Genomic Features of MCR-1 and Extended-Spectrum β-Lactamase-Producing Enterobacterales from Retail Raw Chicken in Egypt

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates, Susceptibility Testing
2.2. Phenotypic Confirmation of Resistance Patterns
2.3. Molecular Assays for Other Resistance Determinants
2.4. Mating-Out Assays
2.5. Plasmid Isolation, Plasmid Analysis
2.6. Clonality Evaluation
2.7. Plasmid Sequencing and Bioinformatic Analysis
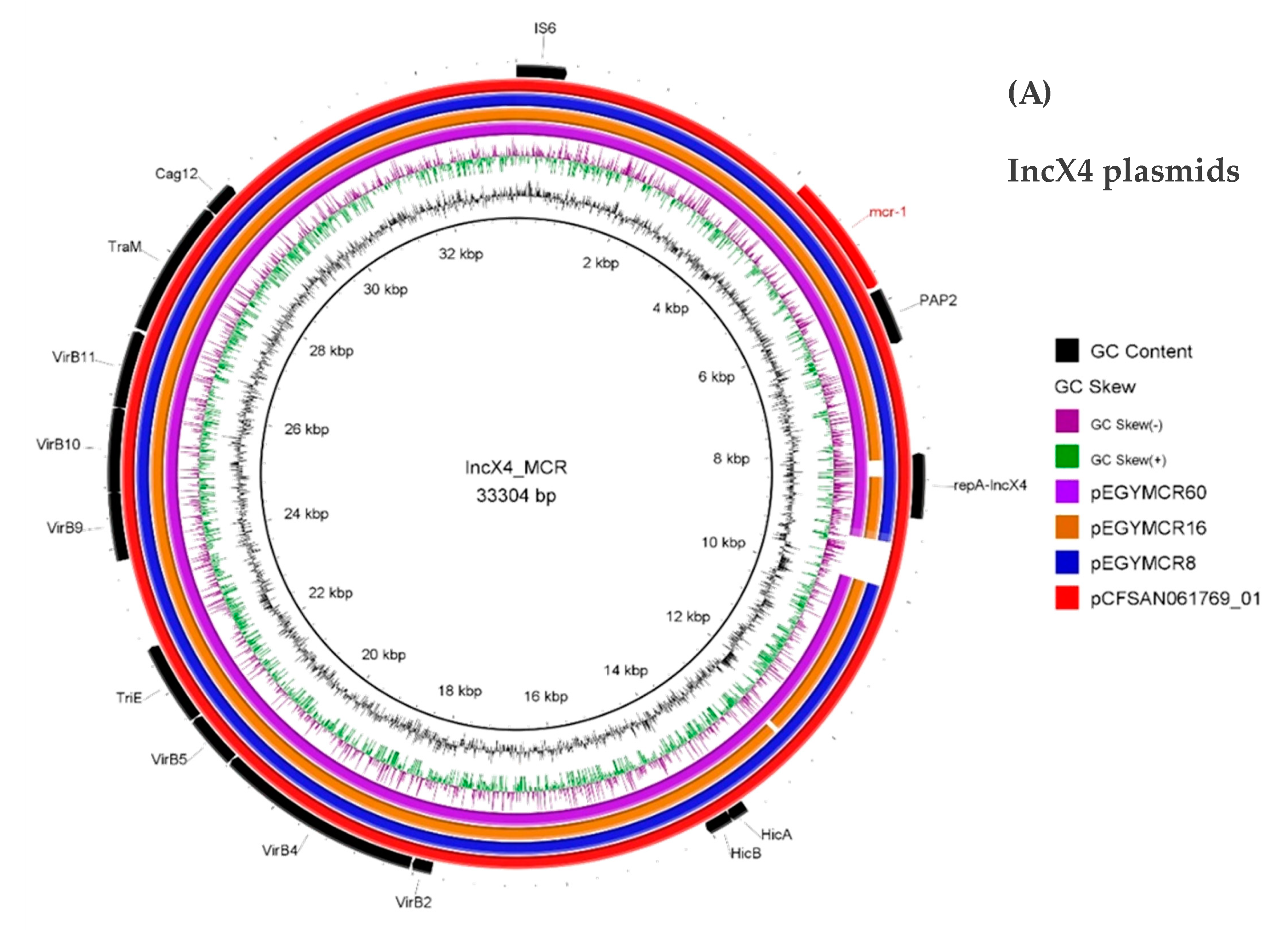
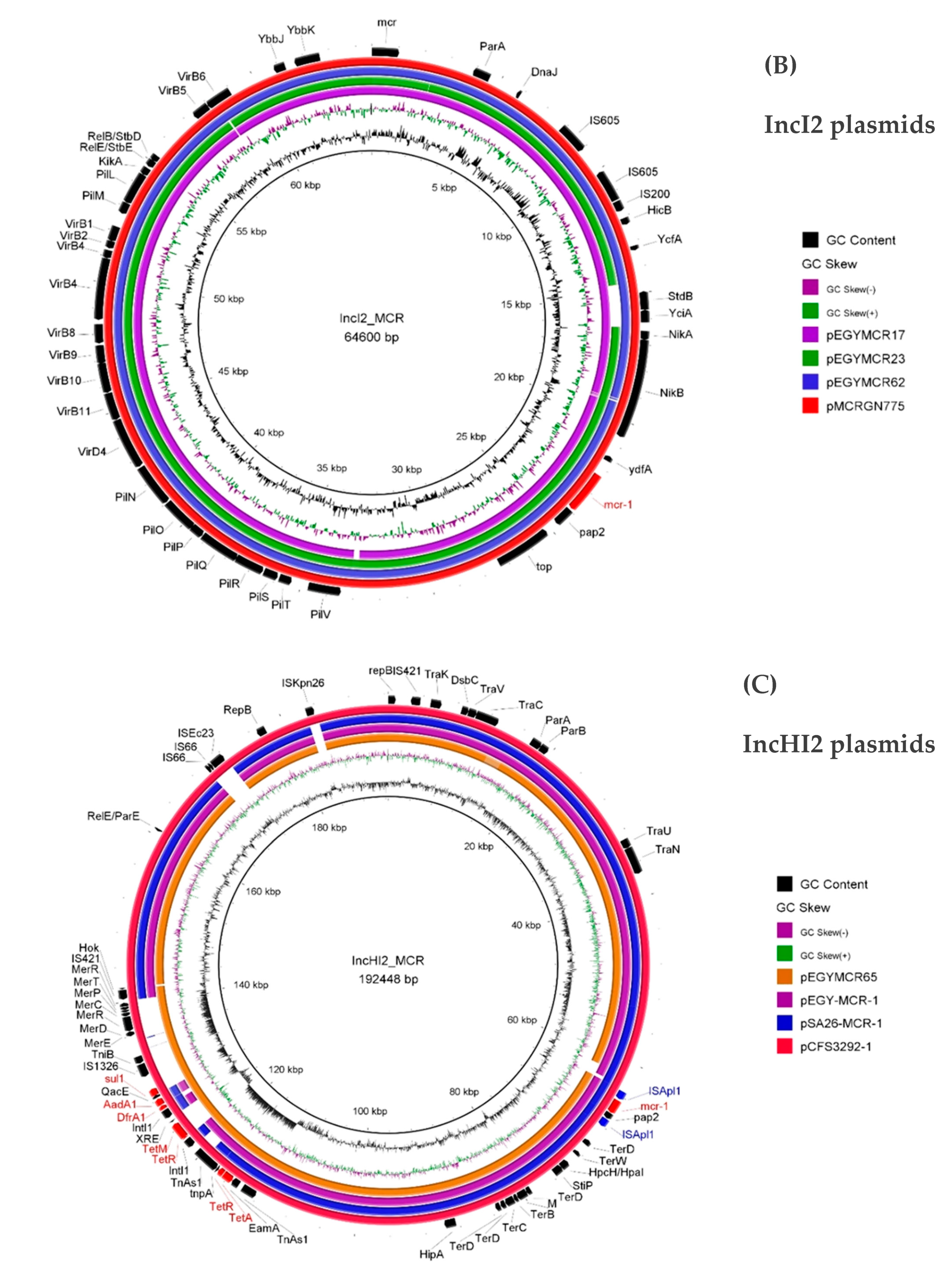
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Poirel, L.; Kieffer, N.; Brink, A.; Coetze, J.; Jayol, A.; Nordmann, P. Genetic features of MCR-1-producing colistin-resistant Escherichia coli isolates in South Africa. Antimicrob. Agents Chemother. 2016, 60, 4394–4397. [Google Scholar] [CrossRef] [Green Version]
- Catry, B.; Cavaleri, M.; Baptiste, K.; Grave, K.; Grein, K.; Holm, A.; Jukes, H.; Liebana, E.; Lopez Navas, A.; Mackay, D.; et al. Use of colistin-containing products within the European Union and European Economic Area (EU/EEA): Development of resistance in animals and possible impact on human and animal health. Int. J. Antimicrob. Agents 2015, 46, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Xavier, B.B.; Lammens, C.; Ruhal, R.; Kumar-Singh, S.; Butaye, P.; Goossens, H.; Malhotra-Kumar, S. Identification of a novel plasmid-mediated colistin-resistance gene, mcr-2, in Escherichia coli, Belgium, June 2016. Eurosurveillance 2016, 21, 1560–7917. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Li, H.; Shen, Y.; Liu, Z.; Wang, S.; Shen, Z.; Zhang, R.; Walsh, T.R.; Shen, J.; Wang, Y. Novel plasmid-mediated colistin resistance gene mcr-3 in Escherichia coli. mBio 2017, 8, e00543-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carattoli, A.; Villa, L.; Feudi, C.; Curcio, L.; Orsini, S.; Luppi, A.; Pezzotti, G.; Magistrali, C.F. Novel plasmid-mediated colistin resistance mcr-4 gene in Salmonella and Escherichia coli, Italy 2013, Spain and Belgium, 2015 to 2016. Eurosurveillance 2017, 22, 30589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borowiak, M.; Fischer, J.; Hammerl, J.A.; Hendriksen, R.S.; Szabo, I.; Malorny, B. Identification of a novel transposon-associated phosphoethanolamine transferase gene, mcr-5, conferring colistin resistance in d-tartrate fermenting Salmonella enterica subsp. enterica serovar Paratyphi B. J. Antimicrob. Chemother. 2017, 72, 3317–3324. [Google Scholar] [CrossRef] [Green Version]
- AbuOun, M.; Stubberfield, E.J.; Duggett, N.A.; Kirchner, M.; Dormer, L.; Nunez-Garcia, J.; Randall, L.P.; Lemma, F.; Crook, D.W.; Teale, C.; et al. mcr-1 and mcr-2 (mcr-6.1) variant genes identified in Moraxella species isolated from pigs in Great Britain from 2014 to 2015. J. Antimicrob. Chemother. 2018, 73, 2904. [Google Scholar] [CrossRef]
- Yang, Y.Q.; Li, Y.X.; Lei, C.W.; Zhang, A.Y.; Wang, H.N. Novel plasmid-mediated colistin resistance gene mcr-7.1 in Klebsiella pneumoniae. J. Antimicrob. Chemother. 2018, 73, 1791–1795. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, Y.; Zhou, Y.; Li, J.; Yin, W.; Wang, S.; Zhang, S.; Shen, J.; Shen, Z.; Wang, Y. Emergence of a novel mobile colistin resistance gene, mcr-8, in NDM-producing Klebsiella pneumoniae. Emerg. Microbes Infect. 2018, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Carroll, L.M.; Gaballa, A.; Guldimann, C.; Sullivan, G.; Henderson, L.O.; Wiedmann, M. Identification of novel mobilized colistin resistance gene mcr-9 in a multidrug-resistant, colistin-susceptible Salmonella enterica serotype Typhimurium isolate. mBio 2019, 10, e00853-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Feng, Y.; Liu, L.; Wei, L.; Kang, M.; Zong, Z. Identification of novel mobile colistin resistance gene mcr-10. Emerg. Microbes Infect. 2020, 9, 508–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xavier, B.B.; Lammens, C.; Butaye, P.; Goossens, H.; Malhotra-Kumar, S. Complete sequence of an IncFII plasmid harbouring the colistin resistance gene mcr-1 isolated from Belgian pig farms. J. Antimicrob. Chemother. 2016, 71, 2342–2344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhi, C.; Lv, L.; Yu, L.F.; Doi, Y.; Liu, J.H. Dissemination of the mcr-1 colistin resistance gene. Lancet. Infect. Dis. 2016, 16, 292–293. [Google Scholar] [CrossRef] [Green Version]
- McGann, P.; Snesrud, E.; Maybank, R.; Corey, B.; Ong, A.C.; Clifford, R.; Hinkle, M.; Whitman, T.; Lesho, E.; Schaecher, K.E. Escherichia coli harboring mcr-1 and blaCTX-M on a novel IncF plasmid: First report of mcr-1 in the United States. Antimicrob. Agents Chemother. 2016, 60, 4420–4421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zurfluh, K.; Klumpp, J.; Nüesch-Inderbinen, M.; Stephan, R. Full-length nucleotide sequences of mcr-1-harboring plasmids isolated from extended-spectrum-β-lactamase-producing Escherichia coli isolates of different origins. Antimicrob. Agents Chemother. 2016, 60, 5589–5591. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Feng, Y.; Liu, F.; Jiang, H.; Qu, Z.; Lei, M.; Wang, J.; Zhang, B.; Hu, Y.; Ding, J.; et al. A phage-like IncY plasmid carrying the mcr-1 gene in Escherichia coli from a pig farm in China. Antimicrob. Agents Chemother. 2017, 61, e02035-16. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; van Dorp, L.; Shaw, L.P.; Bradley, P.; Wang, Q.; Wang, X.; Jin, L.; Zhang, Q.; Liu, Y.; Rieux, A. The global distribution and spread of the mobilized colistin resistance gene mcr-1. Nat. Commun. 2018, 9, 1179. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial activity, susceptibility testing, and resistance mechanisms encoded by plasmids or chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef] [Green Version]
- Irrgang, A.; Roschanski, N.; Tenhagen, B.A.; Grobbel, M.; Skladnikiewicz-Ziemer, T.; Thomas, K.; Roesler, U.; Käsbohrer, A. Prevalence of mcr-1 in E. coli from livestock and food in Germany, 2010–2015. PLoS ONE 2016, 11, e0159863. [Google Scholar] [CrossRef]
- Hassen, B.; Abbassi, M.S.; Ruiz-Ripa, L.; Mama, O.M.; Hassen, A.; Torres, C.; Hammami, S. High prevalence of mcr-1 encoding colistin resistance and first identification of blaCTX-M-55 in ESBL/CMY-2-producing Escherichia coli isolated from chicken faeces and retail meat in Tunisia. Int. J. Food Microbiol. 2020, 318, 108478. [Google Scholar] [CrossRef] [PubMed]
- Dhaouadi, S.; Soufi, L.; Hamza, A.; Fedida, D.; Zied, C.; Awadhi, E.; Mtibaa, M.; Hassen, B.; Cherif, A.; Torres, C. Co-occurrence of mcr-1 mediated colistin resistance and β-lactamase-encoding genes in multidrug-resistant Escherichia coli from broiler chickens with colibacillosis in Tunisia. J. Glob. Antimicrob. Resist. 2020, 22, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Sadek, M.; Poirel, L.; Nordmann, P.; Nariya, H.; Shimamoto, T.; Shimamoto, T. Draft genome sequence of an mcr-1/IncI2-carrying multidrug-resistant Escherichia coli B1:ST101 isolated from meat and meat products in Egypt. J. Glob. Antimicrob. Resist. 2020, 20, 41–42. [Google Scholar] [CrossRef] [PubMed]
- Hammad, A.M.; Hoffmann, M.; Gonzalez-Escalona, N.; Abbas, N.H.; Yao, K.; Koenig, S.; Allué-Guardia, A.; Eppinger, M. Genomic features of colistin resistant Escherichia coli ST69 strain harboring mcr-1 on IncHI2 plasmid from raw milk cheese in Egypt. Infect. Genet. Evol. 2019, 73, 126–131. [Google Scholar] [CrossRef]
- Hassen, B.; Saloua, B.; Abbassi, M.S.; Ruiz-Ripa, L.; Mama, O.M.; Hassen, A.; Hammami, S.; Torres, C. mcr-1 encoding colistin resistance in CTX-M-1/CTX-M-15- producing Escherichia coli isolates of bovine and caprine origins in Tunisia. First report of CTX-M-15-ST394/D E. coli from goats. Comp. Immunol. Microbiol. Infect. Dis. 2019, 67, 101366. [Google Scholar] [CrossRef]
- Touati, A.; Mairi, A. Plasmid-determined colistin resistance in the North African countries: A systematic review. Microb. Drug Resist. 2020. [Google Scholar] [CrossRef]
- Kieffer, N.; Royer, G.; Decousser, J.W.; Bourrel, A.S.; Palmieri, M.; Ortiz De La Rosa, J.M.; Jacquier, H.; Denamur, E.; Nordmann, P.; Poirel, L. mcr-9, an inducible gene encoding an acquired phosphoethanolamine transferase in Escherichia coli, and its origin. Antimicrob. Agents Chemother. 2019, 63, e00965-19. [Google Scholar] [CrossRef] [Green Version]
- Nordmann, P.; Jayol, A.; Poirel, L. Rapid detection of polymyxin resistance in Enterobacteriaceae. Emerg. Infect. Dis. 2016, 22, 1038–1043. [Google Scholar] [CrossRef] [Green Version]
- Nordmann, P.; Dortet, L.; Poirel, L. Rapid detection of extended-spectrum-β-lactamase-producing Enterobacteriaceae. J. Clin. Microbiol. 2012, 50, 3016–3022. [Google Scholar] [CrossRef] [Green Version]
- Nordmann, P.; Poirel, L.; Mueller, L. Rapid detection of fosfomycin resistance in Escherichia coli. J. Clin. Microbiol. 2019, 57, e01531-18. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Dortet, L.; Bernabeu, S.; Nordmann, P. Genetic features of blaNDM-1-positive Enterobacteriaceae. Antimicrob. Agents Chemother. 2011, 55, 5403–5407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirel, L.; Vuillemin, X.; Kieffer, N.; Mueller, L.; Descombes, M.C.; Nordmann, P. Identification of FosA8, a plasmid-encoded fosfomycin resistance determinant from Escherichia coli, and its origin in Leclercia adecarboxylata. Antimicrob. Agents Chemother. 2019, 63, e01403-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirel, L.; Savov, E.; Nazli, A.; Trifonova, A.; Todorova, I.; Gergova, I.; Nordmann, P. Outbreak caused by NDM-1- and RmtB-producing Escherichia coli in Bulgaria. Antimicrob. Agents Chemother. 2014, 58, 2472–2474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threlfall, E.J. Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods. 2005, 63, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Kieffer, N.; Nordmann, P.; Aires-de-Sousa, M.; Poirel, L. High prevalence of carbapenemase-producing enterobacteriaceae among hospitalized children in Luanda, Angola. Antimicrob. Agents Chemother. 2016, 60, 6189–6192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zurfluh, K.; Nüesch-Inderbinen, M.; Klumpp, J.; Poirel, L.; Nordmann, P.; Stephan, R. Key features of mcr-1-bearing plasmids from Escherichia coli isolated from humans and food. Antimicrob. Resist. Infect. Control. 2017, 6, 91. [Google Scholar] [CrossRef]
- Kieser, T. Factors affecting the isolation of CCC DNA from Streptomyces lividans and Escherichia coli. Plasmid. 1984, 12, 19–36. [Google Scholar] [CrossRef]
- Peirano, G.; van der Bij, A.K.; Freeman, J.L.; Poirel, L.; Nordmann, P.; Costello, M.; Tchesnokova, V.L.; Pitout, J.D. Characteristics of Escherichia coli sequence type 131 isolates that produce extended-spectrum β-lactamases: Global distribution of the H30-Rx sublineage. Antimicrob. Agents Chemother. 2014, 58, 3762–3767. [Google Scholar] [CrossRef] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genomics 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Alikhan, N.F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genomics 2011, 12, 402. [Google Scholar] [CrossRef] [Green Version]
- Lima Barbieri, N.; Nielsen, D.W.; Wannemuehler, Y.; Cavender, T.; Hussein, A.; Yan, S.G.; Nolan, L.K.; Logue, C.M. mcr-1 identified in avian pathogenic Escherichia coli (APEC). PLoS ONE 2017, 12, e0172997. [Google Scholar] [CrossRef] [Green Version]
- Dandachi, I.; Chaddad, A.; Hanna, J.; Matta, J.; Daoud, Z. Understanding the epidemiology of multi-drug resistant Gram-negative bacilli in the middle east using a one health approach. Front. Microbiol. 2019, 10, 1941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zafer, M.M.; El-Mahallawy, H.A.; Abdulhak, A.; Amin, M.A.; Al-Agamy, M.H.; Radwan, H.H. Emergence of colistin resistance in multidrug-resistant Klebsiella pneumoniae and Escherichia coli strains isolated from cancer patients. Ann. Clin. Microbiol. Antimicrob. 2019, 18, 40. [Google Scholar] [CrossRef]
- Khalifa, H.O.; Ahmed, A.M.; Oreiby, A.F.; Eid, A.M.; Shimamoto, T.; Shimamoto, T. Characterisation of the plasmid-mediated colistin resistance gene mcr-1 in Escherichia coli isolated from animals in Egypt. Int. J. Antimicrob. Agents 2016, 47, 413–414. [Google Scholar] [CrossRef] [PubMed]
- Moawad, A.A.; Hotzel, H.; Neubauer, H.; Ehricht, R.; Monecke, S.; Tomaso, H.; Hafez, H.M.; Roesler, U.; El-Adawy, H. Antimicrobial resistance in Enterobacteriaceae from healthy broilers in Egypt: Emergence of colistin-resistant and extended-spectrum β-lactamase-producing Escherichia coli. Gut Pathog. 2018, 10, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Wang, Y.; Shi, X.; Wang, S.; Ren, H.; Shen, Z.; Wang, Y.; Lin, J.; Wang, S. Rapid rise of the ESBL and mcr-1 genes in Escherichia coli of chicken origin in China, 2008–2014. Emerg. Microbes. Infect. 2018, 7, 30. [Google Scholar] [CrossRef] [Green Version]
- Carfora, V.; Alba, P.; Leekitcharoenphon, P.; Ballarò, D.; Cordaro, G.; Di Matteo, P.; Donati, V.; Ianzano, A.; Iurescia, M.; Stravino, F. Colistin resistance mediated by mcr-1 in ESBL-producing, multidrug resistant Salmonella Infantis in broiler chicken industry, Italy (2016–2017). Front. Microbiol. 2018, 9, 1880. [Google Scholar] [CrossRef]
- Kieffer, N.; Poirel, L.; Clerc, O.; Lienhard, R.; Nordmann, P. Co-production of MCR-1 and extended-spectrum β-lactamase in Escherichia coli recovered from urinary tract infections in Switzerland. Infection 2018, 46, 143–144. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, J.; Wang, X.; Bai, X.; Ma, J.; Dang, R.; Xiong, Y.; Fanning, S.; Bai, L.; Yang, Z. Characterization of five Escherichia coli isolates co-expressing ESBL and MCR-1 resistance mechanisms from different origins in China. Front. Microbiol. 2019, 10, 1994. [Google Scholar] [CrossRef] [Green Version]
- Fournier, C.; Aires-de-Sousa, M.; Nordmann, P.; Poirel, L. Occurrence of CTX-M-15- and MCR-1-producing Enterobacterales in pigs in Portugal: Evidence of direct links with antibiotic selective pressure. Int. J. Antimicrob. Agents. 2020, 55, 105802. [Google Scholar] [CrossRef]
- Liu, G.; Ali, T.; Gao, J.; Ur Rahman, S.; Yu, D.; Barkema, H.W.; Huo, W.; Xu, S.; Shi, Y.; Kastelic, J.P. Co-occurrence of plasmid-mediated colistin resistance (mcr-1) and extended-spectrum β-lactamase encoding genes in Escherichia coli from bovine mastitic milk in China. Microb. Drug Resist. 2020, 26, 685–696. [Google Scholar] [CrossRef] [PubMed]
- Grami, R.; Mansour, W.; Mehri, W.; Bouallègue, O.; Boujaâfar, N.; Madec, J.Y.; Haenni, M. Impact of food animal trade on the spread of mcr-1-mediated colistin resistance, Tunisia, July 2015. Eurosurveillance 2016, 21, 30144. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Li, X.P.; Yang, R.S.; Fang, L.X.; Huo, W.; Li, S.M.; Jiang, P.; Liao, X.P.; Liu, Y.H. Complete nucleotide sequence of an IncI2 plasmid coharboring blaCTX-M-55 and mcr-1. Antimicrob. Agents Chemother. 2016, 60, 5014–5017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Fang, L.X.; Wu, Z.; Deng, H.; Yang, R.S.; Li, X.P.; Li, S.M.; Liao, X.P.; Feng, Y.; Liu, Y.H. Genetic analysis of the IncX4 plasmids: Implications for a unique pattern in the mcr-1 acquisition. Sci. Rep. 2017, 7, 424. [Google Scholar] [CrossRef] [PubMed]
- Saidani, M.; Messadi, L.; Chaouechi, A.; Tabib, I.; Saras, E.; Soudani, A.; Daaloul-Jedidi, M.; Mamlouk, A.; Ben Chehida, F.; Chakroun, C.; et al. High genetic diversity of Enterobacteriaceae clones and plasmids disseminating resistance to extended-spectrum cephalosporins and colistin in healthy chicken in Tunisia. Microb. Drug Resist. 2019, 25, 1507–1513. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, G.; Wachino, J.; Sato, N.; Kimura, K.; Yamada, K.; Jin, W.; Shibayama, K.; Yagi, T.; Kawamura, K.; Arakawa, Y. Practical agar-based disk potentiation test for detection of fosfomycin-nonsusceptible Escherichia coli clinical isolates producing glutathione S-transferases. J. Clin. Microbiol. 2014, 52, 3175–3179. [Google Scholar] [CrossRef] [Green Version]
- Mowlaboccus, S.; Daley, D.; Pang, S.; Gottlieb, T.; Merlino, J.; Nimmo, G.R.; George, N.; Korman, T.M.; Streitberg, R.; Robson, J.; et al. Identification and characterization of fosfomycin resistance in Escherichia coli urinary tract infection isolates from Australia. Int. J. Antimicrob. Agents 2020, 56, 106121. [Google Scholar] [CrossRef]
- Hou, J.; Yang, X.; Zeng, Z.; Lv, L.; Yang, T.; Lin, D.; Liu, J.H. Detection of the plasmid-encoded fosfomycin resistance gene fosA3 in Escherichia coli of food-animal origin. J. Antimicrob. Chemother. 2013, 68, 766–770. [Google Scholar] [CrossRef]
- Wang, X.M.; Dong, Z.; Schwarz, S.; Zhu, Y.; Hua, X.; Zhang, Y.; Liu, S.; Zhang, W.J. Plasmids of diverse inc groups disseminate the fosfomycin resistance gene fosA3 among Escherichia coli isolates from pigs, chickens, and dairy cows in Northeast China. Antimicrob. Agents Chemother. 2017, 61, e00859-17. [Google Scholar] [CrossRef] [Green Version]
- Lupo, A.; Saras, E.; Madec, J.Y.; Haenni, M. Emergence of blaCTX-M-55 associated with fosA, rmtB and mcr gene variants in Escherichia coli from various animal species in France. J. Antimicrob. Chemother. 2018, 73, 867–872. [Google Scholar] [CrossRef]
- Palmeira, J.D.; Ferreira, H.; Madec, J.Y.; Haenni, M. Pandemic Escherichia coli ST648 isolate harbouring fosA3 and blaCTX-M-8 on an IncI1/ST113 plasmid: A new successful combination for the spread of fosfomycin resistance? J. Glob. Antimicrob. Resist. 2018, 15, 254–255. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.Y.; Lu, P.L.; Tseng, S.P. Update on fosfomycin-modified genes in Enterobacteriaceae. J. Microbiol. Immunol. Infect. 2019, 52, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Falgenhauer, L.; Waezsada, S.E.; Yao, Y.; Imirzalioglu, C.; Käsbohrer, A.; Roesler, U.; Michael, G.B.; Schwarz, S.; Werner, G.; Kreienbrock, L.; et al. Colistin resistance gene mcr-1 in extended-spectrum β-lactamase-producing and carbapenemase-producing Gram-negative bacteria in Germany. Lancet. Infect. Dis. 2016, 16, 282–283. [Google Scholar] [CrossRef] [Green Version]
- Veldman, K.; van Essen-Zandbergen, A.; Rapallini, M.; Wit, B.; Heymans, R.; van Pelt, W.; Mevius, D. Location of colistin resistance gene mcr-1 in Enterobacteriaceae from livestock and meat. J. Antimicrob. Chemother. 2016, 71, 2340–2342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Xie, M.; Lv, J.; Wai-Chi Chan, E.; Chen, S. Complete genetic analysis of plasmids carrying mcr-1 and other resistance genes in an Escherichia coli isolate of animal origin. J. Antimicrob. Chemother. 2017, 72, 696–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.T.; Song, F.J.; Zou, M.; Zhang, Q.D.; Shan, H. High incidence of Escherichia coli strains coharboring mcr-1 and blaNDM from chickens. Antimicrob. Agents Chemother. 2017, 61, e02347-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matamoros, S.; van Hattem, J.M.; Arcilla, M.S.; Willemse, N.; Melles, D.C.; Penders, J.; Vinh, T.N.; Thi Hoa, N.; Bootsma, M.; van Genderen, P.J.; et al. Global phylogenetic analysis of Escherichia coli and plasmids carrying the mcr-1 gene indicates bacterial diversity but plasmid restriction. Sci. Rep. 2017, 7, 15364. [Google Scholar] [CrossRef] [Green Version]
- Roschanski, N.; Falgenhauer, L.; Grobbel, M.; Guenther, S.; Kreienbrock, L.; Imirzalioglu, C.; Roesler, U. Retrospective survey of mcr-1 and mcr-2 in German pig-fattening farms, 2011–2012. Int. J. Antimicrob. Agents 2017, 50, 266–271. [Google Scholar] [CrossRef]
- Tijet, N.; Faccone, D.; Rapoport, M.; Seah, C.; Pasterán, F.; Ceriana, P.; Albornoz, E.; Corso, A.; Petroni, A.; Melano, R.G. Molecular characteristics of mcr-1-carrying plasmids and new mcr-1 variant recovered from polyclonal clinical Escherichia coli from Argentina and Canada. PLoS ONE 2017, 12, e0180347. [Google Scholar] [CrossRef]
- Li, R.; Xie, M.; Zhang, J.; Yang, Z.; Liu, L.; Liu, X.; Zheng, Z.; Chan, E.W.; Chen, S. Genetic characterization of mcr-1-bearing plasmids to depict molecular mechanisms underlying dissemination of the colistin resistance determinant. J. Antimicrob. Chemother. 2017, 72, 393–401. [Google Scholar] [CrossRef] [Green Version]
- Snesrud, E.; Ong, A.C.; Corey, B.; Kwak, Y.I.; Clifford, R.; Gleeson, T.; Wood, S.; Whitman, T.J.; Lesho, E.P.; Hinkle, M.; et al. Analysis of serial isolates of mcr-1-positive Escherichia coli reveals a highly active ISApl1 transposon. Antimicrob. Agents Chemother. 2017, 61, e00056-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, A.; Yang, Y.; Miao, M.; Chavda, K.D.; Mediavilla, J.R.; Xie, X.; Feng, P.; Tang, Y.W.; Kreiswirth, B.N.; Chen, L.; et al. Complete sequences of mcr-1-harboring plasmids from extended-spectrum-β-lactamase- and carbapenemase-producing Enterobacteriaceae. Antimicrob. Agents Chemother. 2016, 60, 4351–4354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Li, R.; Zheng, Z.; Chen, K.; Xie, M.; Chan, E.W.; Geng, S.; Chen, S. Molecular characterization of Escherichia coli isolates carrying mcr-1, fosA3, and extended-spectrum-β-lactamase genes from food samples in China. Antimicrob. Agents Chemother. 2017, 61, e00064-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Strains | Species | Origin (Source) | ST (CC) | Resistance Genes | MIC of Colistin (µg/mL) | Rapid Polymyxin NP Test | Rapid ESBL NP Test | Resistance Profile | mcr-1-Harboring Plasmid | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Inc Group (Kb) | MIC Colistin (mg/L) | Co-Resistance Markers | |||||||||
| EC10.2 | E. coli | Chicken carcass | ST101 (CC101) | mcr-1, blaTEM-1 | 4 | + | - | AMX, TIC, SUL, NAL, CIP, CHL, TET, SXT, GMI, TMN | IncI2 (≈66) | 4 | none |
| EC16.1 | E. coli | Chicken carcass | ST1196 | mcr-1, blaTEM-1, blaCTX-M-9 | 4 | + | + | AMX, TIC, PIL, CTX, CEF, FEP, TET, TMN, SXT, KMN, CIP, GMI, NAL, SUL, CHL | IncX4 (31.8) | 4 | none |
| EC20.1 | E. coli | Chicken carcass | ST1196 | mcr-1, blaTEM-1, blaCTX-M-9 | 8 | + | + | AMX, TIC, PIL, FEP, CTX, CEF, NAL, CIP, TET, SUL, CHL, KMN, GMI, TMN, SXT | IncX4 (≈32) | 4 | none |
| EC8.1 | E. coli | Chicken carcass | ST371 (CC350) | mcr-1, fosA4, blaTEM-1 | 4 | + | - | AMX, TIC, PIL, SUL, NAL, KMN, TET, SXT, FOS | IncX4 (32.1) | 4 | none |
| EC23.1 | E. coli | Chicken carcass | ST398 (CC398) | mcr-1, blaTEM-1, blaSHV-12 | 4 | + | + | AMX, TIC, PIL, CEF, CZD, ATM, CTX, NAL, CIP, TET, SUL, CHL, SXT, KMN, GMI, TMN | IncI2 (67.7) | 4 | none |
| EC49.2 | E. coli | Chicken carcass | ST1125 | mcr-1, blaTEM-1 | 4 | + | - | AMX, TIC, PIL, SUL, CHL, TET | IncI2 (≈66) | 4 | none |
| EC62.2 | E. coli | Chicken carcass | ST5687 | mcr-1, blaCTX-M-15 | 4 | + | + | AMX, TIC, PIL, CEF, FEP, CZD, ATM, CTX, NAL, CIP, TET, SUL, CHL, KMN, GMI, SXT | IncI2 (64.5) | 4 | none |
| EC59.1 | E. coli | Chicken carcass | ST5687 | mcr-1, blaCTX-M-15 | 4 | + | + | AMX, TIC, PIL, FEP, CEF, CZD, ATM, CTX, NAL, CIP, TET, SUL, CHL, KMN, GMI, SXT | IncI2 (≈66) | 4 | none |
| EC65.2 | E. coli | Chicken carcass | ST1011 | mcr-1, blaTEM-1, blaCTX-M-14 | 16 | + | + | AMX, TIC, PIL, CTX, FEP, CEF, NAL, CIP, TET, SUL, CHL, KMN, GMI, SXT. | IncHI2 (195.8) | 8 | TET, SUL, CHL |
| EC53.2 | E. coli | Chicken carcass | ST156 (CC156) | mcr-1, blaTEM-1, blaCTX-M-14 | 4 | + | + | AMX, TIC, PIL, CEF, CTX, NAL, CIP, TET, SUL, CHL, KMN, GMI, TMN | IncX4 (≈32) | 4 | none |
| EC60.2 | E. coli | Chicken carcass | ST156 (CC156) | mcr-1, blaTEM-1, blaCTX-M-14 | 4 | + | + | AMX, TIC, PIL, CEF, CTX, NAL, CIP, TET, SUL, CHL, KMN, GMI, TMN | IncX4 (32.1) | 4 | none |
| EC54.2 | E. coli | Chicken carcass | ST156 (CC156) | mcr-1, blaTEM-1, blaCTX-M-14 | 8 | + | + | AMX, PIL, TIC, CEF, CTX, NAL, CIP, TET, SUL, CHL, KMN, TMN, GMI | IncX4 (≈32) | 4 | none |
| EC56.2 | E. coli | Chicken carcass | ST986 | mcr-1 | 8 | + | - | TET, SUL, CHL, KMN | IncHI2 (>154) | 8 | TET, SUL, CHL, KMN |
| EC57.2 | E. coli | Chicken carcass | ST986 | mcr-1 | 8 | + | - | TET, SUL, CHL, KMN | IncHI2 (>154) | 4 | TET, SUL, CHL, KMN |
| EC52.2 | E. coli | Chicken carcass | Unknown | mcr-1, blaTEM-1 | 8 | + | - | AMX, TIC, SUL, CHL, SXT, TET | IncI2 (≈66) | 8 | none |
| CF12.2 | C. freundii | Chicken carcass | ND | mcr-1, blaTEM-1 | 8 | + | - | AMX, TIC, FOX, CEF, AMC, CHL, SUL | IncI2 (≈66) | 8 | none |
| EC17.1 | E. coli | Chicken carcass | ST373 (CC168) | mcr-1, blaTEM-1 | 8 | + | - | AMX, TIC, SUL, CHL, TET, PIL, SXT, NAL | IncI2 (64.2) | 4 | none |
| EC18.2 | E. coli | Chicken carcass | Unknown | mcr-1, blaTEM-1 | 8 | + | - | AMX, TIC, PIL, AMC, SUL, CHL, NAL, TET, FOX | IncI2 (≈66) | 4 | none |
| EC15.2 | E. coli | Chicken carcass | ST373 (CC168) | mcr-1, blaTEM-1 | 8 | + | - | AMX, TIC, SUL, CHL, TET | IncI2 (≈66) | 8 | none |
| EC13.1 | E. coli | Chicken carcass | ST373 (CC168) | mcr-1, blaTEM-1 | 8 | + | - | AMX, TIC, PIL, TET, SUL, NAL, CHL | IncI2 (≈66) | 4 | none |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadek, M.; Ortiz de la Rosa, J.M.; Abdelfattah Maky, M.; Korashe Dandrawy, M.; Nordmann, P.; Poirel, L. Genomic Features of MCR-1 and Extended-Spectrum β-Lactamase-Producing Enterobacterales from Retail Raw Chicken in Egypt. Microorganisms 2021, 9, 195. https://doi.org/10.3390/microorganisms9010195
Sadek M, Ortiz de la Rosa JM, Abdelfattah Maky M, Korashe Dandrawy M, Nordmann P, Poirel L. Genomic Features of MCR-1 and Extended-Spectrum β-Lactamase-Producing Enterobacterales from Retail Raw Chicken in Egypt. Microorganisms. 2021; 9(1):195. https://doi.org/10.3390/microorganisms9010195
Chicago/Turabian StyleSadek, Mustafa, José Manuel Ortiz de la Rosa, Mohamed Abdelfattah Maky, Mohamed Korashe Dandrawy, Patrice Nordmann, and Laurent Poirel. 2021. "Genomic Features of MCR-1 and Extended-Spectrum β-Lactamase-Producing Enterobacterales from Retail Raw Chicken in Egypt" Microorganisms 9, no. 1: 195. https://doi.org/10.3390/microorganisms9010195
APA StyleSadek, M., Ortiz de la Rosa, J. M., Abdelfattah Maky, M., Korashe Dandrawy, M., Nordmann, P., & Poirel, L. (2021). Genomic Features of MCR-1 and Extended-Spectrum β-Lactamase-Producing Enterobacterales from Retail Raw Chicken in Egypt. Microorganisms, 9(1), 195. https://doi.org/10.3390/microorganisms9010195