Molecular Methods for the Detection of Toxoplasma gondii Oocysts in Fresh Produce: An Extensive Review

 , , , and
, , , and
Abstract
:1. Introduction
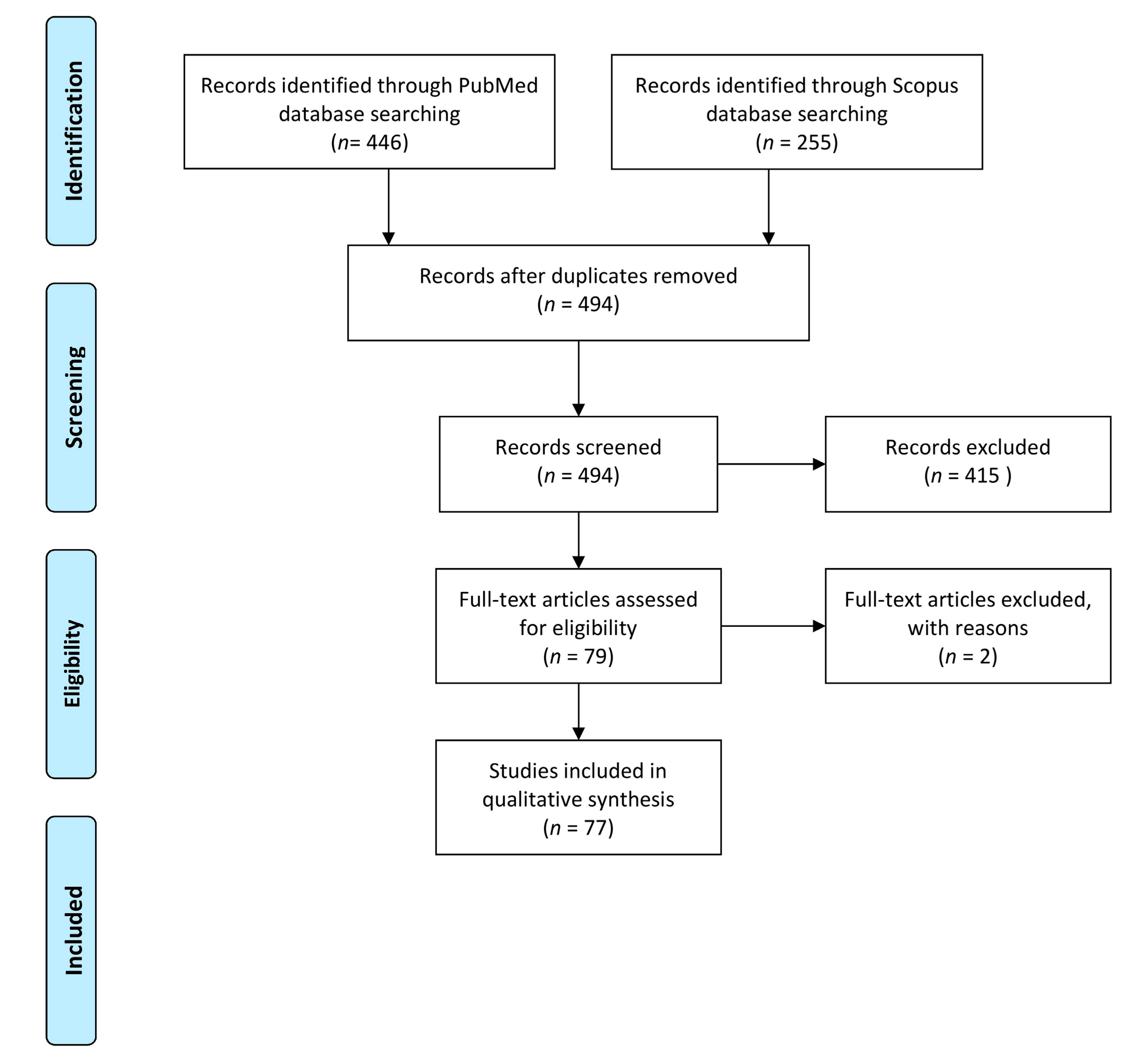
2. Materials and Methods
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dubey, J.P. Toxoplasmosis of Animals and Humans, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Torgerson, P.R.; Devleesschauwer, B.; Praet, N.; Speybroeck, N.; Willingham, A.L.; Kasuga, F.; Rokni, M.B.; Zhou, X.N.; Fèvre, E.M.; Sripa, B.; et al. World Health Organization Estimates of the Global and Regional Disease Burden of 11 Foodborne Parasitic Diseases, 2010: A Data Synthesis. PLoS Med. 2015, 12, e1001920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ECDC. European Centre for Disease Prevention and Control. Congenital toxoplasmosis. In Annual Epidemiological Report for 2015; ECDC: Stockholm, Sweden, 2018. [Google Scholar]
- Hald, T.; Aspinall, W.; Devleesschauwer, B.; Cooke, R.; Corrigan, T.; Havelaar, A.H.; Gibb, H.J.; Torgerson, P.R.; Kirk, M.D.; Angulo, F.J.; et al. World Health Organization Estimates of the Relative Contributions of Food to the Burden of Disease Due to Selected Foodborne Hazards: A Structured Expert Elicitation. PLoS ONE 2016, 11, e0145839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA Panel on Biological Hazards (BIOHAZ); Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Herman, L.; et al. Public health risks associated with food-borne parasites. Efsa J. 2018, 16, e05495. [Google Scholar] [CrossRef] [PubMed]
- Bouwknegt, M.; Devleesschauwer, B.; Graham, H.; Robertson, L.J.; van der Giessen, J.W. The Euro-Fbp Workshop Participants. Prioritisation of food-borne parasites in Europe, 2016. Eurosurveillance 2018, 23, 17-00161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food and Agriculture Organization of the United Nations/World Health Organization. Multicriteria Based Ranking for Risk Management of Food Borne Parasites; FAO, World Health Organization: Rome, Italy, 2014; 287p. [Google Scholar]
- Castro-Ibáñez, I.; Gil, M.I.; Allende, A. Ready-to-eat vegetables: Current problems and potential solutions to reduce microbial risk in the production chain. LWT Food Sci. Technol. 2017, 85, 284–292. [Google Scholar] [CrossRef]
- Murphy, H.R.; Almeria, S.; da Silva, A.J. BAM Chapter 19b: Molecular Detection of Cyclospora cayetanensis in Fresh Produce Using Real-Time PCR. U.S: Food and Drug Administration. 2019. Available online: https://www.fda.gov/food/laboratory-methods-food/bam-chapter-19b-molecular-detection-cyclospora-cayetanensis-fresh-produce-using-real-time-pcr (accessed on 10 July 2020).
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. The PRISMA Group. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [Green Version]
- Lalle, M.; Slana, I.; Bier, N.; Mayer-Scholl, A.; Jokelainen, P. Deliverable D-JRP-TOXOSOURCES-WP3.1 Report on Available Analytical Procedures for Detection of Toxoplasma Gondii in Fresh Produce and List of Promising Analytical Procedures. April 2020. Available online: https://zenodo.org/record/3778719#.X8ENmbPSLcs (accessed on 10 July 2020).
- Chandra, V.; Torres, M.; Ortega, Y.R. Efficacy of wash solutions in recovering Cyclospora cayetanensis, Cryptosporidium parvum, and Toxoplasma gondii from basil. J. Food Prot. 2014, 77, 1348–1354. [Google Scholar] [CrossRef]
- Hohweyer, J.; Cazeaux, C.; Travaillé, E.; Languet, E.; Dumètre, A.; Aubert, D.; Terryn, C.; Dubey, J.P.; Azas, N.; Houssin, M.; et al. Simultaneous detection of the protozoan parasites Toxoplasma, Cryptosporidium and Giardia in food matrices and their persistence on basil leaves. Food Microbiol. 2016, 57, 36–44. [Google Scholar] [CrossRef]
- Lalle, M.; Possenti, A.; Dubey, J.P.; Pozio, E. Loop-Mediated Isothermal Amplification-Lateral-Flow Dipstick (LAMP-LFD) to detect Toxoplasma gondii oocyst in ready-to-eat salad. Food Microbiol. 2018, 70, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, K.; Kim, M.; Rajal, V.B.; Arrowood, M.J.; Packham, A.; Aguilar, B.; Wuertz, S. Simultaneous detection of four protozoan parasites on leafy greens using a novel multiplex PCR assay. Food Microbiol. 2019, 84, 103252. [Google Scholar] [CrossRef] [PubMed]
- de Souza, C.Z.; Rafael, K.; Sanders, A.P.; Tiyo, B.T.; Marchioro, A.A.; Colli, C.M.; Gomes, M.L.; Falavigna-Guilherme, A.L. An alternative method to recover Toxoplasma gondii from greenery and fruits. Int. J. Environ. Health Res. 2016, 26, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, L.F.; Gajadhar, A.A. Optimization and validation of methods for isolation and real-time PCR identification of protozoan oocysts on leafy green vegetables and berry fruits. Food Waterborne Parasitol. 2016, 2, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Marchioro, A.A.; Tiyo, B.T.; Colli, C.M.; de Souza, C.Z.; Garcia, J.L.; Gomes, M.L.; Falavigna-Guilherme, A.L. First Detection of Toxoplasma gondii DNA in the Fresh Leafs of Vegetables in South America. Vector Borne Zoonotic Dis. 2016, 16, 624–626. [Google Scholar] [CrossRef] [PubMed]
- Lass, A.; Pietkiewicz, H.; Szostakowska, B.; Myjak, P. The first detection of Toxoplasma gondii DNA in environmental fruits and vegetables samples. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1101–1108. [Google Scholar] [CrossRef] [Green Version]
- Temesgen, T.T.; Robertson, L.J.; Tysnes, K.R. A novel multiplex real-time PCR for the detection of Echinococcus multilocularis, Toxoplasma gondii, and Cyclospora cayetanensis on berries. Food Res. Int. 2019, 125, 108636. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, L.F.; Gajadhar, A.A. Detection of Cyclospora cayetanensis, Cryptosporidium spp., and Toxoplasma gondii on imported leafy green vegetables in Canadian survey. Food Waterborne Parasitol. 2016, 2, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Caradonna, T.; Marangi, M.; Del Chierico, F.; Ferrari, N.; Reddel, S.; Bracaglia, G.; Normanno, G.; Putignani, L.; Giangaspero, A. Detection and prevalence of protozoan parasites in ready-to-eat packaged salads on sale in Italy. Food Microbiol. 2017, 67, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Lass, A.; Ma, L.; Kontogeorgos, I.; Zhang, X.; Li, X.; Karanis, P. First molecular detection of Toxoplasma gondii in vegetable samples in China using qualitative, quantitative real-time PCR and multilocus genotyping. Sci. Rep. 2019, 26, 17581. [Google Scholar] [CrossRef] [PubMed]
- Slany, M.; Dziedzinska, R.; Babak, V.; Kralik, P.; Moravkova, M.; Slana, I. Toxoplasma gondii in vegetables from fields and farm storage facilities in the Czech Republic. FEMS Microbiol. Lett. 2019, 366, fnz170. [Google Scholar] [CrossRef]
- Herrmann, D.C.; Maksimov, A.; Pantchev, N.; Vrhovec, M.G.; Conraths, F.J.; Schares, G. Comparison of different commercial DNA extraction kits to detect Toxoplasma gondii oocysts in cat faeces. Berl Munch Tierarztl. Wochenschr. 2011, 124, 497–502. [Google Scholar] [PubMed]
- Staggs, S.E.; Keely, S.P.; Ware, M.W.; Schable, N.; See, M.J.; Gregorio, D.; Zou, X.; Su, C.; Dubey, J.P.; Villegas, E.N. The development and implementation of a method using blue mussels (Mytilus spp.) as biosentinels of Cryptosporidium spp. and Toxoplasma gondii contamination in marine aquatic environments. Parasitol. Res. 2015, 114, 4655–4667. [Google Scholar] [CrossRef] [PubMed]
- Manore, A.J.W.; Harper, S.L.; Aguilar, B.; Weese, J.S.; Shapiro, K. Comparison of freeze-thaw cycles for nucleic acid extraction and molecular detection of Cryptosporidium parvum and Toxoplasma gondii oocysts in environmental matrices. J. Microbiol. Methods 2019, 156, 1–4. [Google Scholar] [CrossRef]
- Durand, L.; La Carbona, S.; Geffard, A.; Possenti, A.; Dubey, J.P.; Lalle, M. Comparative evaluation of loop-mediated isothermal amplification (LAMP) vs qPCR for detection of Toxoplasma gondii oocysts DNA in mussels. Exp. Parasitol. 2020, 208, 107809. [Google Scholar] [CrossRef] [PubMed]
- Géba, E.; Aubert, D.; Durand, L.; Escotte, S.; La Carbona, S.; Cazeaux, C.; Bonnard, I.; Bastien, F.; Palos Ladeiro, M.; Dubey, J.P.; et al. Use of the bivalve Dreissena polymorpha as a biomonitoring tool to reflect the protozoan load in freshwater bodies. Water Res. 2020, 170, 115297. [Google Scholar] [CrossRef] [PubMed]
- Escotte-Binet, S.; Da Silva, A.M.; Cancès, B.; Aubert, D.; Dubey, J.; La Carbona, S.; Villena, I.; Poulle, M.L. A rapid and sensitive method to detect Toxoplasma gondii oocysts in soil samples. Vet. Parasitol. 2019, 274, 108904. [Google Scholar] [CrossRef]
- Galvani, A.T.; Christ, A.P.G.; Padula, J.A.; Barbosa, M.R.F.; de Araújo, R.S.; Sato, M.I.Z.; de Araújo, R.S.; Sato, M.I.Z.; Razzolini, M.T.P. Real-time PCR detection of Toxoplasma gondii in surface water samples in São Paulo, Brazil. Parasitol. Res. 2019, 118, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, L.A.; Santos, L.K.; Brito, P.A., Jr.; Maciel, B.M.; Da Silva, A.V.; Albuquerque, G.R. Detection of Toxoplasma gondii DNA in Brazilian oysters (Crassostrea rhizophorae). Genet. Mol. Res. 2015, 14, 4658–4665. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Lindquist, H.D.; Cama, V.; Schaefer, F.W., 3rd; Villegas, E.; Fayer, R.; Lewis, E.J.; Feng, Y.; Xiao, L. Detection of Toxoplasma gondii oocysts in water sample concentrates by real-time PCR. Appl. Environ. Microbiol. 2009, 75, 3477–3483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lass, A.; Pietkiewicz, H.; Modzelewska, E.; Dumètre, A.; Szostakowska, B.; Myjak, P. Detection of Toxoplasma gondii oocysts in environmental soil samples using molecular methods. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Salant, H.; Markovics, A.; Spira, D.T.; Hamburger, J. The development of a molecular approach for coprodiagnosis of Toxoplasma gondii. Vet. Parasitol. 2007, 146, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Tavalla, M.; Oormazdi, H.; Akhlaghi, L.; Shojaee, S.; Razmjou, E.; Hadighi, R.; Meamar, A. Genotyping of Toxoplasma gondii Isolates from Soil Samples in Tehran, Iran. Iran. J. Parasitol. 2013, 8, 227–233. [Google Scholar]
- Du, F.; Feng, H.L.; Nie, H.; Tu, P.; Zhang, Q.L.; Hu, M.; Zhou, Y.Q.; Zhao, J.L. Survey on the contamination of Toxoplasma gondii oocysts in the soil of public parks of Wuhan, China. Vet. Parasitol. 2012, 184, 141–146. [Google Scholar] [CrossRef]
- Schares, G.; Vrhovec, M.G.; Pantchev, N.; Herrmann, D.C.; Conraths, F.J. Occurrence of Toxoplasma gondii and Hammondia hammondi oocysts in the faeces of cats from Germany and other European countries. Vet. Parasitol. 2008, 152, 34–45. [Google Scholar] [CrossRef]
- Matsuo, J.; Kimura, D.; Rai, S.K.; Uga, S. Detection of Toxoplasma oocysts from soil by modified sucrose flotation and PCR methods. Southeast. Asian J. Trop Med. Public Health 2004, 35, 270–274. [Google Scholar] [PubMed]
- Chemoh, W.; Sawangjaroen, N.; Nissapatorn, V.; Sermwittayawong, N. Molecular investigation on the occurrence of Toxoplasma gondii oocysts in cat feces using TOX-element and ITS-1 region targets. Vet. J. 2016, 215, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, D.C.; Pantchev, N.; Vrhovec, M.G.; Barutzki, D.; Wilking, H.; Fröhlich, A.; Lüder, C.G.; Conraths, F.J.; Schares, G. Atypical Toxoplasma gondii genotypes identified in oocysts shed by cats in Germany. Int. J. Parasitol. 2010, 40, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Lélu, M.; Gilot-Fromont, E.; Aubert, D.; Richaume, A.; Afonso, E.; Dupuis, E.; Gotteland, C.; Marnef, F.; Poulle, M.L.; Dumètre, A.; et al. Development of a sensitive method for Toxoplasma gondii oocyst extraction in soil. Vet. Parasitol. 2011, 183, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Villena, I.; Aubert, D.; Gomis, P.; Ferté, H.; Inglard, J.C.; Denis-Bisiaux, H.; Dondon, J.M.; Pisano, E.; Ortis, N.; Pinon, J.M. Evaluation of a strategy for Toxoplasma gondii oocyst detection in water. Appl. Environ. Microbiol. 2004, 70, 4035–4039. [Google Scholar] [CrossRef] [Green Version]
- de Wit, L.A.; Kilpatrick, A.M.; VanWormer, E.; Croll, D.A.; Tershy, B.R.; Kim, M.; Shapiro, K. Seasonal and spatial variation in Toxoplasma gondii contamination in soil in urban public spaces in California, United States. Zoonoses Public Health 2020, 67, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Wells, B.; Shaw, H.; Innocent, G.; Guido, S.; Hotchkiss, E.; Parigi, M.; Opsteegh, M.; Green, J.; Gillespie, S.; Innes, E.A.; et al. Molecular detection of Toxoplasma gondii in water samples from Scotland and a comparison between the 529bp real-time PCR and ITS1 nested PCR. Water Res. 2015, 87, 175–181. [Google Scholar] [CrossRef] [Green Version]
- Poulle, M.L.; Bastien, M.; Richard, Y.; Josse-Dupuis, É.; Aubert, D.; Villena, I.; Knapp, J. Detection of Echinococcus multilocularis and other foodborne parasites in fox, cat and dog faeces collected in kitchen gardens in a highly endemic area for alveolar echinococcosis. Parasite 2017, 24, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigot-Clivot, A.; Palos Ladeiro, M.; Lepoutre, A.; Bastien, F.; Bonnard, I.; Dubey, J.P.; Villena, I.; Aubert, D.; Geffard, O.; François, A.; et al. Bioaccumulation of Toxoplasma and Cryptosporidium by the freshwater crustacean Gammarus fossarum: Involvement in biomonitoring surveys and trophic transfer. Ecotoxicol. Environ. Saf. 2016, 133, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Wang, H.; Wang, H.; Qin, H.; Xiao, J. Land use and soil contamination with Toxoplasma gondii oocysts in urban areas. Sci. Total Environ. 2016, 568, 1086–1091. [Google Scholar] [CrossRef]
- Poulle, M.L.; Forin-Wiart, M.A.; Josse-Dupuis, É.; Villena, I.; Aubert, D. Detection of Toxoplasma gondii DNA by qPCR in the feces of a cat that recently ingested infected prey does not necessarily imply oocyst shedding. Parasite 2016, 23, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palos Ladeiro, M.; Bigot-Clivot, A.; Aubert, D.; Villena, I.; Geffard, A. Assessment of Toxoplasma gondii levels in zebra mussel (Dreissena polymorpha) by real-time PCR: An organotropism study. Environ. Sci. Pollut. Res. Int. 2015, 22, 13693–13701. [Google Scholar]
- Bier, N.S.; Schares, G.; Johne, A.; Martin, A.; Nöckler, K.; Mayer-Scholl, A. Performance of three molecular methods for detection of Toxoplasma gondii in pork. Food Waterborne Parasitol. 2019, 14, e00038. [Google Scholar] [CrossRef] [PubMed]
- Gotteland, C.; Gilot-Fromont, E.; Aubert, D.; Poulle, M.L.; Dupuis, E.; Dardé, M.L.; Forin-Wiart, M.A.; Rabilloud, M.; Riche, B.; Villena, I. Spatial distribution of Toxoplasma gondii oocysts in soil in a rural area: Influence of cats and land use. Vet. Parasitol. 2014, 205, 629–637. [Google Scholar] [CrossRef]
- Afonso, E.; Lemoine, M.; Poulle, M.L.; Ravat, M.C.; Romand, S.; Thulliez, P.; Villena, I.; Aubert, D.; Rabilloud, M.; Riche, B.; et al. Spatial distribution of soil contamination by Toxoplasma gondii in relation to cat defecation behaviour in an urban area. Int. J. Parasitol. 2008, 38, 1017–1023. [Google Scholar] [CrossRef]
- Aubert, D.; Villena, I. Detection of Toxoplasma gondii oocysts in water: Proposition of a strategy and evaluation in Champagne-Ardenne Region, France. Mem Inst. Oswaldo Cruz. 2009, 104, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Sroka, J.; Karamon, J.; Dutkiewicz, J.; Wójcik-Fatla, A.; Cencek, T. Optimization of flotation, DNA extraction and PCR methods for detection of Toxoplasma gondii oocysts in cat faeces. Ann. Agric. Environ. Med. 2018, 25, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Reischl, U.; Bretagne, S.; Krüger, D.; Ernault, P.; Costa, J.M. Comparison of two DNA targets for the diagnosis of Toxoplasmosis by real-time PCR using fluorescence resonance energy transfer hybridization probes. BMC Infect. Dis. 2003, 3, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, K.; Mazet, J.A.; Schriewer, A.; Wuertz, S.; Fritz, H.; Miller, W.A.; Largier, J.; Conrad, P.A. Detection of Toxoplasma gondii oocysts and surrogate microspheres in water using ultrafiltration and capsule filtration. Water Res. 2010, 44, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Arkush, K.D.; Miller, M.A.; Leutenegger, C.M.; Gardner, I.A.; Packham, A.E.; Heckeroth, A.R.; Tenter, A.M.; Barr, B.C.; Conrad, P.A. Molecular and bioassay-based detection of Toxoplasma gondii oocyst uptake by mussels (Mytilus galloprovincialis). Int. J. Parasitol. 2003, 33, 1087–1097. [Google Scholar] [CrossRef]
- Marquis, N.D.; Bishop, T.J.; Record, N.R.; Countway, P.D.; Fernández Robledo, J.A. Molecular Epizootiology of Toxoplasma gondii and Cryptosporidium parvum in the Eastern Oyster (Crassostrea virginica) from Maine (USA). Pathogens 2019, 8, 125. [Google Scholar] [CrossRef] [Green Version]
- Coupe, A.; Howe, L.; Shapiro, K.; Roe, W.D. Comparison of PCR assays to detect Toxoplasma gondii oocysts in green-lipped mussels (Perna canaliculus). Parasitol. Res. 2019, 118, 2389–2398. [Google Scholar] [CrossRef] [PubMed]
- Opsteegh, M.; Langelaar, M.; Sprong, H.; den Hartog, L.; De Craeye, S.; Bokken, G.; Ajzenberg, D.; Kijlstra, A.; van der Giessen, J. Direct detection and genotyping of Toxoplasma gondii in meat samples using magnetic capture and PCR. Int. J. Food Microbiol. 2010, 139, 193–201. [Google Scholar] [CrossRef]
- Aksoy, U.; Marangi, M.; Papini, R.; Ozkoc, S.; Bayram Delibas, S.; Giangaspero, A. Detection of Toxoplasma gondii and Cyclospora cayetanensis in Mytilus galloprovincialis from Izmir Province coast (Turkey) by Real Time PCR/High-Resolution Melting analysis (HRM). Food Microbiol. 2014, 44, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Marangi, M.; Giangaspero, A.; Lacasella, V.; Lonigro, A.; Gasser, R.B. Multiplex PCR for the detection and quantification of zoonotic taxa of Giardia, Cryptosporidium and Toxoplasma in wastewater and mussels. Mol. Cell Probes. 2015, 29, 122–125. [Google Scholar] [CrossRef] [PubMed]
- Chalmers, R.; Robertson, L.; Dorny, P.; Suzanne, J.; Kärssin, A.; Katzer, F.; La Carbona, S.; Lalle, M.; Lassen, B.; Mladineo, I.; et al. Parasite detection in food: Current status and future needs for validation. Trends Food Sci. Technol. 2020, 99, 337–350. [Google Scholar] [CrossRef]
- Kourenti, C.; Karanis, P. Development of a sensitive polymerase chain reaction method for the detection of Toxoplasma gondii in water. Water Sci. Technol. 2004, 50, 287–291. [Google Scholar] [CrossRef]
- Cook, N.; Paton, C.A.; Wilkinson, N.; Nichols, R.A.; Barker, K.; Smith, H.V. Towards standard methods for the detection of Cryptosporidium parvum on lettuce and raspberries. Part 1: Development and optimization of methods. Int. J. Food Microbiol. 2006, 109, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Schrader, C.; Schielke, A.; Ellerbroek, L.; Johne, R. PCR inhibitors—Occurrence, properties and removal. J. Appl. Microbiol. 2012, 113, 1014–1026. [Google Scholar] [CrossRef] [PubMed]
- International Standards Organisation. Microbiology of Food and Animal Feeding Stuffs–Polymerase Chain Reaction (PCR) for the Detection of Food-Borne Pathogens—General Requirements and Definitions (ISO 22174:2005); International Standards Organisation: Geneva, Switzerland, 2005. [Google Scholar]
- Hoorfar, J.; Malorny, B.; Abdulmawjood, A.; Cook, N.; Wagner, M.; Fach, P. Practical considerations in design of internal amplification controls for diagnostic PCR assays. J. Clin. Microbiol. 2004, 42, 1863–1868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustin, S.A. Why the need for qPCR publication guidelines?—The case for MIQE. Methods 2010, 50, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.; Wakem, M.; Dijkman, G.; Alsarraj, M.; Nguyen, M. A practical approach to RT-qPCR-Publishing data that conform to the MIQE guidelines. Methods 2010, 50, S1–S5. [Google Scholar] [CrossRef] [PubMed]
- International Standards Organisation. Microbiology of Food and Animal Feeding Stuffs—Polymerase Chain Reaction (PCR) for the Detection of Food-Borne Pathogens (ISO 22174:2005); International Standards Organisation: Geneva, Switzerland, 2005. [Google Scholar]
- International Standards Organisation. Microbiology of Food and Animal Feeding Stuffs—Polymerase Chain Reaction (PCR) for the Detection of Food-Borne Pathogens—Performance Testing for Thermal Cyclers (ISO/TS 20836:2005); International Standards Organisation: Geneva, Switzerland, 2005. [Google Scholar]
- International Standards Organisation. Microbiology of Food and Animal Feeding Stuffs—Polymerase Chain Reaction (PCR) for the Detection of Food-Borne Pathogens—Requirements for Sample Preparation for Qualitative Detection (ISO 20837:2006); International Standards Organisation: Geneva, Switzerland, 2006. [Google Scholar]
- International Standards Organisation. Microbiology of Food and Animal Feeding Stuffs—Polymerase Chain Reaction (PCR) for the Detection of Food-Borne Pathogens—Requirements for Amplification and Detection for Qualitative Methods (ISO 20838:2006); International Standards Organisation: Geneva, Switzerland, 2006. [Google Scholar]
- International Standards Organisation. Microbiology of Food and Animal Feeding Stuffs—General Requirements and Guidance for Microbiological Examinations (ISO 7218:2007 and Amendments 2013); International Standards Organisation: Geneva, Switzerland, 2013. [Google Scholar]
- International Standards Organisation. Microbiology of the Food Chain—Method Validation—Part. 2: Protocol for the Validation of Alternative (Proprietary) Methods against a Reference Method (ISO 16140-2:2016); International Standards Organisation: Geneva, Switzerland, 2016. [Google Scholar]
- Food and Drug Administration (US); Foods Program Regulatory Science Steering Committee (RSSC). Guidelines for the Validation of Analytical Methods for the Detection of Microbial Pathogens in Foods and Feeds, 3rd ed.; U.S. FDA: Silver Spring, MD, USA, 2019.



{kind=link}
{kind=link}
| Type of Spiking | Matrix (Grams) | Spiking Level (oo)Cysts | Time after Spiking | Processing Method | Washing Buffer (mL) | Recovery (%) (Quantitative Evaluation) ### | Pre-Treatment before DNA Extract | DNA Extraction | Detection Method (Target Gene) | Amplicon Size (bp) | LoD (Oocysts) | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| dripping | basil (25) | 102 | ON at 4 °C | wash by hand shaking for 15 s, hand rubbing for 30 s, shaking vigorously for 15 s and centrifugtion | six different buffers (200 mL) # | NR | BB (5.5 m/s for 30 s) | Fast DNA Spin for Soil kit | PCR (529 RE) | 529 | depend on the washing buffer ## | [12] |
| basil (30) raspberries (30) | 5 to 104 | 2 h at RT | wash by automatic shaker (80 rpm 10 min), centrifugation, IMS Toxo | 1 M glycine pH 5.5 (200 mL) | basil: 0.2% microscopy; 35% qPCR raspberry: 2% microscopy; 29% qPCR | FT 6× (−80 °C for 5 min/95 °C for 5 min) and US (1 min at 37 Hz) | InstaGene Matrix | qPCR Taqman (529 RE) | 81 | Basil: <33/g Raspberries: <33/g | [13] | |
| wash by automatic shaker (80 rpm 10 min), centrifugation | basil: 35% qPCR raspberries: 2.5% qPCR | Basil: <1/g Raspberries: <1/g | ||||||||||
| baby lettuce (50) | 25, 50, 100 for 50 g | ON at 4 °C | wash by stomaching b and centrifugation | 1 M glycine pH 5.5 (200 mL) | NR | BB 2× (6.0 m/s for 40 s) | FastPrep for soil kit | LAMP (529 RE) | NA | 0.5/g or 5/mL | [14] | |
| 5, 10, 50 in 800 µL (pellet) | ON at −20 °C | qPCR_Taqman (529 RE) | 163 | |||||||||
| spinach (10) | 101 to 104 | 2 h at RT | wash by stomaching and centrifugation | 0.1% Tween 80 (100 mL) | ≤30% by microscopy (IMS/membrane filtration) | FT 1× (4 min in N2 and 4 min 100°C) | DNeasy Blood and Tissue Kit | nPCR (rDNA 18S) | 715 | 0.1–1/g | [15] | |
| manual wash and centrifugation | ≥40% by microscopy (IMS/membrane filtration) | qPCR_Taqman (529 RE) | 163 | 1/g | ||||||||
| strawberry (50) lettuce (50) | 101 to 104, in 100 mL ddH2O for 250 g sample | 30 min at RT | wash by manual stirring, filtration through a cellulose ester membrane and centrifugation | 1% Tween 80 (100 mL) | NR | US 5× (45 s at 20 Hz, at 2 min intervals | Axy Prep Blood Genomic DNA kit | PCR (B1 gene) | 115 | 1000/250 g by immersion using any pre-treatment | [16] | |
| 10/250 g by drip using FT or US | ||||||||||||
| blackberries, blueberries, cranberries, raspberries, strawberries (60) herbs a (35) green onions (35) | 5 × 103 Eimeria papillata | ON at 4 °C | wash by orbital shaking in bottles for 1 min, centrifugation and Sheather’s solution in the flotation | 1 M glycine pH 5.5 (200 mL) | higher for blueberries, leafy herbs and thyme | FT 8× (1 min in N2/1 min at 95 °C) and proteinase K | spin columns (QIAamp DNA Mini Kit | qPCR_HRM (18S rDNA) | 312 | For all berry types, 3/g For herbs and green onions, 5–9/g | [17] * | |
| ON at RT | wash by stomaching in stomacher filter bag, centrifugation and Sheather’s solution in the flotation | elution solution 0.563 mM H2Na2P2O7/42.8 mM NaCl (200 mL) | higher for blackberries, cranberries, raspberries and green onions | |||||||||
| wash by shaking in stomacher filter bag (30 min per side), centrifugation and Sheather’s solution in the flotation | ||||||||||||
| immersion | lettuce (50) | 10 to 104, in 2000 mL ddH2O | NR | wash by manual stirring, filtration through a cellulose ester membrane and centrifugation | 1% Tween 80 (100 mL) | NR | FT 5× (5 min in N2/5 min at 65 °C) | spin columns (Axy Prep Blood Genomic DNA | PCR (B1 gene) | 115 | ≥10/ µL | [18] |
| PCR (529 RE) | 529 | ≥100/ µL | ||||||||||
| strawberry (50) lettuce (50) | 101–104, in 2000 mL ddH2O for 250 g sample | stirred 15 s followed by 30 min incubation at RT | wash by manual stirring, filtration through a cellulose ester membrane and centrifugation | 1% Tween 80 (100 mL) | NR | NO | Axy Prep Blood Genomic DNA kit | PCR (529 RE) | 529 | 1000/250 g by immersion using FT or US | [16] | |
| FT 5× (5 min in N2/5 min at 65 °C) | 100/250 g by drip using FT or US | |||||||||||
| NR | radish (NR) strawberries (NR) | 101 to 104 | NR | wash in glass beaker by automatic shaker (100 rpm 2 h), flocculation method using CaCO3 solution ON, centrifugation | 1% Tween 80 (2000 mL) | NR | BB (6.5 m/s for 2 min), chloroform treatment, and proteinase K incubation | FAST-Prep for soil matrix E combined with Genomic Mini Kit | qPCR_Taqman (B1 gene) | 128 | 100/single radish 10,000/single strawberries | [19] |
| NR | raspberries (30) blueberries (30) | 10 or 50 in 250 µL (sediment) | NA | wash by automatic shaker (300-600 rpm 10 min) and centrifugation | 1% Alconox (200 mL) | NA | BB 2× (4 m/s for 60 s) | DNeasy Power Soil | multiplex qPCR_Taqman (529 RE) | 163 | 10/250 µL (sediment from 30 g berries) | [20] |
| Matrix (Number) | Source (Number) | Amount of Sample Tested | Processing Method | Washing Buffer (mL) | Pre-Treatment before DNA Extraction | DNA Extraction | Detection Method (Target Gene) | Amplicon Size (bp) | LoD (Oocysts) | Sample Positive (%) | Reference |
| strawberries (60) carrot (46) radish (60) lettuce (50) | retail shops and marketplaces (175) kitchen-gardens and allotments (41) | 1 kg strawberries, 0,5 kg carrot, 20 radishes, 1 lettuce | wash in glass beaker? by automatic shaker (100 rpm 2 h), flocculation method using CaCO3 solution ON, centrifugation | 1% Tween 80 (2000 mL) | BB (6.5 m/s for 2 min) | Chloroform, proteinase K incubation and Genomic Mini Kit | qPCR_Taqman (B1) | 128 | 1 | 21/216 (10% ) 3 radish 9 carrot 9 lettuce | [19] * |
| arugula/baby arugula (107) kale (44) spinach/baby spinach (387) romaine (113) chard (39) leaf lettuces1 (226) spring mix (124) leafy green mixes (91) other mixes (3) | retail outlets (1171) | 35 g | wash by orbital shaker or stomacher (romaine, red or green leafy lettuces only), centrifugation and flotation procedure using Sheather’s solution | 1 M glycine pH 5.5 (200 mL) | FT 8× (1 min N2/1 min. 95 °C) and proteinase K | QIAamp DNA Micro Kit or Dneasy Blood and Tissue Kit | Multiplex qPCR_HRM (18S rDNA) | 312 | 10 | 3/387 (0.78%) baby spinach | [21] |
| crisp lettuce (106) regular lettuce (62) chicory (40) rocket (7) parsley (5) | open fairs (77) from producers’ fairs (81), from community fairs (80) | 50 g | wash by manual stirring, filtration through a cellulose ester membrane and centrifugation | 1% Tween 80 (100 mL) | FT 5× (5 min N2/5 min. 65 °C) | Axy Prep Blood Genomic DNA | PCR (B1) | 115 | NR | 9/238 (3.8 %) 4 crisp lettuce 1 chicory 1 rocket 1 parsley | [18]* |
| PCR (529 RE) | 529 | NR | 1 chicory 1 regular lettuce | ||||||||
| RTE mixed salad (curly and escarole lettuce, red radish, rocket salad and carrots) (648) | retail shops (648) | 100 g (9 salad packages, 72 pools) | wash by orbital shaker for 15 min at 120 rpm and centrifugation | 10X PBS, 0.1% Tween-80, 0.1% SDS and 0.05% antifoam B (200 mL) | FT 15× (1 min N2/1 min. 95 °C) | QiAmp Plant Mini Kit | qPCR_HRM (B1) | 128 | NR | 0.8% | [22] |
| lettuce (71) spinach (50) pak choi (34) chinese cabbage (26) rape (22) asparagus (18) chrysanthemum coronarium (16) endive (14) chinese chives (11) cabbage (9) red cabbage (8) | open markets | NR | sample rinsing and Al2(SO4)3 flocculation of washing suspensions | NR (NR) | FT 10× (N2/water bath) | TIANamp Micro DNA Ki | qPCR_Taqman (B1) | 129 | 1 | 10/279 (3.6%) 5 lettuce 2 spinach, 1 pak choi 1 red cabbage 1 rape | [23] |
| carrots (93) slicing cucumbers (109) salads (90) (butterhead lettuce, iceberg lettuce, little gem and lollo lettuce) | 9 farms (292) | 100 g | wash by automatic shaker for 20 min and centrifugation | Tris–glycine beef extract pH 9.5 (230 mL) | BB (6400 rpm for 60 s) | Power-Soil DNA isolation kit | Triplex qPCR_Taqman (B1 + 529RE) | 129 (B1) 163 (529 RE) 157 (IAC) | NR | 28/292 (9.6%) 7 Carrots 13 cucumbers 8 salads | [24] |
| Matrix (Amount) | Spiking Level | Pre-Treatment before DNA Extract | DNA Extraction | Detection (Target Gene) | Limit of Detection (oo)cysts | Reference |
|---|---|---|---|---|---|---|
| faeces (200 mg) | 101–104 | BB | NucleoSpin Soil using Buffer SL2 + Enhancer SLX | PCR (529 RE) | 10 | [25] |
| BB | ZR fecal DNA Kit | 10 | ||||
| FT 3× (10 min at −20 °C/2 min at RT) | phenol/chloroform extraction (in-house) | 100 | ||||
| water or mussels tissues (NR) | 100–103 | FT 5× (liquid N2/70 °C) +BB (glass beads, 30 s at 4200 rpm) + proteinase K 1 h at 56 °C | spin column | PCR (529 RE) | 1 (100%) in water and hemolymph | [26] |
| vortexing (PowerSoil beads max speed for 10 min) + BB (30 s at 4200 rpm) | PowerSoil™ DNA Isolation Kit | 10 (100%) in water and hemolymph, (50%) in dig. glands; 100 (100%) in gills and dig. glands | ||||
| water or mussels tissue homogenate (100 µl) | 100–103 | FT 1× (4 min liquid N2/4 min 100 °C) | spin column | nested PCR (B1) | 100 (100%) with 1, 3 or 6 cycles; 10 (60%) with 1 or 6 cycles; 1 (30%) with 1 cycles | [27] |
| FT 3× (4 min liquid N2/4 min 100 °C | ||||||
| FT 6× (4 min liquid N2/4 min 100 °C |
| Target Gene | Amplicon Size (bp) | LoD (Number of Spiked Oocysts That Provide Positive Amplification) | Primer Pairs | Matrix | Reference |
|---|---|---|---|---|---|
| B1 | 115 | ≥10 oocysts in 250 g of strawberry or 1 lettuce head | B22: 5′-AACGGGCGAGTAGCACCTGAGGAGA-3′ B23: 5′-TGGGTCTACGTCGATGGCATGACAAC-3′ | fresh produce | [16] |
| 10 oocysts | water | [16] | |||
| 10 oocysts/µL spiking level | fresh produce | [18] | |||
| 194 | NR | Oligo1: 5′-GGAACTGCATCCGTTCATGAG-3′ Oligo4: 5′-TCTTTAAAGCGTTCGTGGTC-3′ | soil | [36] | |
| 50 tachyzoites /0.5 g soil | soil | [37] | |||
| NR | soil | [34] | |||
| 100 oocysts | faeces | [38] | |||
| 25 oocysts/30 g soil or ≤1 oocyst/1g soil | soil | [39] | |||
| 529 RE | 529 | ≥100 oocyst in 250 g of strawberry or 1 lettuce head | TOX4: 5′-CTGCAGGGAGGAAGACGAAAGTTG-3′ TOX5: 5′-CTGCAGACAGAGTGCATCTGGATT-3′ | Fresh produce | [16] |
| 10 oocysts | water | [16] | |||
| ≥100 oocysts/ µl spiking level | fresh produce | [18] | |||
| NR | faeces | [40] | |||
| 1 oocyst in water and mussel hemolymph | mussels | [26] | |||
| 100 oocysts in mussel gills and dig. Glands | mussels | [26] | |||
| NR | oysters | [32] | |||
| NR | food | [12] | |||
| 5 tachyzoites /0.5 g soil | soil | [37] | |||
| 10 oocysts | faeces | [38] | |||
| 1–2 oocysts per 200 µL | faeces | [35] | |||
| 450 | 1 oocyst | TOX-8: 5′-CCCAGCTGCGTCTGTCGGGAT-3′ TOX-5: 5′-GACGTCTGTGTCACGTAGACCTAAG-3′ | faeces | [38] | |
| 10 oocysts/200 mg feces | faeces | [25] | |||
| 529 and 450 | NR | TOX4/TOX5 and TOX-8/TOX-5 | faeces | [41] | |
| 134 | NR | TOXO-F: 5′ AGGCGAGGGTGAGGATGA-3′ TOXO-R: 5′-TCGTCTCGTCTGGATCGCAT-3’ | soil | [34] | |
| NR | soil | [36] |
| Type of qPCR | N° of Studies | Target Gene | N° of Studies |
|---|---|---|---|
| MC/HRMC | 9 | B1 | 4 |
| 529RE | 1 | ||
| 18SrDNA | 3 | ||
| B1 and 529RE | 1 | ||
| Taqman | 26 | B1 | 7 |
| 529RE | 16 | ||
| ITS1 | 1 | ||
| 18SrDNA | 1 | ||
| B1 and 18SrDNA | 1 | ||
| FRET | 1 | 529RE | 1 |
| HRM and FRET | 1 | B1 and 529RE | 1 |
| MC and Taqman | 1 | B1 | 1 |
| Total | 38 | 38 |
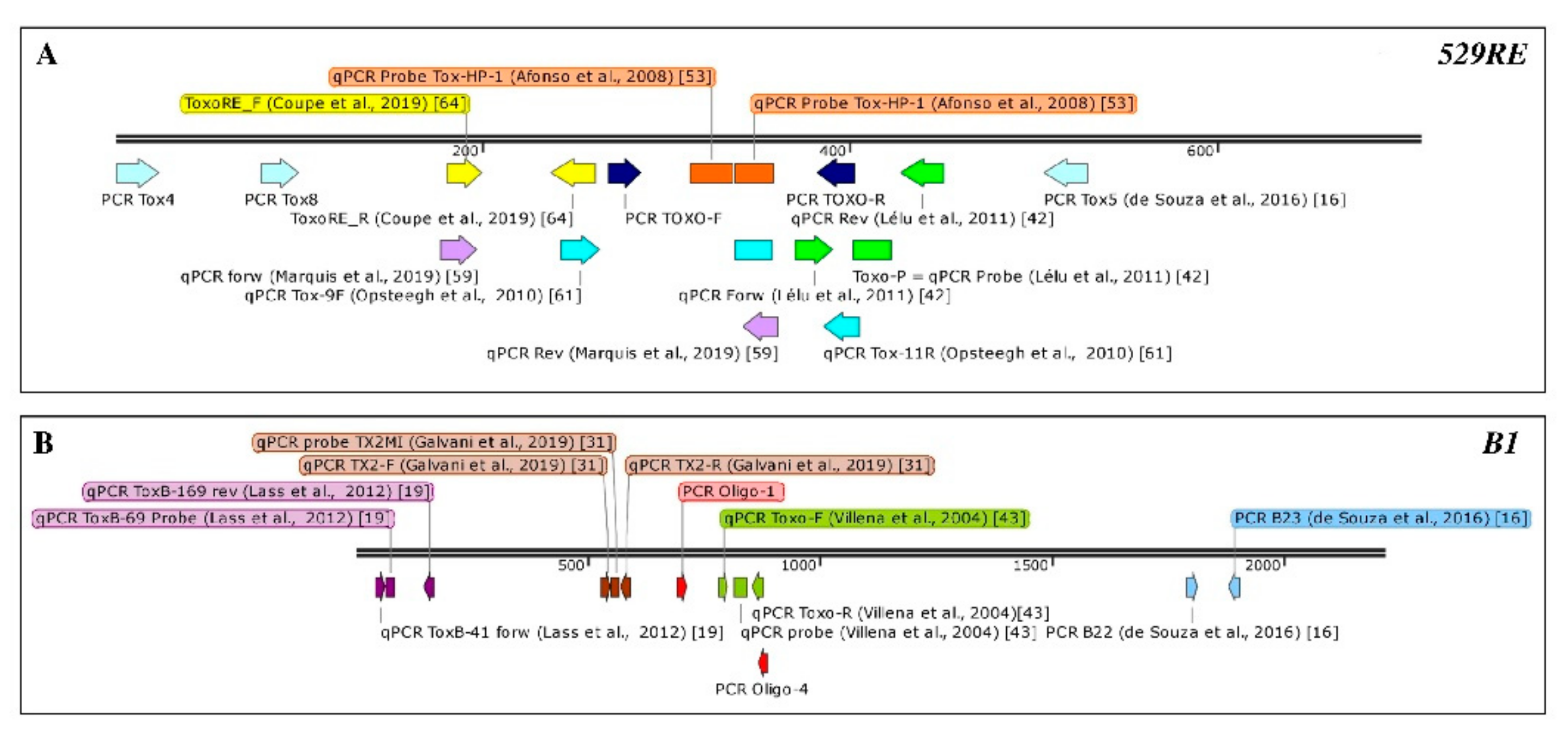
| Target | Amplicon Size (bp) | IAC (Y/N) | Primer Sequence | Analytical Specificity | Analytical Sensitivity | LoD (Oocysts/g) | LoQ | Matrix | Reference |
|---|---|---|---|---|---|---|---|---|---|
| 529 RE | 169 | N | Tox-9F: 5′-AGGAGAGATATCAGGACTGTAG-3′ Tox-11R: 5′-GCGTCGTCTCGTCTAGATCG-3′ Tox-TP1: 5′-6-FAM-CCGGCTTGGCTGCTTTTCCT-BHQ1-3′ | [14] | 90–100% | 5 | NR | mussel hemolymph | [28] |
| 100 oocysts (90%) 10 oocyst (30%) | 10–100 | mussel tissue | |||||||
| Y | Tox-9F; Tox-11R and Tox-TP1 | NR | 100% 2000 oocysts/g | 200 | NR | soil | [44] | ||
| 164 | Y a | Tox-9F; Tox-11R and Tox-TP1 | 1 with 95% CI of 0.69–1.00 | 100% (Crypto or Neospora) | 50 fg/µL | NR | water | [45] | |
| N | Tox-9F; Tox-11R and Tox-TP1 | NR | NR | 1–10 in water and hemolymph | NR | hemolymph and mussel tissue | [26] | ||
| 163 | Y b | Tox-9F; Tox-11R and Tox-TP1 5’-HEX | NR | NR | NR | NR | fresh produce | [24] f | |
| NR | NR | NR | NR | water | |||||
| 162 | N | Tox-9F; Tox-11R and Tox-TP1 5’-Cy5 | In silico 100% | NR | 0.3 | NR | raspberries blueberries | [20] | |
| N | Tox-9F; Tox-11R and Tox-TP1 | 100% (test on parasites, mammalian and plant DNA) | 50–100 oocysts (100%) 25 oocysts (83%) | 0.5 | baby lettuce | [14] | |||
| 81 | Y | [42] Toxo-P 5’-Cy3-ACGCTTTCCTCGTGGTGATGGCG-BHQ2-3’ | NR | NR | 0.1–1 | 5 oocyst/5 g | mussel | [29] | |
| NR | NR | 10–50 | 100 oocysts | haemolymph | |||||
| N | [42] | NR | NR | NR | NR | faeces | [46] | ||
| N | [42]; Toxo-P5′ | NR | NR | NR | NR | mussels | [47] | ||
| NR | NR | NR | NR | water | |||||
| N | [42] | NR | NR | NR | NR | soil | [48] | ||
| N | [42]; Toxo-P5′ | NR | NR | NR | NR | faeces | [49] | ||
| N | [42]; Toxo-P5′ | NR | NR | <1 | NR | basil leaves raspberries | [13] | ||
| N N | [42]; Toxo-P5′ | NR | NR | NR | NR | hemolymph and mussel tissue | [50] | ||
| NR | NR | NR | NR | water | |||||
| N | [42] | NR [51] | Reported in [42] | 10-100 | NR | soil | [52] | ||
| N | FOR_5′-AGAGACACCGGAATGCGATCT-3′ REV_5′-CCCTCTTCTCCACTCTTCAATTCT-3′ Probe 5′-6FAM-ACGCTTTCCTCGTGGTGATGGGG-3´TAMRA | NR | NR | 10-100 | NR | soil | [42] | ||
| N | [42] Probe Tox-HP-1 GAGTCGGAGAGGGAGAAGATGTT-FL Probe Tox-HP-2 Red 640-CCGGCTTGGCTGCTTTTCCTG-Ph | NR | NR | NR | NR | faeces | [53] | ||
| B1 | 129 | Y c | [54] | NR | NR | NR | NR | fresh produce | [23] |
| Y b | [54] ToxB-69p 5′-FAM-ACCGCGAGATGCACCCGCA- BHQ -3′ | NR | NR | NR | NR | fresh produce | [24] f | ||
| NR | NR | NR | NR | water | |||||
| ToxB-41: 5′′-TCGAAGCTGAGATGCTCAAAGTC-3′ ToxB-169 5′′-AATCCACGTCTGGGAAGAACTC-3′ 5′′-FAM-ACCGCGAGATGCACCCGCA TAMRA-3′ | tested by sequencing | 10 molecules of plasmid | 100 | NR | fruits and vegetables | [19] | |||
| 98 | Y d | Toxo-F 5′-TCCCCTCTGCTGGCGAAAAGT-3′ Toxo-R 5′-AGCGTTCGTGGTCAACTATCGATTG-3′ probe V5′-FAM-TCTGTGCAACTTTGGTGTATTCGCAG-3′ TAMRA | NR | NR | NR | NR | water | [43] | |
| N | [43] Toxo-F; Toxo-R and probe V5′ | NR | NR | NR | 1 | water | [55] | ||
| NR | NR | NR | 250 | faeces | |||||
| Y (N.S.) | [43] Toxo-F; Toxo-R and probe V5′ | NR | NR | NR | NR | water | [54] | ||
| 62 | N | TX2-F 5″-CTAGTATCGTGCGGCAATGTG-3′ TX2-R 5′-GGCAGCGTCTCTTCCTCTTTT-3′ TX2M1 5″-(6-FAM)-CCACCTCGCCTCTTGG-(NFQ-MGB)-3′ | NR | NR | 5 genomic copies/µL | 50 genomic copies/µL | water | [31] | |
| 18S rRNA | NR | N | [56] Tox18-213f 5′-CCGGTGGTCCTCAGGTGAT-3′ Tox18-332r 5′-TGCCACGGTAGTCCAATACAGTA-3′ Tox18-249p 5′-FAM-ATCGCGTTGACTTCGGTCTGCGAC-TAMRA-3′ | NR | NR | NR | NR | water | [57] |
| 120 | Y e | Tox18-213f; Tox18-332r and Tox18-249p | 100% (various parasites tested) | NR | 10 molecules | NR | hemolymph and mussels | [58] | |
| ITS1 | NR | N | For 5′-GATTTGCATTCAAGAAGCGTGATAGTA-3′ Rev 5′-AGTTTAGGAAGCAATCTGAAAGCACATC-3′ Probe 5′-/-TET/-CTGCGCTGC/ZEN/TTCCAATATTGG-/-IABkFQ-/-3′ | NR | NR | NR | NR | oysters | [59] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slana, I.; Bier, N.; Bartosova, B.; Marucci, G.; Possenti, A.; Mayer-Scholl, A.; Jokelainen, P.; Lalle, M. Molecular Methods for the Detection of Toxoplasma gondii Oocysts in Fresh Produce: An Extensive Review. Microorganisms 2021, 9, 167. https://doi.org/10.3390/microorganisms9010167
Slana I, Bier N, Bartosova B, Marucci G, Possenti A, Mayer-Scholl A, Jokelainen P, Lalle M. Molecular Methods for the Detection of Toxoplasma gondii Oocysts in Fresh Produce: An Extensive Review. Microorganisms. 2021; 9(1):167. https://doi.org/10.3390/microorganisms9010167
Chicago/Turabian StyleSlana, Iva, Nadja Bier, Barbora Bartosova, Gianluca Marucci, Alessia Possenti, Anne Mayer-Scholl, Pikka Jokelainen, and Marco Lalle. 2021. "Molecular Methods for the Detection of Toxoplasma gondii Oocysts in Fresh Produce: An Extensive Review" Microorganisms 9, no. 1: 167. https://doi.org/10.3390/microorganisms9010167
APA StyleSlana, I., Bier, N., Bartosova, B., Marucci, G., Possenti, A., Mayer-Scholl, A., Jokelainen, P., & Lalle, M. (2021). Molecular Methods for the Detection of Toxoplasma gondii Oocysts in Fresh Produce: An Extensive Review. Microorganisms, 9(1), 167. https://doi.org/10.3390/microorganisms9010167