google Scholar]
- Adesemoye, A.O.; Mayorquin, J.S.; Peacock, B.B.; Moreno, K.; Hajeri, S.; Yokomi, R.; Eskalen, A. Association of Neonectria macrodidyma with dry root rot of citrus in California. J. Plant. Pathol. Microbiol. 2016, 8, 391. [Google Scholar]
- Hamelin, R.C.; Berube, P.; Gignac, M.; Bourassa, M. Identification of root rot fungi in nursery seedlings by nested multiplex PCR. Appl. Environ. Microbiol. 1996, 62, 4026–4031. [Google Scholar] [CrossRef] [Green Version]
- Menkis, A.; Burokienė, D. Distribution and genetic diversity of the root-rot pathogen Neonectria macrodidyma in a forest nursery. For. Pathol. 2012, 42, 79–83. [Google Scholar] [CrossRef]
- Derevnina, L.; Petre, B.; Kellner, R.; Dagdas, Y.F.; Sarowar, M.N.; Giannakopoulou, A.; De la Concepcion, J.C.; Chaparro-Garcia, A.; Pennington, H.G.; van West, P.; et al. Emerging oomycete threats to plants and animals. Philosophical transactions of the Royal Society of London. Proc. R. Soc. B 2016, 371, 20150459. [Google Scholar]
- Lazerg, F.; Belabid, L.; Sanchez, J.; Gallego, E. Root rot and damping-off of Aleppo pine seedlings caused by Pythium spp. in Algerian forest nurseries. For. Sci. 2016, 62, 322–328. [Google Scholar] [CrossRef] [Green Version]
- Paulitz, T.C.; Adams, K. Composition and distribution of Pythium communities in wheat fields in eastern washington state. Ecol. Popul. Biol. 2003, 93, 867. [Google Scholar] [CrossRef] [Green Version]
- Jiménez, J.J.; Sánchez, J.E.; Romero, M.A.; Belbahri, L.; Trapero, A.; Lefort, F.; Sánchez, M.E. Pathogenicity of Pythium spiculum and P. sterilum on feeder roots of Quercus rotundifolia. Plant. Pathol. 2008, 57, 369. [Google Scholar] [CrossRef]
- Serrano, M.D.; de Vita, P.; Fernández-Rebollo, P.; Coelho, A.C.; Belbahri, L.; Sánchez, M.E. Phytophthora cinnamomi and Pythium spiculum as main agents of Quercus decline in southern Spain and Portugal. IOBC/WPRS Bull. 2012, 76, 97–100. [Google Scholar]
- de Vita, P.; Serrano, M.S.; Belbahri, L.; García, L.V.; Ramo, C.; Sánchez, M.E. Germination of hyphal bodies of Pythium spiculum isolated from declining cork oaks at Doñana National Park (Spain). Phytopathol. Mediterr. 2011, 50, 478–481. [Google Scholar]
- Paul, B.; Bala, K.; Belbahri, L.; Calmin, G.; Sánchez, E.; Lefort, F. A new species of Pythium with ornamented oogonia: Morphology, taxonomy, ITS region of its rDNA, and its comparison with related species. FEMS Microbiol. Lett. 2006, 254, 317–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, C.; Kageyama, K.; Suga, H.; Hyakumachi, M. Intraspecific DNA polymorphisms of Pythium irregulare. Mycol. Res. 2000, 104, 1333–1341. [Google Scholar] [CrossRef]
- Thines, M.; Choi, Y.J. Evolution, diversity, and taxonomy of the Peronosporaceae, with focus on the genus Peronospora. Phytopathology 2016, 106, 6–18. [Google Scholar] [CrossRef] [Green Version]
- Scanu, B.; Jones, B.; Webber, J.F. A new disease of Nothofagus in Britain caused by Phytophthora pseudosyringae. New Dis. Rep. 2012, 25, 27. [Google Scholar] [CrossRef] [Green Version]
- Scheewe, P. Identification of pathogenic races of Phytophthora fragariae Hickman in Germany. In Progress in Temperate Fruit Breeding, Developments in Plant Breeding; Schmidt, H., Kellerhals, M., Eds.; Springer: Dordrecht, Germany, 1994; Volume 1, pp. 67–71. [Google Scholar]
- Jung, T.; Nechwatal, J.; Cooke, D.E.; Hartmann, G.; Blaschke, M.; Oßwald, W.F.; Duncan, J.M.; Delatour, C. Phytophthora pseudosyringae sp. nov., a new species causing root and collar rot of deciduous tree species in Europe. Mycol Res. 2003, 107, 772–789. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Nursery | Area of the Nursery, ha | Total No. of Seedlings Produced, Millions | No. of Seedlings Produced by Tree Species, Millions | Mean Monthly Temperature, °C * | Mean Monthly Precipitation, mm * | Soil Type | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Alnus glutinosa | Betula pendula | Picea abies | Pinus sylvestris | Quercus robur | ||||||
| Alytus | 23.1 | 4.7 | 0.2 | 0.4 | 2.6 | 1.1 | 0.3 | 10.5 | 49.6 | Nc |
| Anykščiai | 34.0 | 5.6 | 0.3 | 0.3 | 4.6 | 0.2 | 0.2 | 10.1 | 38.2 | Nc |
| Dubrava | 56.3 | 7.1 | 0.3 | 0.2 | 5.2 | 1.2 | 0.2 | 10.2 | 64.8 | Nc |
| Kaišiado-rys | 27.0 | 4.6 | 0.2 | 0.2 | 3.6 | 0.2 | 0.3 | 9.9 | 52.1 | Nc |
| Kretinga | 66.0 | 5.5 | 0.1 | 0.1 | 4.4 | 0.4 | 0.2 | 8.8 | 61.6 | Nc |
| Panevėžys | 66.0 | 8.8 | 0.4 | 0.3 | 7.4 | 0.3 | 0.5 | 9.8 | 46.0 | Lc |
| Trakai | 31.4 | 5.1 | 0.1 | 0.2 | 3.2 | 1.3 | 0.1 | 10.3 | 41.0 | Nb |
| Samples | Tree Species | No. of Fungal Sequences/Taxa | Shannon Diversity Index H a |
|---|---|---|---|
| Roots | Alnus glutinosa | 6979/691 | 2.16–4.33 |
| Betula pendula | 2620/256 | 1.93–4.09 | |
| Picea abies | 12,532/591 | 1.75–4.28 | |
| Pinus sylvestris | 11,910/718 | 2.62–4.32 | |
| Quercus robur | 5270/453 | 2.14–4.23 | |
| Total Roots | 39,311/1414 | ||
| Soil | Alnus glutinosa | 4292/664 | 3.28–5.03 |
| Betula pendula | 7525/836 | 2.57–5.20 | |
| Picea abies | 7961/873 | 3.28–4.89 | |
| Pinus sylvestris | 8745/881 | 3.56–4.69 | |
| Quercus robur | 3862/646 | 3.83–5.17 | |
| Total Soil | 32,385/1737 | ||
| All | 71,696/2003 |
| OTU | Phylum | Reference | Similarity,% a | Alnus glutinosa | Betula pendula | Picea abies | Pinus sylvestris | Quercus robur | All | All | Total% | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Roots% | Soil% | Roots% | Soil% | Roots% | Soil% | Roots% | Soil% | Roots% | Soil% | Roots% | Soil% | |||||
| Malassezia restricta | Basidiomycota | CP030254 | 727/728 (99) | 5.79 | 3.59 | 8.02 | 4.96 | 6.57 | 2.85 | 10.43 | 6.00 | 20.75 | 0.85 | 9.01 | 4.05 | 6.70 |
| Wilcoxina mikolae | Ascomycota | KU061020 | 592/593 (99) | 0.11 | 0.09 | 0.38 | 0.07 | 22.28 | 0.95 | 3.82 | 1.46 | 0.88 | 0.08 | 8.82 | 0.67 | 5.03 |
| Pustularia sp. 3993_4 | Ascomycota | MF352743 | 513/518 (99) | - | 0.26 | 0.15 | 4.52 | 7.50 | 0.04 | 15.53 | 0.03 | 0.82 | - | 7.56 | 1.10 | 4.56 |
| Fusarium oxysporum | Ascomycota | GU136492 | 547/547 (100) | 2.41 | 5.24 | 0.73 | 4.44 | 3.28 | 4.81 | 2.99 | 4.63 | 1.91 | 2.54 | 2.72 | 4.46 | 3.53 |
| Tomentella sp. 3993_7 | Basidiomycota | KX095160 | 648/663 (98) | 0.01 | 0.05 | - | 0.07 | 14.20 | 0.15 | 0.07 | 0.01 | 0.15 | 0.00 | 4.80 | 0.06 | 2.60 |
| Suillus luteus | Basidiomycota | KU721223 | 688/688 (100) | - | 0.02 | 0.04 | 0.00 | 0.06 | 0.01 | 6.90 | 0.77 | 0.12 | 0.08 | 2.23 | 0.22 | 1.30 |
| Mycoarthris corallina | Ascomycota | AH009124 | 467/473 (99) | 6.53 | 0.44 | 0.57 | 9.13 | 0.02 | 0.82 | 0.06 | 0.11 | 0.09 | 0.16 | 1.30 | 2.43 | 1.82 |
| Paraphaeosphaeria sporulosa | Ascomycota | KY977581 | 594/594 (100) | 1.83 | 1.49 | 0.53 | 1.62 | 1.29 | 2.42 | 1.22 | 3.90 | 0.52 | 1.11 | 1.25 | 2.36 | 1.76 |
| Chaetomium cochliodes | Ascomycota | KT895345 | 570/570 (100) | 0.42 | 2.10 | 0.27 | 1.73 | 1.51 | 3.13 | 1.20 | 3.56 | 0.40 | 1.32 | 1.02 | 2.57 | 1.74 |
| Pleotrichocladium opacum | Ascomycota | NR155696 | 545/549 (99) | 0.44 | 0.40 | 28.66 | 0.54 | 0.80 | 0.78 | 0.42 | 0.29 | 0.79 | 0.54 | 2.57 | 0.51 | 1.61 |
| Halenospora varia | Ascomycota | AJ608987 | 538/538 (100) | 1.09 | 0.91 | 4.77 | 0.43 | 1.39 | 1.88 | 1.39 | 1.44 | 3.71 | 1.58 | 1.78 | 1.26 | 1.54 |
| Tomentella sublilacina | Basidiomycota | HM189981 | 648/662 (98) | 12.94 | 1.28 | 0.04 | 0.00 | 0.06 | - | 0.03 | 0.01 | 0.18 | 0.05 | 2.47 | 0.18 | 1.41 |
| Umbelopsis vinacea | Mucoromycota | KC489498 | 628/633 (99) | 0.46 | 0.91 | 0.42 | 0.33 | 1.39 | 3.49 | 1.11 | 2.53 | 0.27 | 0.13 | 0.96 | 1.75 | 1.33 |
| Cladosporium cladosporioides | Ascomycota | MG664765 | 547/547 (100) | 4.28 | 0.70 | 1.22 | 4.16 | 0.22 | 0.14 | 0.49 | 0.27 | 1.12 | 0.54 | 1.22 | 1.23 | 1.22 |
| Suillus granulatus | Basidiomycota | AJ272409 | 677/678 (99) | - | - | - | - | 0.01 | - | 6.83 | 0.01 | 0.03 | - | 2.18 | 0.00 | 1.17 |
| Solicoccozyma terricola | Basidiomycota | KY558367 | 641/641 (100) | 0.50 | 2.89 | 0.15 | 1.10 | 0.26 | 3.52 | 0.29 | 1.53 | 0.12 | 1.84 | 0.29 | 2.14 | 1.15 |
| Phialocephala fortinii | Ascomycota | AY078131 | 560/561 (99) | 0.96 | 0.19 | 0.27 | 0.09 | 1.61 | 1.42 | 2.34 | 0.35 | 1.22 | 0.05 | 1.59 | 0.50 | 1.08 |
| Tuber sp. 3993_24 | Ascomycota | KT215193 | 646/646 (100) | 0.03 | 0.02 | 0.04 | 0.00 | 2.35 | 0.06 | 2.64 | 0.00 | 5.20 | 0.03 | 2.10 | 0.02 | 1.13 |
| Dactylonectria macrodidyma | Ascomycota | JN859422 | 541/541 (100) | 0.63 | 2.10 | 0.38 | 0.69 | 0.46 | 0.90 | 2.17 | 0.89 | 1.22 | 1.04 | 1.10 | 1.03 | 1.06 |
| Mortierella sp. 3993_26 | Mucoromycota | KP311420 | 641/641 (100) | 0.99 | 1.10 | 0.31 | 0.98 | 0.78 | 1.53 | 0.97 | 1.02 | 0.30 | 1.97 | 0.80 | 1.26 | 1.02 |
| Saitozyma podzolica | Basidiomycota | KY320605 | 511/511 (100) | 1.00 | 1.56 | 0.11 | 1.48 | 0.34 | 1.75 | 0.26 | 1.97 | 0.21 | 1.14 | 0.41 | 1.65 | 0.99 |
| Unidentified sp. 3993_29 | Ascomycota | FN393100 | 546/550 (99) | 0.34 | 0.12 | 0.34 | 0.56 | 1.33 | 0.40 | 1.90 | 1.01 | 0.91 | 0.41 | 1.22 | 0.57 | 0.92 |
| Pseudogymnoascus sp. 3993_33 | Ascomycota | KY977601 | 562/562 (100) | 0.19 | 3.84 | 3.21 | 1.01 | 0.33 | 1.39 | 0.22 | 0.59 | 0.46 | 0.67 | 0.48 | 1.33 | 0.87 |
| Penicillium sp. 3993_47 | Ascomycota | MK226541 | 579/579 (100) | 0.29 | 2.54 | 0.04 | 1.05 | 0.29 | 1.04 | 0.43 | 2.24 | 0.12 | 0.80 | 0.30 | 1.54 | 0.87 |
| Mortierella sp. 3993_38 | Mucoromycota | HG935763 | 640/640 (100) | 1.43 | 1.77 | - | 0.74 | 0.34 | 1.42 | 0.39 | 1.36 | 0.15 | 1.29 | 0.52 | 1.28 | 0.87 |
| Trichoderma crassum | Ascomycota | NR134370 | 610/610 (100) | 0.97 | 0.98 | 0.15 | 0.74 | 0.51 | 0.88 | 0.79 | 1.22 | 1.64 | 0.70 | 0.76 | 0.93 | 0.84 |
| Fusarium solani | Ascomycota | MF782768 | 562/562 (100) | 0.14 | 3.87 | 0.08 | 1.09 | 0.31 | 1.65 | 0.31 | 0.82 | 0.12 | 1.45 | 0.25 | 1.57 | 0.86 |
| Pestalotiopsis sp. 3993_40 | Ascomycota | KT963804 | 588/588 (100) | 1.65 | 0.70 | 0.08 | 0.52 | 0.26 | 0.92 | 0.26 | 1.77 | 0.49 | 1.68 | 0.53 | 1.12 | 0.80 |
| Fusarium sp. 3993_41 | Ascomycota | MH550484 | 559/559 (100) | 1.03 | 0.70 | 1.11 | 1.24 | 0.41 | 1.19 | 0.29 | 1.18 | 0.18 | 0.44 | 0.51 | 1.04 | 0.76 |
| Sebacina sp. 3993_35 | Basidiomycota | JX844771 | 641/643 (99) | 0.03 | 0.00 | 0.27 | 0.03 | 3.67 | 0.00 | 0.06 | 0.48 | 0.06 | - | 1.28 | 0.14 | 0.75 |
| Total of 30 OTUs | 46.51 | 39.84 | 52.33 | 43.31 | 73.83 | 39.56 | 65.78 | 41.48 | 44.15 | 22.50 | 62.03 | 38.95 | 51.3 | |||
| OTU | Reference | Similarity % a | Alnus glutinosa | Betula pendula | Picea abies | Pinus sylvestris | Quercus robur | All | All | Total % | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Roots % | Soil % | Roots % | Soil % | Roots % | Soil % | Roots % | Soil % | Roots % | Soil % | Roots % | Soil % | ||||
| Pythium ultimum var. ultimum | AY598657 | 917/917 (100) | 0.258 | 0.559 | 0.076 | 1.435 | 0.048 | 0.829 | 0.067 | 1.349 | 0.030 | 1.554 | 0.094 | 1.161 | 0.59 |
| Pythium heterothallicum | AY598654 | 882/889 (99) | 0.143 | 0.210 | 0.076 | 0.545 | - | 0.251 | 0.008 | 0.812 | 0.030 | 0.829 | 0.038 | 0.534 | 0.27 |
| Pythium spiculum | HQ643790 | 972/978 (99) | 0.072 | 0.047 | 0.038 | 0.040 | 0.032 | 0.088 | 1.243 | 0.114 | - | - | 0.423 | 0.068 | 0.26 |
| Pythium sylvaticum | AY598645 | 997/999 (99) | 0.172 | 0.326 | 0.496 | 0.385 | 0.088 | 0.138 | 0.243 | 0.057 | 0.030 | 0.466 | 0.177 | 0.238 | 0.21 |
| Pythium irregulare | AY598702 | 1026/1029 (99) | 0.043 | 0.280 | 0.076 | 0.133 | 0.128 | 0.301 | 0.151 | 0.114 | 0.091 | 0.207 | 0.113 | 0.198 | 0.15 |
| Peronospora sp. 3993_148 | MF372507 | 803/852 (94) | - | - | - | - | - | - | - | - | 2.765 | - | 0.244 | - | 0.13 |
| Pythium intermedium | KU211482 | 957/959 (99) | - | 0.489 | 0.038 | 0.133 | - | 0.038 | 0.059 | 0.057 | - | 0.026 | 0.021 | 0.124 | 0.07 |
| Pythium amasculinum | AY598671 | 856/857 (99) | 0.029 | - | - | 0.159 | 0.008 | 0.025 | 0.008 | 0.023 | - | 0.363 | 0.011 | 0.093 | 0.05 |
| Pythium sp. 3993_349 | KU211471 | 894/907 (99) | 0.014 | 0.047 | - | 0.053 | 0.024 | - | 0.008 | 0.057 | - | 0.363 | 0.013 | 0.077 | 0.04 |
| Unidentified sp. 3993_508 | MF570293 | 101/119 (85) | 0.029 | 0.047 | 0.038 | 0.040 | 0.008 | 0.050 | 0.025 | 0.069 | 0.030 | 0.104 | 0.021 | 0.059 | 0.04 |
| Pythium acanthicum | AY598617 | 858/859 (99) | 0.014 | - | - | 0.013 | 0.040 | 0.050 | 0.017 | 0.057 | 0.030 | 0.104 | 0.024 | 0.043 | 0.03 |
| Pythium apiculatum | HQ643443 | 948/954 (99) | 0.072 | 0.023 | - | 0.120 | - | 0.063 | - | 0.011 | 0.061 | - | 0.019 | 0.049 | 0.03 |
| Pythium rostratifingens | KU211363 | 1053/1064 (99) | 0.115 | 0.023 | 0.038 | 0.066 | - | - | - | 0.034 | 0.030 | 0.104 | 0.027 | 0.040 | 0.03 |
| Phytopythium citrinum | HM061322 | 852/857 (99) | - | - | 0.496 | - | - | - | 0.008 | - | 0.030 | - | 0.040 | - | 0.02 |
| Unidentified sp. 3993_709 | KJ716873 | 724/865 (84) | - | - | - | - | 0.016 | - | 0.109 | 0.011 | - | - | 0.040 | 0.003 | 0.02 |
| Pythium pleroticum | AY598642 | 958/959 (99) | - | 0.093 | - | - | - | 0.063 | - | - | - | 0.078 | - | 0.037 | 0.02 |
| Pythium sp. 3993_1159 | AY598639 | 940/966 (97) | 0.086 | - | - | - | - | 0.075 | - | - | - | - | 0.016 | 0.019 | 0.02 |
| Unidentified sp. 3993_729 | HQ643756 | 226/240 (94) | - | - | - | - | 0.064 | 0.013 | 0.008 | 0.023 | - | - | 0.024 | 0.009 | 0.02 |
| Pythium rostratifingens | KU209835 | 962/969 (99) | - | 0.047 | - | 0.053 | - | 0.050 | - | - | - | - | - | 0.031 | 0.01 |
| Pythium sp. 3993_1163 | AY598696 | 993/1036 (96) | 0.043 | - | - | - | - | 0.013 | - | 0.034 | - | 0.078 | 0.008 | 0.022 | 0.01 |
| Unidentified sp. 3993_943 | MH671329 | 870/883 (99) | - | - | - | - | 0.048 | 0.025 | - | 0.011 | - | 0.026 | 0.016 | 0.012 | 0.01 |
| Unidentified sp. 3993_1191 | KF318041 | 606/754 (80) | - | 0.023 | - | - | - | - | - | 0.046 | - | 0.104 | - | 0.028 | 0.01 |
| Phytophthora fragariae | KJ755093 | 896/905 (99) | 0.029 | 0.116 | 0.038 | - | - | - | - | - | - | - | 0.008 | 0.015 | 0.01 |
| Phytophthora pseudosyringae | EU074793 | 838/848 (99) | - | - | - | - | - | - | - | 0.011 | - | 0.181 | - | 0.025 | 0.01 |
| Pythium violae | AY598717 | 1001/1006 (99) | 0.043 | 0.023 | - | 0.013 | - | 0.025 | - | 0.011 | - | - | 0.008 | 0.015 | 0.01 |
| Unidentified sp. 3993_1954 | LC176476 | 143/157 (91) | 0.057 | - | - | 0.053 | - | - | - | - | - | - | 0.011 | 0.012 | 0.01 |
| Unidentified sp. 3993_981 | KF318041 | 602/754 (80) | - | 0.023 | - | 0.040 | - | - | - | - | - | 0.104 | - | 0.025 | 0.01 |
| Pythium sp. 3993_1117 | JF431913 | 804/842 (95) | - | - | - | 0.013 | - | 0.050 | - | - | - | 0.052 | - | 0.022 | 0.01 |
| Unidentified sp. 3993_1171 | KJ716873 | 797/854 (93) | 0.014 | - | - | 0.040 | - | - | - | - | - | 0.078 | 0.003 | 0.019 | 0.01 |
| Hyaloperonospora brassicae | MG757782 | 940/943 (99) | - | - | - | - | - | - | 0.050 | - | - | - | 0.016 | - | 0.01 |
| Total of 30 OTUs | 1.232 | 2.377 | 1.412 | 3.336 | 0.503 | 2.148 | 2.007 | 2.905 | 3.130 | 4.816 | 1.414 | 2.977 | 2.14 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marčiulynienė, D.; Marčiulynas, A.; Lynikienė, J.; Vaičiukynė, M.; Gedminas, A.; Menkis, A. DNA-Metabarcoding of Belowground Fungal Communities in Bare-Root Forest Nurseries: Focus on Different Tree Species. Microorganisms 2021, 9, 150. https://doi.org/10.3390/microorganisms9010150
Marčiulynienė D, Marčiulynas A, Lynikienė J, Vaičiukynė M, Gedminas A, Menkis A. DNA-Metabarcoding of Belowground Fungal Communities in Bare-Root Forest Nurseries: Focus on Different Tree Species. Microorganisms. 2021; 9(1):150. https://doi.org/10.3390/microorganisms9010150
Chicago/Turabian StyleMarčiulynienė, Diana, Adas Marčiulynas, Jūratė Lynikienė, Miglė Vaičiukynė, Artūras Gedminas, and Audrius Menkis. 2021. "DNA-Metabarcoding of Belowground Fungal Communities in Bare-Root Forest Nurseries: Focus on Different Tree Species" Microorganisms 9, no. 1: 150. https://doi.org/10.3390/microorganisms9010150
APA StyleMarčiulynienė, D., Marčiulynas, A., Lynikienė, J., Vaičiukynė, M., Gedminas, A., & Menkis, A. (2021). DNA-Metabarcoding of Belowground Fungal Communities in Bare-Root Forest Nurseries: Focus on Different Tree Species. Microorganisms, 9(1), 150. https://doi.org/10.3390/microorganisms9010150