Improving the Cathodic Biofilm Growth Capabilities of Kyrpidia spormannii EA-1 by Undirected Mutagenesis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Liquid Cultivation of Kyrpidia spormannii
2.2. Bioelectrochemical Cultivation
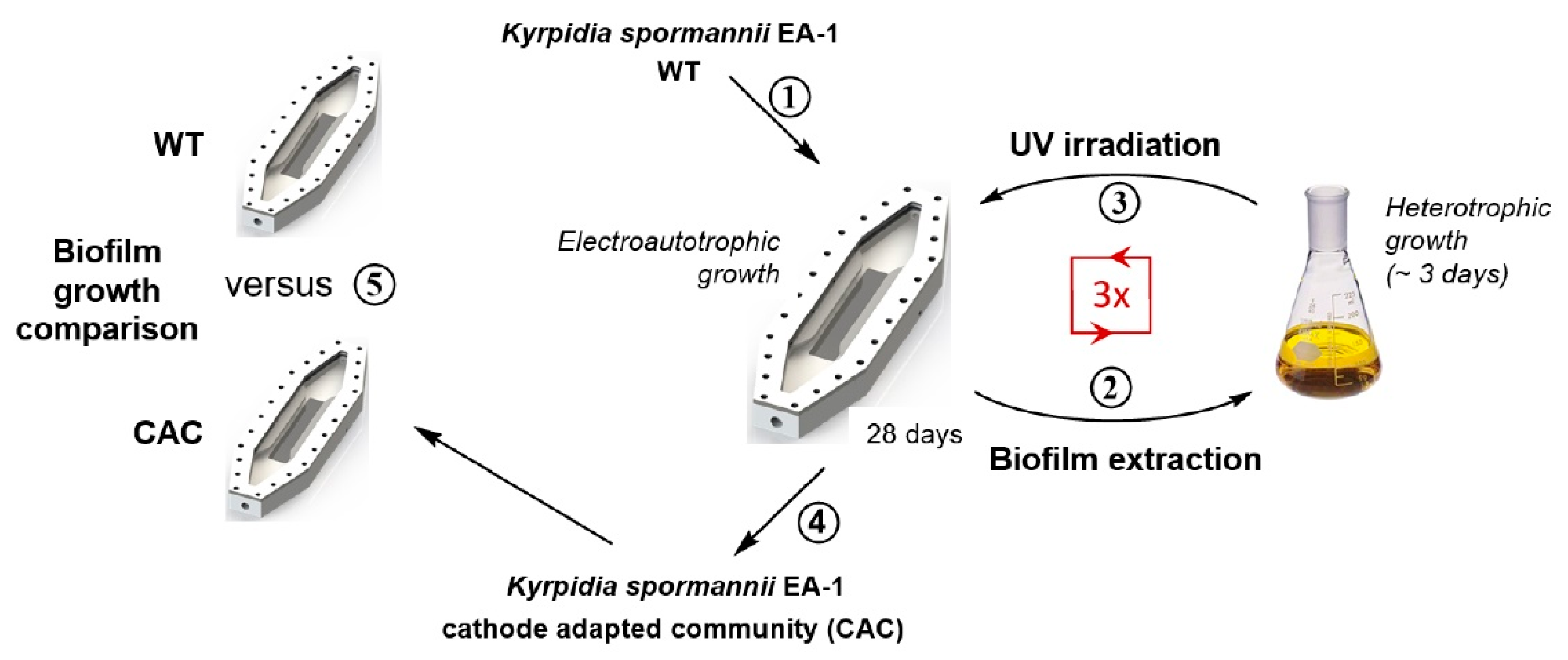
2.3. UV-Assisted Cathodic Adaptation
2.4. Genome Sequencing and Variant Detection
2.5. Ion Exchange Chromatography (IC)
2.6. Blast- and Protein Domain Analyses
3. Results and Discussion
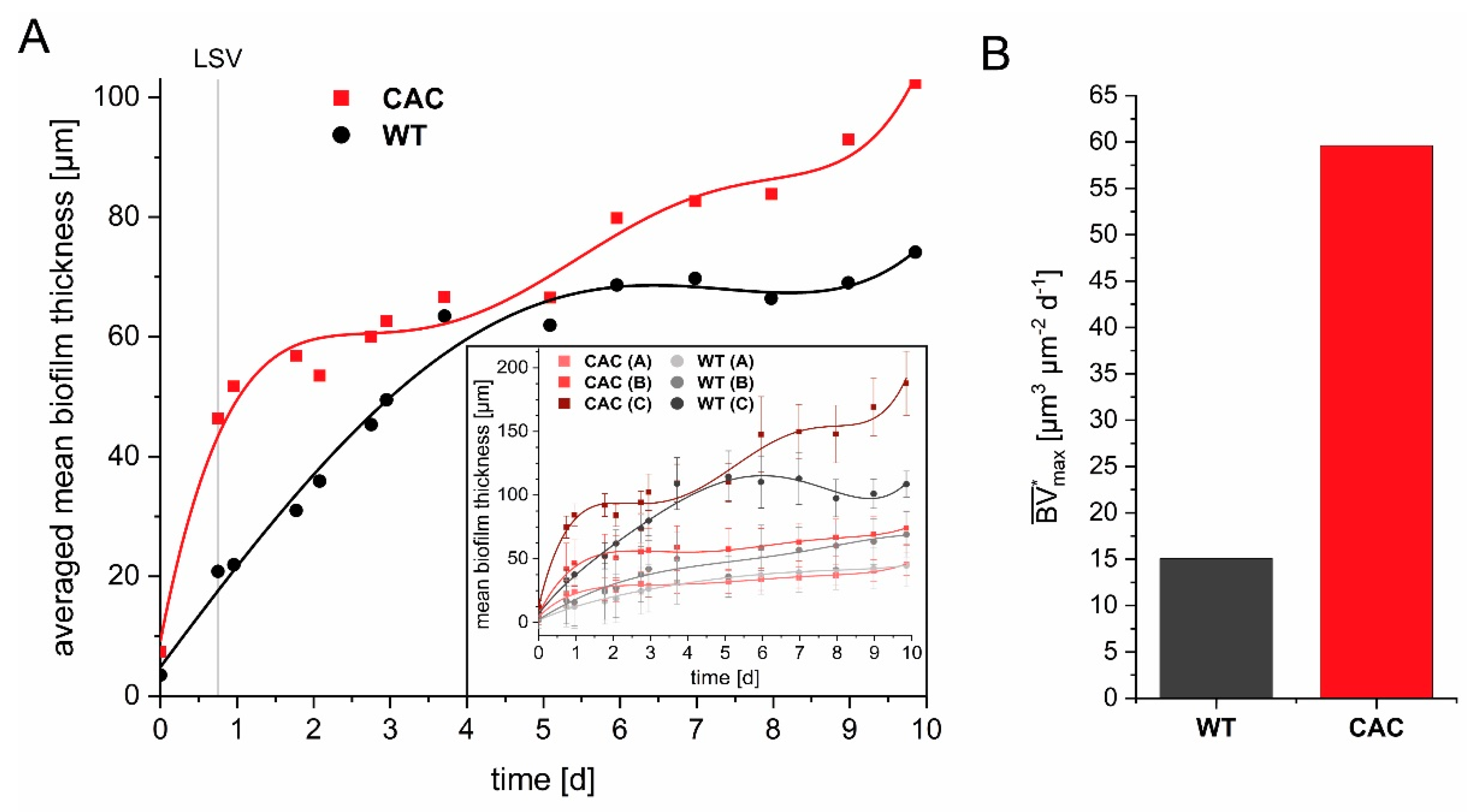
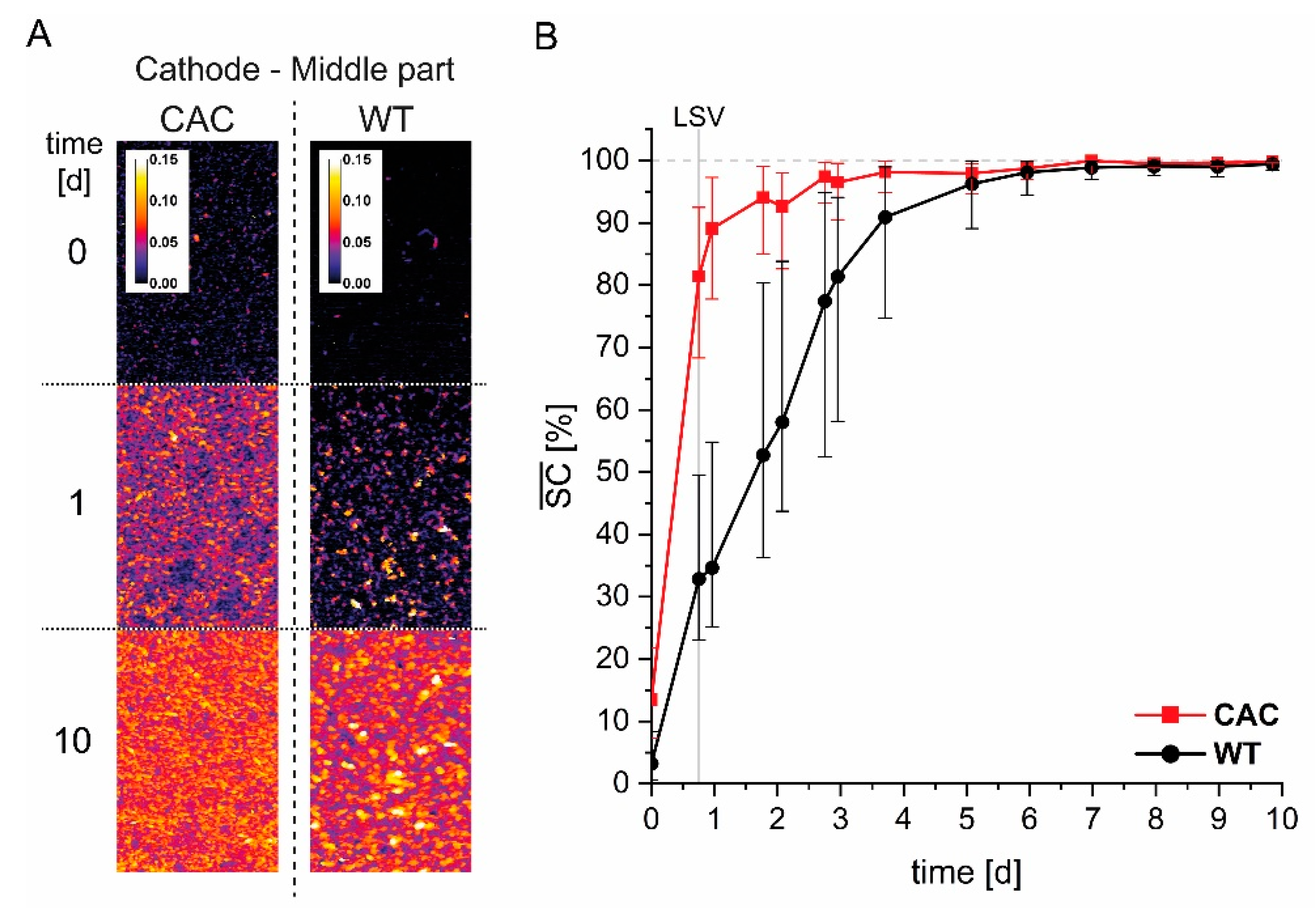
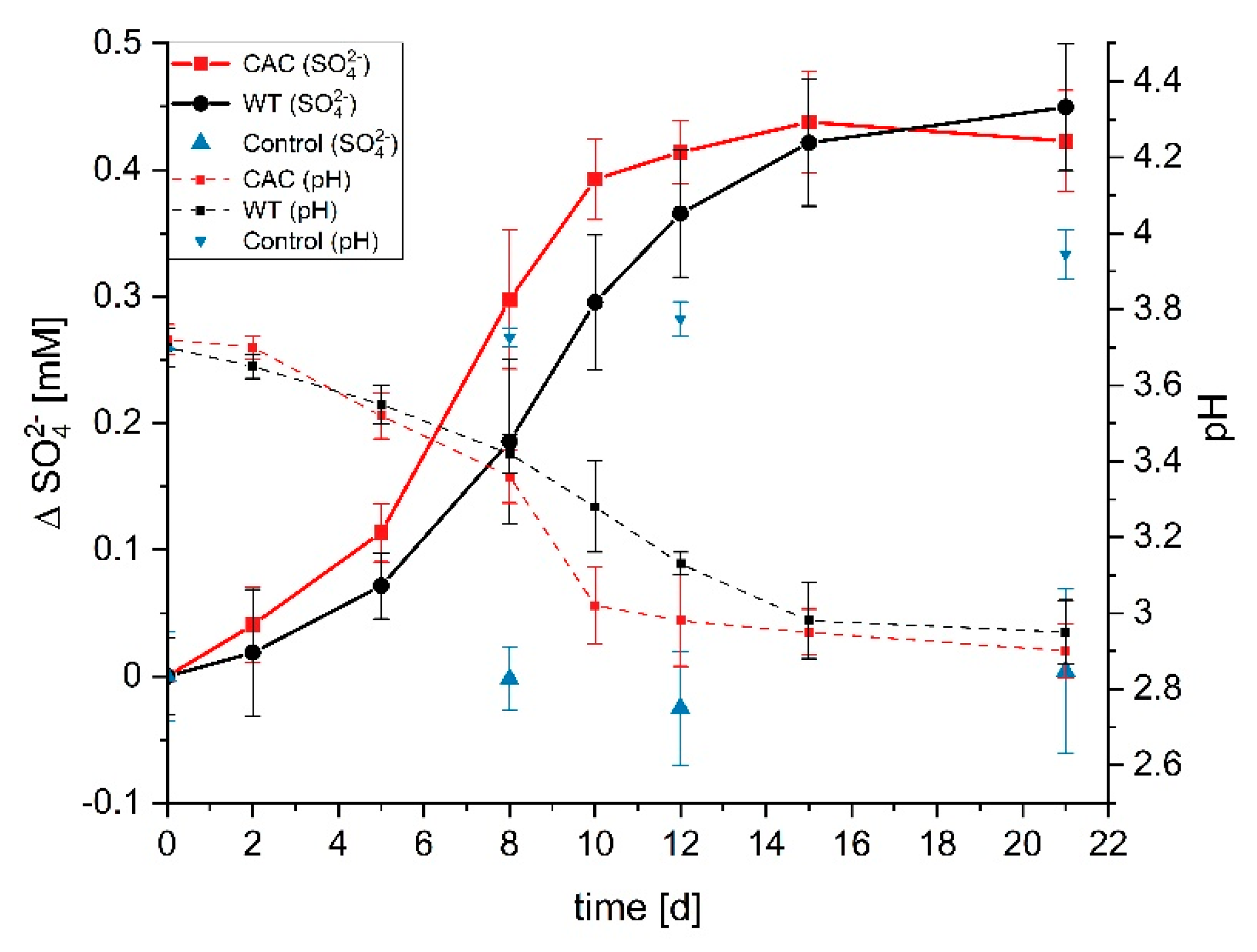
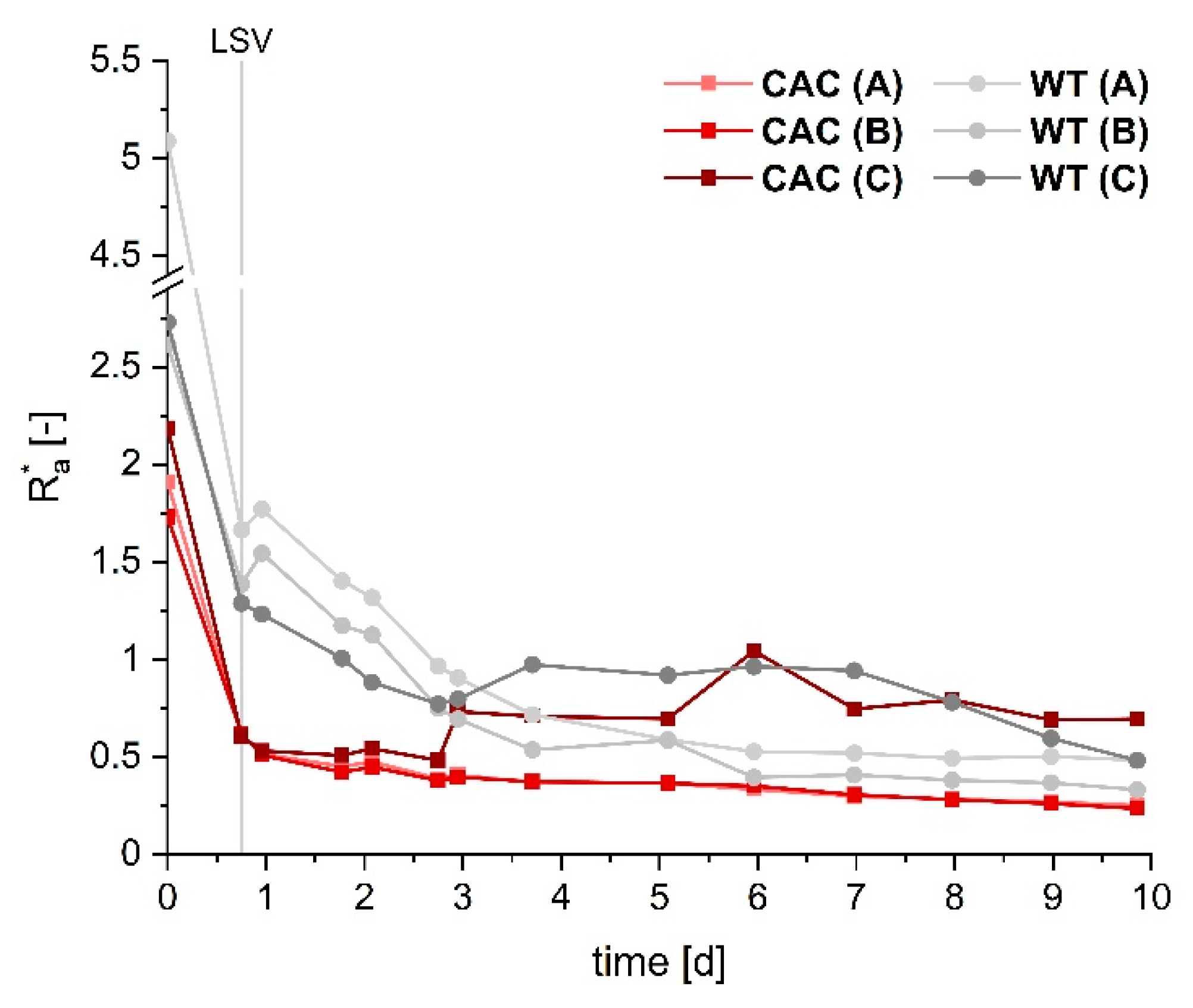
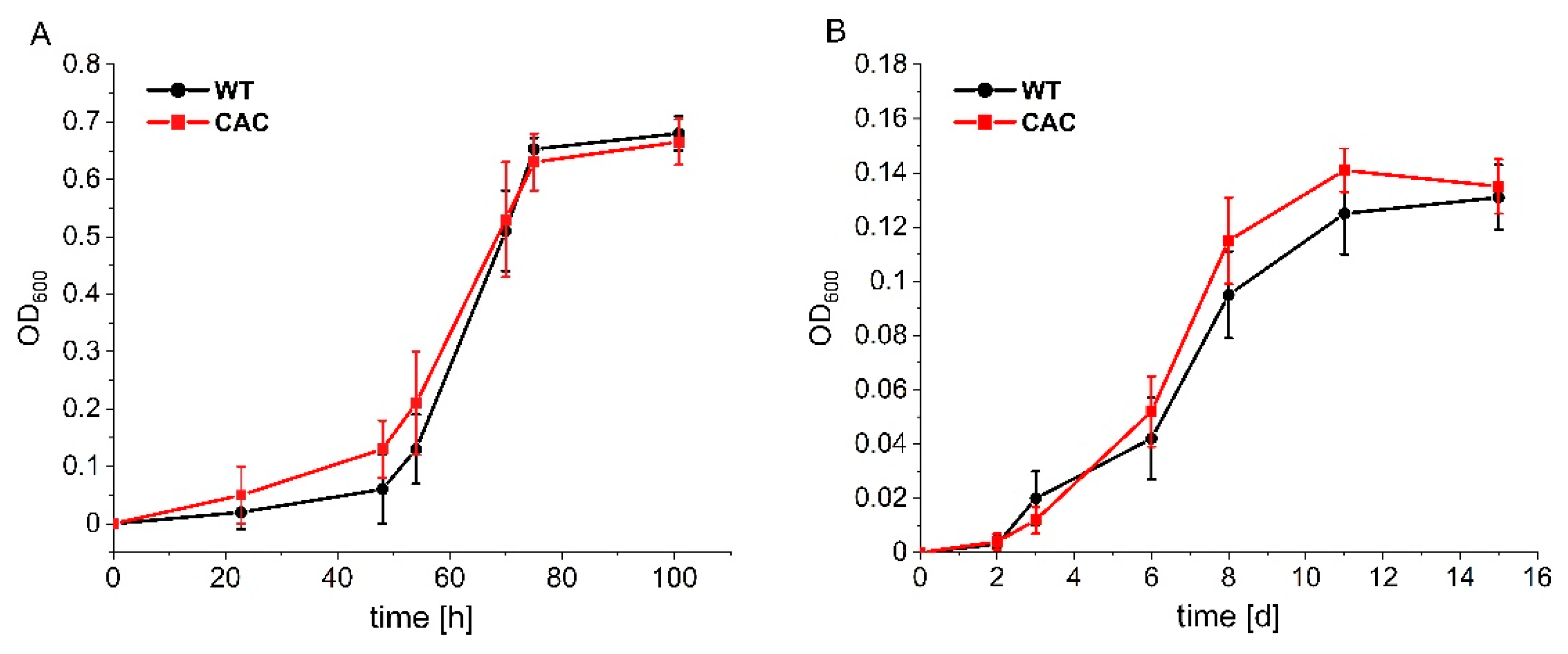
3.1. Cathodic Biofilm Growth
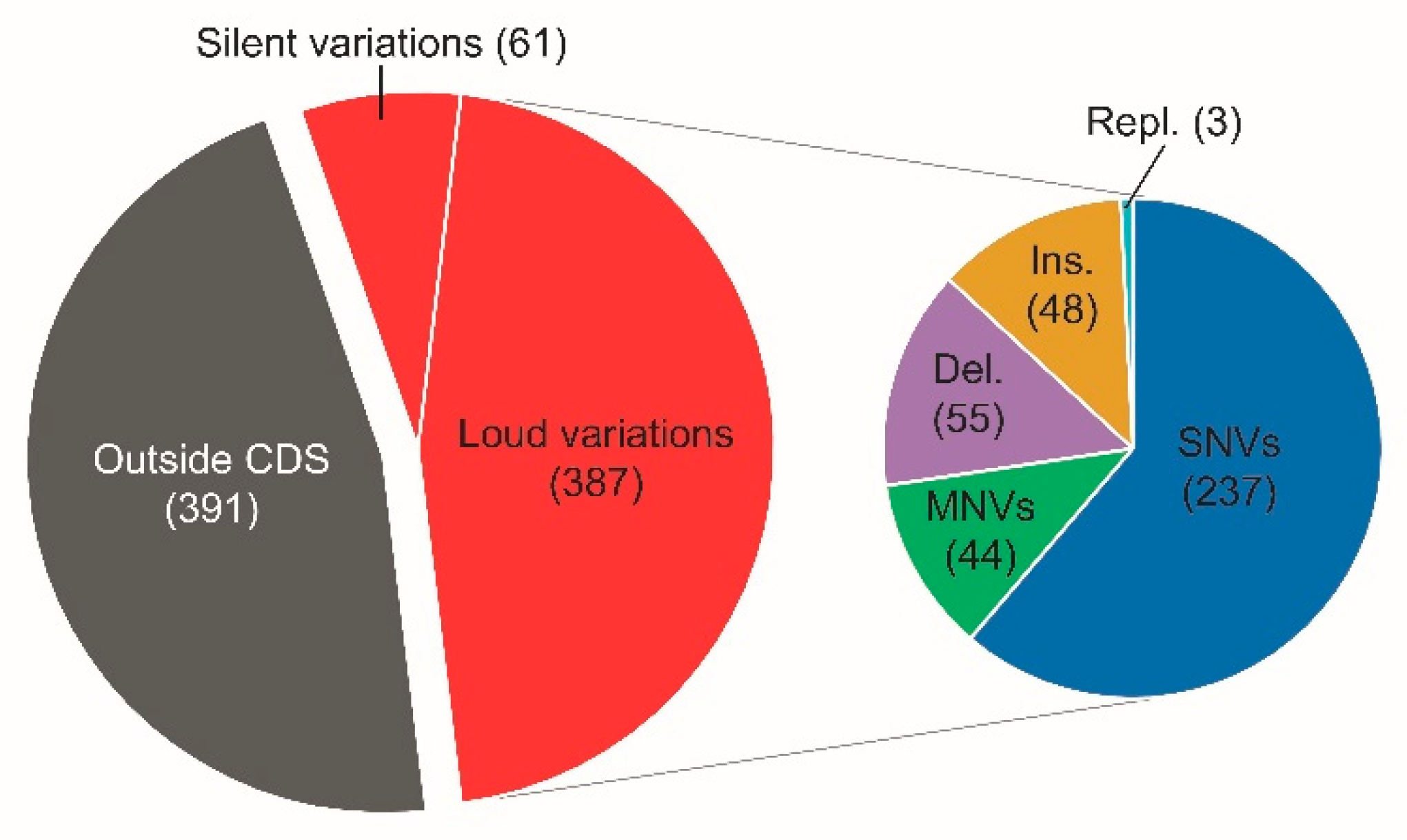
3.2. Comparative Genome Variation Analysis
3.2.1. Adaption to Oxidative Stress
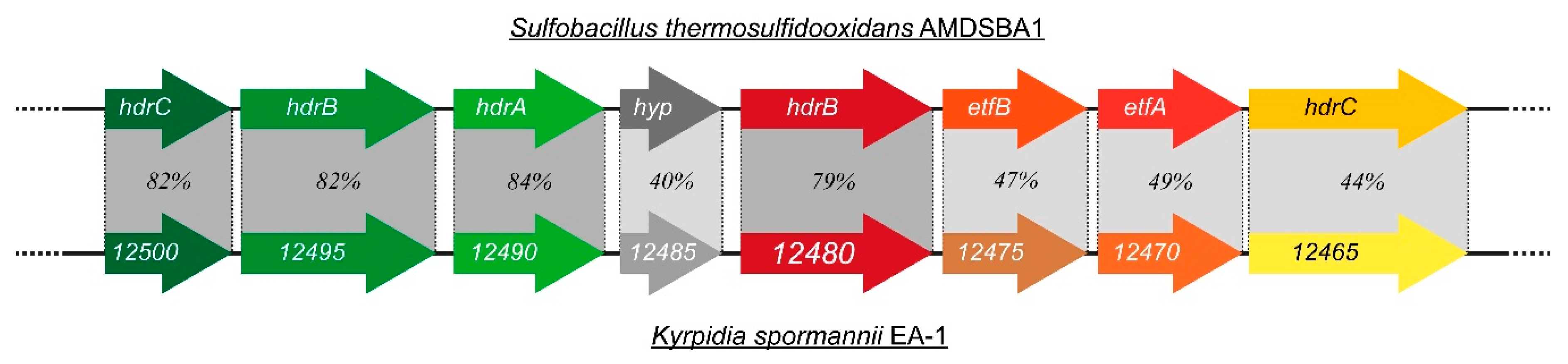
3.2.2. Sulfur-Species as Potential Electron Donor
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mfd(CVV_00395) | |||
| Blast-Analysis: | |||
| Description | Next Relatives | e-Value | Identity [%] |
| transcription-repair coupling factor | Kyrpidia tusciae | 0 | 96.74 |
| Effusibacillus pohliae | 0 | 62.30 | |
| Protein Domain Analysis: | |||
| Protein domain (accession-No.) | Description | e-value | |
| Mfd (COG1197) | transcription-repair coupling factor (superfamily II helicase) | 0 | |
| Mfd (TIGR00580) | transcription-repair coupling factor (mfd) | 0 | |
| CVV_06825 | |||
| Blast-Analysis: | |||
| Description | Next relatives | e-value | Identity [%] |
| transcriptional repressor | Kyrpidia tusciae | 0 | 98.59 |
| peroxide-responsive transcriptional repressor PerR | Halobacillus halophilus | 0 | 60.74 |
| Protein Domain Analysis: | |||
| Protein domain (accession-No.) | Description | e-value | |
| Fur (COG0735) | Fe2+ or Zn2+ uptake regulation protein | 1.6 × e−33 | |
| Fur_like (cd0753) | Ferric uptake regulator (Fur) and related metalloregulatory proteins; DNA-binding repressors and activators | 3.2 × e−32 | |
| CVV_12480 | |||
| Blast-Analysis: | |||
| Description | Next relatives | e-value | Identity [%] |
| (Fe-S)-binding protein | Kyrpidia tusciae | 0 | 99.01 |
| Sulfobacillus thermosulfidooxidans | 0 | 79.11 | |
| Protein Domain Analysis: | |||
| Protein domain (accession-No.) | Description | e-value | |
| GlpC (COG0247) | Fe-S oxidoreductase [Energy production and conversion] | 6.4 × e−32 | |
| PRK06259 | succinate dehydrogenase/fumarate reductase iron–sulfur subunit | 3.4 × e−29 | |
| CoB_CoM_SS_C (TIGR03290) | CoB-CoM heterodisulfide reductase, subunit C; The last step in methanogenesis leaves two coenzymes of methanogenesis, CoM and CoB; Similar enzyme complex subunits are found in various other species, but likely act on a different substrate | 2.9 × e−14 | |

References
- Cheng, S.; Xing, D.; Call, D.F.; Logan, B.E. Direct biological conversion of electrical current into methane by electromethanogenesis. Environ. Sci. Technol. 2009, 43, 3953–3958. [Google Scholar] [CrossRef]
- Nevin, K.P.; Woodard, T.L.; Franks, A.E.; Summers, Z.M.; Lovley, D.R. Microbial electrosynthesis: Feeding microbes electricity to convert carbon dioxide and water to multicarbon extracellular organic compounds. MBio 2010, 1, e00103-10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prévoteau, A.; Carvajal-Arroyo, J.M.; Ganigué, R.; Rabaey, K. Microbial electrosynthesis from CO2: Forever a promise? Curr. Opin. Biotechnol. 2020, 62, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Bajracharya, S.; Srikanth, S.; Mohanakrishna, G.; Zacharia, R.; Strik, D.P.; Pant, D. Biotransformation of carbon dioxide in bioelectrochemical systems: State of the art and future prospects. J. Power Sour. 2017, 356, 256–273. [Google Scholar] [CrossRef]
- Tian, J.-H.; Lacroix, R.; Quéméner, E.D.-L.; Bureau, C.; Midoux, C.; Bouchez, T. Upscaling of microbial electrolysis cell integrating microbial electrosynthesis: Insights, challenges and perspectives. bioRxiv 2019. [Google Scholar] [CrossRef]
- Aryal, N.; Wan, L.; Overgaard, M.H.; Stoot, A.C.; Chen, Y.; Tremblay, P.L.; Zhang, T. Increased carbon dioxide reduction to acetate in a microbial electrosynthesis reactor with a reduced graphene oxide-coated copper foam composite cathode. Bioelectrochemistry 2019, 128, 83–93. [Google Scholar] [CrossRef]
- Kracke, F.; Vassilev, I.; Krömer, J.O. Microbial electron transport and energy conservation—The foundation for optimizing bioelectrochemical systems. Front. Microbiol. 2015, 6, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Choi, O.; Sang, B.-I. Extracellular electron transfer from cathode to microbes: Application for biofuel production. Biotechnol. Biofuels 2016, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- Philips, J. Extracellular electron uptake by acetogenic bacteria: Does H2 consumption favor the H2 evolution reaction on a cathode or metallic iron? Front. Microbiol. 2020, 10, 2997. [Google Scholar] [CrossRef]
- Logan, B.E.; Rossi, R.; Ragab, A.; Saikaly, P.E. Electroactive microorganisms in bioelectrochemical systems. Nat. Rev. Microbiol. 2019, 17, 307–319. [Google Scholar] [CrossRef]
- Rowe, A.R.; Rajeev, P.; Jain, A.; Pirbadian, S.; Okamoto, A.; Gralnick, J.A.; El-Naggar, M.Y.; Nealson, K.H. Tracking electron uptake from a cathode into Shewanella cells: Implications for energy acquisition from solid-substrate electron donors. MBio 2018, 9, e02203-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremblay, P.L.; Angenent, L.T.; Zhang, T. Extracellular electron uptake: Among autotrophs and mediated by surfaces. Trends Biotechnol. 2017, 35, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Lienemann, M.; Deutzmann, J.S.; Milton, R.D.; Sahin, M.; Spormann, A.M. Mediator-free enzymatic electrosynthesis of formate by the Methanococcus maripaludis heterodisulfide reductase supercomplex. Bioresour. Technol. 2018, 254, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, P.L.; Zhang, T. Electrifying microbes for the production of chemicals. Front. Microbiol. 2015, 6, 201. [Google Scholar] [CrossRef] [Green Version]
- Rabaey, K.; Read, S.; Clauwaert, P.; Freguia, S.; Bond, P.L.; Blackall, L.L.; Keller, J. Cathodic oxygen reduction catalyzed by bacteria in microbial fuel cells. ISME J. 2008, 2, 519–527. [Google Scholar] [CrossRef]
- Bause, S.; Decker, M.; Neubauer, P.; Vonau, W. Optimization of the chemolithoautotrophic biofilm growth of Cupriavidus necator by means of electrochemical hydrogen synthesis. Chem. Pap. 2018, 72, 1205–1211. [Google Scholar] [CrossRef]
- Reiner, J.E.; Jung, T.; Lapp, C.J.; Siedler, M.; Bunk, B.; Overmann, J.; Gescher, J. Kyrpidia spormannii sp. nov., a thermophilic, hydrogen-oxidizing, facultative autotroph, isolated from hydrothermal systems at São Miguel Island, and emended description of the genus Kyrpidia. Int. J. Syst. Evol. Microbiol. 2018, 68. [Google Scholar] [CrossRef]
- Hackbarth, M.; Jung, T.; Reiner, J.E.; Gescher, J.; Horn, H.; Hille-Reichel, A.; Wagner, M. Monitoring and quantification of bioelectrochemical Kyrpidia spormannii biofilm development in a novel flow cell setup. Chem. Eng. J. 2020, 390, 124604. [Google Scholar] [CrossRef]
- Reiner, J.E.; Geiger, K.; Hackbarth, M.; Fink, M.; Lapp, C.J.; Jung, T.; Dötsch, A.; Hügler, M.; Wagner, M.; Hille-Reichel, A.; et al. From an extremophilic community to an electroautotrophic production strain: Identifying a novel Knallgas bacterium as cathodic biofilm biocatalyst. ISME J. 2020, 14, 1125–1140. [Google Scholar] [CrossRef]
- Bauer, A.; Wagner, M.; Saravia, F.; Bartl, S.; Hilgenfeldt, V.; Horn, H. In-situ monitoring and quantification of fouling development in membrane distillation by means of optical coherence tomography. J. Memb. Sci. 2019, 577, 145–152. [Google Scholar] [CrossRef]
- Wagner, M.; Horn, H. Optical coherence tomography in biofilm research: A comprehensive review. Biotechnol. Bioeng. 2017, 114, 1386–1402. [Google Scholar] [CrossRef] [PubMed]
- Reiner, J.E.; Lapp, C.J.; Bunk, B.; Spröer, C.; Overmann, J.; Gescher, J. Complete genome sequence of Kyrpidia sp. strain EA-1, a thermophilic knallgas bacterium, isolated from the azores. Genome Announc. 2018, 6, e01505-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashiwagi, A.; Sugawara, R.; Sano Tsushima, F.; Kumagai, T.; Yomo, T. Contribution of silent mutations to thermal adaptation of RNA bacteriophage Q. J. Virol. 2014, 88, 11459–11468. [Google Scholar] [CrossRef] [Green Version]
- Lauring, A.S.; Acevedo, A.; Cooper, S.B.; Andino, R. Codon usage determines the mutational robustness, evolutionary capacity, and virulence of an RNA virus. Cell Host Microbe 2012, 12, 623–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossein Khademi, S.M.; Sazinas, P.; Jelsbak, L. Within-host adaptation mediated by intergenic evolution in Pseudomonas aeruginosa. Genome Biol. Evol. 2019, 11, 1385–1397. [Google Scholar] [CrossRef] [Green Version]
- Roberts, J.; Park, J.S. Mfd, the bacterial transcription repair coupling factor: Translocation, repair and termination. Curr. Opin. Microbiol. 2004, 7, 120–125. [Google Scholar] [CrossRef]
- Selby, C.P. Mfd protein and transcription–repair coupling in Escherichia coli. Photochem. Photobiol. 2017, 93, 280–295. [Google Scholar] [CrossRef] [Green Version]
- De Bruijn, F.J. Stress and Environmental Regulation of Gene Expression and Adaptation in Bacteria; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; Volume 1. [Google Scholar]
- Mongkolsuk, S.; Helmann, J.D. Regulation of inducible peroxide stress responses. Mol. Microbiol. 2002, 45, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Traoré, D.A.K.; El Ghazouani, A.; Ilango, S.; Dupuy, J.; Jacquamet, L.; Ferrer, J.L.; Caux-Thang, C.; Duarte, V.; Latour, J.M. Crystal structure of the apo-PerR-Zn protein from Bacillus subtilis. Mol. Microbiol. 2006, 61, 1211–1219. [Google Scholar] [CrossRef]
- Uversky, V.N.; Permiakov, E.A. Methods in Protein Structure and Stability Analysis: Conformational Stability, Size, Shape, and Surface of Protein Molecules; Nova Science Publishers: Hauppauge, NY, USA, 2007. [Google Scholar]
- Faulkner, M.J.; Ma, Z.; Fuangthong, M.; Helmann, J.D. Derepression of the Bacillus subtilis PerR peroxide stress response leads to iron deficiency. J. Bacteriol. 2012, 194, 1226–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, X.; Koch, T.; Steffens, L.; Finkensieper, J.; Zigann, R.; Cronan, J.E.; Dahl, C. Lipoate-binding proteins and specific lipoate-protein ligases in microbial sulfur oxidation reveal an atpyical role for an old cofactor. Elife 2018, 7, e37439. [Google Scholar] [CrossRef] [PubMed]
- Justice, N.B.; Norman, A.; Brown, C.T.; Singh, A.; Thomas, B.C.; Banfield, J.F. Comparison of environmental and isolate Sulfobacillus genomes reveals diverse carbon, sulfur, nitrogen, and hydrogen metabolisms. BMC Genom. 2014, 15, 1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho, D.; Frazao, R.; Fouillen, A.; Nanci, A.; Lang, B.F.; Apte, S.C.; Baron, C.; Warren, L.A. New Insights into Acidithiobacillus thiooxidans sulfur metabolism through coupled gene expression, solution chemistry, microscopy, and spectroscopy analyses. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Lin, J.Q.; Liu, X.M.; Pang, X.; Zhang, C.J.; Yang, C.L.; Gao, X.Y.; Lin, C.M.; Li, Y.Q.; Li, Y.; et al. Sulfur oxidation in the acidophilic autotrophic Acidithiobacillus spp. Front. Microbiol. 2019, 9, 3290. [Google Scholar] [CrossRef]
- Carbajosa, S.; Malki, M.; Caillard, R.; Lopez, M.F.; Palomares, F.J.; Martín-Gago, J.A.; Rodríguez, N.; Amils, R.; Fernández, V.M.; De Lacey, A.L. Electrochemical growth of Acidithiobacillus ferrooxidans on a graphite electrode for obtaining a biocathode for direct electrocatalytic reduction of oxygen. Biosens. Bioelectron. 2010, 26, 877–880. [Google Scholar] [CrossRef] [Green Version]
- De Campos Rodrigues, T.; Rosenbaum, M.A. Microbial electroreduction: Screening for new cathodic biocatalysts. ChemElectroChem 2014, 1, 1916–1922. [Google Scholar] [CrossRef]
- Deutzmann, J.S.; Spormann, A.M. Enhanced microbial electrosynthesis by using defined co-cultures. ISME J. 2017, 11, 704–714. [Google Scholar] [CrossRef] [Green Version]
- Salimijazi, F.; Parra, E.; Barstow, B. Electrical energy storage with engineered biological systems. J. Biol. Eng. 2019, 13. [Google Scholar] [CrossRef] [Green Version]
- Koch, T.; Dahl, C. A novel bacterial sulfur oxidation pathway provides a new link between the cycles of organic and inorganic sulfur compounds. ISME J. 2018, 12, 2479–2491. [Google Scholar] [CrossRef] [Green Version]
- Kaden, J.; Galushko, A.; Schink, B. Cysteine-mediated electron transfer in syntrophic acetate oxidation by cocultures of Geobacter sulfurreducens and Wolinella succinogenes. Arch. Microbiol. 2002, 178, 53–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aubry, S.; Burlina, F.; Dupont, E.; Delaroche, D.; Joliot, A.; Lavielle, S.; Chassaing, G.; Sagan, S. Cell-surface thiols affect cell entry of disulfide-conjugated peptides. FASEB J. 2009, 23, 2956–2967. [Google Scholar] [CrossRef] [PubMed]
- Pillot, G.; Brillet-viel, A.; Prévost, H.; Secalim, U.M.R.; Bretagne-loire, U. Implication of exofacial thiol groups in the reducing activity of Listeria monocytogenes. bioRxiv 2018, 353409. [Google Scholar] [CrossRef]






| NT# | Annotation | Type | Variation | CR-Change | AA-Change | Frequency [%] |
|---|---|---|---|---|---|---|
| 71257 | mfd | SNV | G → T | 2708G > T | G903V | 55.3 |
| 1352442 | CVV65_06825 | SNV | G → T | 410G > T | C137F | 99.8 |
| 1515328 | - | Ins. | - → GG | - | - | 90.8 |
| 2495265 | CVV65_12480 | SNV | A → C | 932T > G | V311G | 99.8 |
| 2838678 | CVV65_14005 | SNV | T → C | 822A > G | - | 99.4 |
| 3335370 | CVV65_16360 | SNV | G → T | 1425G > T | - | 99.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, T.; Hackbarth, M.; Horn, H.; Gescher, J. Improving the Cathodic Biofilm Growth Capabilities of Kyrpidia spormannii EA-1 by Undirected Mutagenesis. Microorganisms 2021, 9, 77. https://doi.org/10.3390/microorganisms9010077
Jung T, Hackbarth M, Horn H, Gescher J. Improving the Cathodic Biofilm Growth Capabilities of Kyrpidia spormannii EA-1 by Undirected Mutagenesis. Microorganisms. 2021; 9(1):77. https://doi.org/10.3390/microorganisms9010077
Chicago/Turabian StyleJung, Tobias, Max Hackbarth, Harald Horn, and Johannes Gescher. 2021. "Improving the Cathodic Biofilm Growth Capabilities of Kyrpidia spormannii EA-1 by Undirected Mutagenesis" Microorganisms 9, no. 1: 77. https://doi.org/10.3390/microorganisms9010077
APA StyleJung, T., Hackbarth, M., Horn, H., & Gescher, J. (2021). Improving the Cathodic Biofilm Growth Capabilities of Kyrpidia spormannii EA-1 by Undirected Mutagenesis. Microorganisms, 9(1), 77. https://doi.org/10.3390/microorganisms9010077