Potential of Herbaspirillum and Azospirillum Consortium to Promote Growth of Perennial Ryegrass under Water Deficit

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Genome Sequencing, Phylogenomic Analysis, and Annotation
2.3. PGP Effects of Single and Co-Inoculated PGPB in Perennial Ryegrass under Water Deficit
2.4. In Vitro PGP Traits of the Bacterial Co-Inoculation Combinations
2.4.1. Indolic Compounds’ (ICs) Production
2.4.2. EPS Production
2.4.3. VOC Production
2.5. Plant Tissue Colonization in Synthetic Medium
2.6. Data Analysis
3. Results
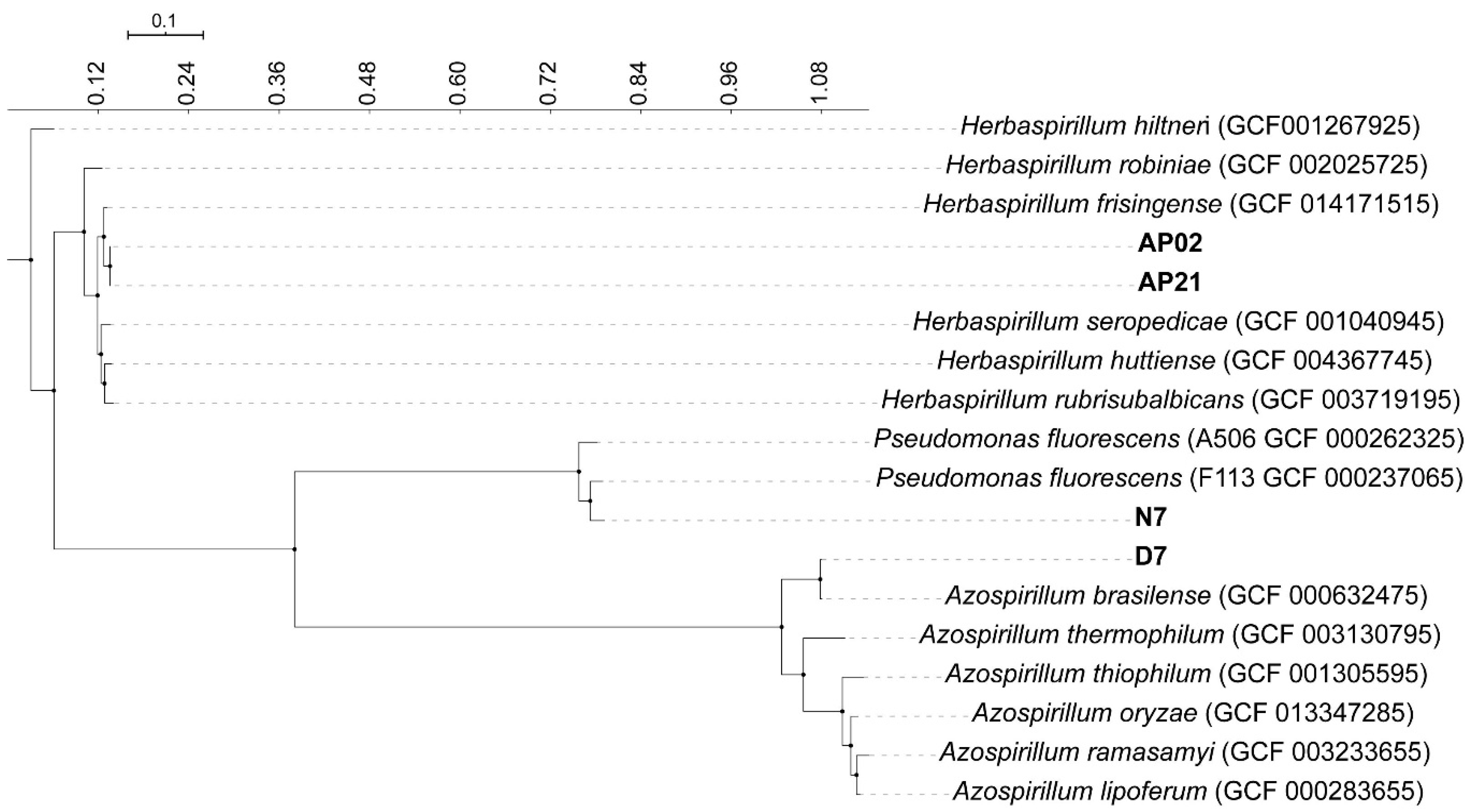
3.1. Phylogenomic Analysis and Annotation of Genes Related to Plant–Microbe Interactions and PGP Traits
3.2. Plant Growth Promotion by Single and Co-Inoculated PGPB under Controlled Conditions
3.3. In Vitro PGP Traits of the Bacteria in the Selected Co-Inoculation Combinations
3.4. Plant Tissue Colonization
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACC | 1-aminocyclopropane-1-carboxylate |
| CFU | Colony forming units |
| DW | Dry weight |
| EPS | Exopolysaccharide |
| FW | Fresh weight |
| IAA | Indole-3-acetic acid |
| IC | Indolic compounds |
| LPS | Lypopolysaccharides |
| PGPB | Plant growth-promoting bacteria |
| RWC | Relative water content |
| TW | Turgid weight |
| VOC | Volatile organic compound |
References
- Compant, S.; Clément, C.; Sessitsch, A. Plant Growth-Promoting Bacteria in the Rhizo- and Endosphere of Plants: Their Role, Colonization, Mechanisms Involved and Prospects for Utilization. Soil Biol. Biochem. 2010, 42, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Kaminsky, L.M.; Trexler, R.V.; Malik, R.J.; Hockett, K.L.; Bell, T.H. The Inherent Conflicts in Developing Soil Microbial Inoculants. Trends Biotechnol. 2019, 37, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, S.R.; Bernstein, H.C.; Song, H.S.; Fredrickson, J.K.; Fields, M.W.; Shou, W.; Johnson, D.R.; Beliaev, A.S. Engineering Microbial Consortia for Controllable Outputs. ISME J. 2016, 10, 2077–2084. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.S.; Nogueira, M.A.; Hungria, M. Microbial Inoculants: Reviewing the Past, Discussing the Present and Previewing an Outstanding Future for the Use of Beneficial Bacteria in Agriculture. AMB Express 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, H.; Singh, V. Potential of Bacterial Endophytes as Plant Growth Promoting Factors. J. Plant Pathol. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Hallmann, J.; Quadt-Hallmann, A.; Mahaffee, W.F.; Kloepper, J.W. Bacterial Endophytes in Agricultural Crops. Can. J. Microbiol. 1997, 43, 895–914. [Google Scholar] [CrossRef]
- Brader, G.; Compant, S.; Mitter, B.; Trognitz, F.; Sessitsch, A. Metabolic Potential of Endophytic Bacteria. Curr. Opin. Biotechnol. 2014, 27, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Carvalhais, L.C.; Crawford, M.; Singh, E.; Dennis, P.G.; Pieterse, C.M.; Schenk, P.M. Inner Plant Values: Diversity, Colonization and Benefits from Endophytic Bacteria. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Puente, M.L.; Gualpa, J.L.; Lopez, G.A.; Molina, R.M.; Carletti, S.M.; Cassán, F.D. The Benefits of Foliar Inoculation with Azospirillum brasilense in Soybean Are Explained by an Auxin Signaling Model. Symbiosis 2018, 76, 41–49. [Google Scholar] [CrossRef]
- Hahn, L.; de Sá, E.L.S.; Osório Filho, B.D.; Machado, R.G.; Damasceno, R.G.; Giongo, A. Rhizobial Inoculation, Alone or Coinoculated with Azospirillum brasilense, Promotes Growth of Wetland Rice. Rev. Bras. Cienc. Solo 2016, 40. [Google Scholar] [CrossRef]
- Zaheer, M.S.; Raza, M.A.S.; Saleem, M.F.; Khan, I.H.; Ahmad, S.; Iqbal, R.; Manevski, K. Investigating the Effect of Azospirillum brasilense and Rhizobium pisi on Agronomic Traits of Wheat (Triticum aestivum L.). Arch. Agron. Soil Sci. 2019, 65, 1554–1564. [Google Scholar] [CrossRef]
- Silva, E.R.; Zoz, J.; Oliveira, C.E.S.; Zuffo, A.M.; Steiner, F.; Zoz, T.; Vendruscolo, E.P. Can Co-Inoculation of Bradyrhizobium and Azospirillum Alleviate Adverse Effects of Drought Stress on Soybean (Glycine max L. merrill.)? Arch. Microbiol. 2019, 201, 325–335. [Google Scholar] [CrossRef]
- Fukami, J.; de la Osa, C.; Ollero, F.J.; Megías, M.; Hungria, M. Co-Inoculation of Maize with Azospirillum brasilense and Rhizobium tropici as a Strategy to Mitigate Salinity Stress. Funct. Plant Biol. 2018, 45, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Bashan, Y.; De-Bashan, L.E. How the Plant Growth-Promoting Bacterium Azospirillum Promotes Plant Growth—A Critical Assessment, 1st ed.; Elsevier Inc.: San Diego, CA, USA, 2010; Volume 108. [Google Scholar] [CrossRef]
- Pereg, L.; de-Bashan, L.E.; Bashan, Y. Assessment of Affinity and Specificity of Azospirillum for Plants. Plant Soil 2016, 399, 389–414. [Google Scholar] [CrossRef]
- Steenhoudt, O.; Vanderleyden, J. Azospirillum, a Free-Living Nitrogen-Fixing Bacterium Closely Associated with Grasses: Genetic, Biochemical and Ecological Aspects. FEMS Microbiol. Rev. 2000, 24, 487–506. [Google Scholar] [CrossRef] [PubMed]
- Muthukumarasamy, R.; Govindarajan, M.; Vadivelu, M.; Revathi, G. N-Fertilizer Saving by the Inoculation of Gluconacetobacter diazotrophicus and Herbaspirillum sp. in Micropropagated Sugarcane Plants. Microbiol. Res. 2006, 161, 238–245. [Google Scholar] [CrossRef]
- Estrada, G.A.; Baldani, V.L.D.; de Oliveira, D.M.; Urquiaga, S.; Baldani, J.I. Selection of Phosphate-Solubilizing Diazotrophic Herbaspirillum and Burkholderia Strains and Their Effect on Rice Crop Yield and Nutrient Uptake. Plant Soil 2013, 369, 115–129. [Google Scholar] [CrossRef]
- Curá, J.A.; Franz, D.R.; Filosofía, J.E.; Balestrasse, K.B.; Burgueño, L.E. Inoculation with Azospirillum sp. and Herbaspirillum sp. Bacteria Increases the Tolerance of Maize to Drought Stress. Microorganisms 2017, 5, 41. [Google Scholar] [CrossRef] [Green Version]
- Santos-Torres, M.; Romero-Perdomo, F.; Mendoza-Labrador, J.; Gutiérrez, A.; Vargas, C.; Castro, E.; Caro-Quintero, A.; Uribe-Vélez, D.; Estrada-Bonilla, G.A. Genomic and Phenotypic Analysis of Rock Phosphate-Solubilizing Rhizobacteria. Rhizosphere 2021, 17, 100290. [Google Scholar] [CrossRef]
- Santoyo, G.; del Orozco-Mosqueda, M.C.; Govindappa, M. Mechanisms of Biocontrol and Plant Growth-Promoting Activity in Soil Bacterial Species of Bacillus and Pseudomonas: A Review. Biocontrol Sci. Technol. 2012, 22, 855–872. [Google Scholar] [CrossRef]
- del Orozco-Mosqueda, M.C.; Duan, J.; DiBernardo, M.; Zetter, E.; Campos-García, J.; Glick, B.R.; Santoyo, G. The Production of ACC Deaminase and Trehalose by the Plant Growth Promoting Bacterium Pseudomonas sp. UW4 Synergistically Protect Tomato Plants against Salt Stress. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, S.; Vaishnav, A.; Jain, S.; Varma, A.; Choudhary, D.K. Induced Drought Tolerance through Wild and Mutant Bacterial Strain Pseudomonas simiae in Mung Bean (Vigna radiata L.). World J. Microbiol. Biotechnol. 2016, 32, 4. [Google Scholar] [CrossRef] [PubMed]
- Bothe, A.; Westermeier, P.; Wosnitza, A.; Willner, E.; Schum, A.; Dehmer, K.J.; Hartmann, S. Drought Tolerance in Perennial Ryegrass (Lolium perenne L.) as Assessed by Two Contrasting Phenotyping Systems. J. Agron. Crop Sci. 2018, 204, 375–389. [Google Scholar] [CrossRef]
- Loaces, I.; Ferrando, L.; Scavino, A.F. Dynamics, Diversity and Function of Endophytic Siderophore-Producing Bacteria in Rice. Microb. Ecol. 2011, 61, 606–618. [Google Scholar] [CrossRef]
- Baldani, J.I.; Reis, V.M.; Videira, S.S.; Boddey, L.H.; Baldani, V.L.D. The Art of Isolating Nitrogen-Fixing Bacteria from Non-Leguminous Plants Using N-Free Semi-Solid Media: A Practical Guide for Microbiologists. Plant Soil 2014, 384, 413–431. [Google Scholar] [CrossRef]
- Andrews, S. Babraham Bioinformatics—FastQC A Quality Control tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 22 October 2020).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Leung, H.C.M.; Yiu, S.M.; Chin, F.Y.L. IDBA-UD: A de Novo Assembler for Single-Cell and Metagenomic Sequencing Data with Highly Uneven Depth. Bioinformatics 2012, 28, 1420–1428. [Google Scholar] [CrossRef] [Green Version]
- Luo, C.; Rodriguez, R.L.M.; Konstantinidis, K.T. MyTaxa: An Advanced Taxonomic Classifier for Genomic and Metagenomic Sequences. Nucleic Acids Res. 2014, 42, e73. [Google Scholar] [CrossRef] [Green Version]
- Yeoman, C.J.; Brutscher, L.M.; Esen, Ö.C.; Ibaoglu, F.; Fowler, C.; Eren, A.M.; Wanner, K.; Weaver, D.K. Genome-Resolved Insights into a Novel Spiroplasma Symbiont of the Wheat Stem Sawfly (Cephus cinctus). PeerJ 2019, 2019, e7548. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. TrimAl: A Tool for Automated Alignment Trimming in Large-Scale Phylogenetic Analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Whelan, S.; Goldman, N. A General Empirical Model of Protein Evolution Derived from Multiple Protein Families Using a Maximum-Likelihood Approach. Mol. Biol. Evol. 2001, 18, 691–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and Clustering Orders of Magnitude Faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nihorimbere, V.; Cawoy, H.; Seyer, A.; Brunelle, A.; Thonart, P.; Ongena, M. Impact of Rhizosphere Factors on Cyclic Lipopeptide Signature from the Plant Beneficial Strain Bacillus amyloliquefaciens S499. FEMS Microbiol. Ecol. 2012, 79, 176–191. [Google Scholar] [CrossRef] [PubMed]
- Soil Survey Staff. Soil Survey Manual Agriculture; USDA: Washington, DC, USA, 2017; Volume 18. [Google Scholar] [CrossRef]
- Sade, N.; Galkin, E.; Moshelion, M. Measuring Arabidopsis, Tomato and Barley Leaf Relative Water Content (RWC). Bio-Protocol 2015, 5, e1451. [Google Scholar] [CrossRef] [Green Version]
- Glickmann, E.; Dessaux, Y. A Critical Examination of the Specificity of the Salkowski Reagent for Indolic Compounds Produced by Phytopathogenic Bacteria. Appl. Environ. Microbiol. 1995, 61, 793–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez Caceres, E.A. Improved Medium for Isolation of Azospirillum spp. Appl. Environ. Microbiol. 1982, 44, 990–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Galván, A.E.; Cortés-Patiño, S.; Romero-Perdomo, F.; Uribe-Vélez, D.; Bashan, Y.; Bonilla, R.R. Proline Accumulation and Glutathione Reductase Activity Induced by Drought-Tolerant Rhizobacteria as Potential Mechanisms to Alleviate Drought Stress in Guinea Grass. Appl. Soil Ecol. 2020, 147, 103367. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Park, Y.S.; Dutta, S.; Ann, M.; Raaijmakers, J.M.; Park, K. Promotion of Plant Growth by Pseudomonas fluorescens Strain SS101 via Novel Volatile Organic Compounds. Biochem. Biophys. Res. Commun. 2015, 461, 361–365. [Google Scholar] [CrossRef]
- Balsanelli, E.; Serrato, R.V.; de Baura, V.A.; Sassaki, G.; Yates, M.G.; Rigo, L.U.; Pedrosa, F.O.; de Souza, E.M.; Monteiro, R.A. Herbaspirillum seropedicae RfbB and RfbC Genes Are Required for Maize Colonization. Environ. Microbiol. 2010, 12, 2233–2244. [Google Scholar] [CrossRef]
- Glick, B.R.; Cheng, Z.; Czarny, J.; Duan, J. Promotion of Plant Growth by ACC Deaminase-Producing Soil Bacteria. Eur. J. Plant Pathol. 2007, 119, 329–339. [Google Scholar] [CrossRef]
- Wisniewski-Dyé, F.; Lozano, L.; Acosta-Cruz, E.; Borland, S.; Drogue, B.; Prigent-Combaret, C.; Rouy, Z.; Barbe, V.; Herrera, A.M.; González, V.; et al. Genome Sequence of Azospirillum brasilense CBG497 and Comparative Analyses of Azospirillum Core and Accessory Genomes Provide Insight into Niche Adaptation. Genes (Basel) 2012, 3, 576–602. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.; Castro-Sowinski, S.; Valverde, A.; Lerner, H.; Dror, R.; Okon, Y.; Burdman, S. The Azospirillum brasilense sp7 NoeJ and NoeL Genes Are Involved in Extracellular Polysaccharide Biosynthesis. Microbiology 2009, 155, 4058–4068. [Google Scholar] [CrossRef] [Green Version]
- Rossi, F.A.; Medeot, D.B.; Liaudat, J.P.; Pistorio, M.; Jofré, E. In Azospirillum brasilense, Mutations in FlmA or FlmB Genes Affect Polar Flagellum Assembly, Surface Polysaccharides, and Attachment to Maize Roots. Microbiol. Res. 2016, 190, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Cordovez, V.; Etalo, D.W.; van der Voort, M.; Raaijmakers, J.M. Role of the GacS Sensor Kinase in the Regulation of Volatile Production by Plant Growth-Promoting Pseudomonas fluorescens SBW25. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Hatier, J.H.B.; Faville, M.J.; Hickey, M.J.; Koolaard, J.P.; Schmidt, J.; Carey, B.L.; Jones, C.S. Plant Vigour at Establishment and Following Defoliation Are Both Associated with Responses to Drought in Perennial Ryegrass (Lolium perenne L.). J. Exp. Bot. 2014. [Google Scholar] [CrossRef] [Green Version]
- Redondo-Nieto, M.; Barret, M.; Morrissey, J.; Germaine, K.; Martínez-Granero, F.; Barahona, E.; Navazo, A.; Sánchez-Contreras, M.; Moynihan, J.A.; Muriel, C.; et al. Genome Sequence Reveals That Pseudomonas fluorescens F113 Possesses a Large and Diverse Array of Systems for Rhizosphere Function and Host Interaction. BMC Genom. 2013, 14, 54. [Google Scholar] [CrossRef] [Green Version]
- Lucas García, J.A.; Probanza, A.; Ramos, B.; Barriuso, J.; Gutierrez Mañero, F.J. Effects of Inoculation with Plant Growth Promoting Rhizobacteria (PGPRs) and Sinorhizobium Fredii on Biological Nitrogen Fixation, Nodulation and Growth of Glycine Max Cv. Osumi. Plant Soil 2005, 267, 145–153. [Google Scholar] [CrossRef]
- Rosier, A.; Bishnoi, U.; Lakshmanan, V.; Sherrier, D.J.; Bais, H.P. A Perspective on Inter-Kingdom Signaling in Plant–Beneficial Microbe Interactions. Plant Mol. Biol. 2016, 90, 537–548. [Google Scholar] [CrossRef]
- Helman, Y.; Chernin, L. Silencing the Mob: Disrupting Quorum Sensing as a Means to Fight Plant Disease. Mol. Plant Pathol. 2015, 16, 316–329. [Google Scholar] [CrossRef]
- Maroniche, G.A.; Diaz, P.R.; Borrajo, M.P.; Valverde, C.F.; Creus, C.M. Friends or Foes in the Rhizosphere: Traits of Fluorescent Pseudomonas That Hinder Azospirillum brasilense Growth and Root Colonization. FEMS Microbiol. Ecol. 2018, 94. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Mishra, S.; Dixit, V.; Kumar, M.; Agarwal, L.; Chauhan, P.S.; Nautiyal, C.S. Synergistic Effect of Pseudomonas putida and Bacillus amyloliquefaciens Ameliorates Drought Stress in Chickpea (Cicer arietinum L.). Plant Signal. Behav. 2016, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakar, K.U.; Ren, X.L.; Nawaz, Z.; Cui, Z.Q.; Li, B.; Xie, G.L.; Hassan, M.A.; Ali, E.; Sun, G.C. A Consortium of Rhizobacterial Strains and Biochemical Growth Elicitors Improve Cold and Drought Stress Tolerance in Rice (Oryza sativa L.). Plant Biol. 2016, 18, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Verma, J.P. Does Plant—Microbe Interaction Confer Stress Tolerance in Plants: A Review? Microbiol. Res. 2018, 207, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Shiva, S.; Vurukonda, K.P.; Vardharajula, S.; Shrivastava, M.; Skz, A. Enhancement of Drought Stress Tolerance in Crops by Plant Growth Promoting Rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef]
- Kaushal, M.; Wani, S.P. Rhizobacterial-Plant Interactions: Strategies Ensuring Plant Growth Promotion under Drought and Salinity Stress. Agric. Ecosyst. Environ. 2016, 231, 68–78. [Google Scholar] [CrossRef]
- Radwan, T.E.S.E.D.; Mohamed, Z.K.; Reis, V.M. Production of Indole-3-Acetic Acid by Different Strains of Azospirillum and Herbaspirillum spp. Symbiosis 2002, 32, 39–54. [Google Scholar]
- El-Desouk Radwan, T.E.S.; Mohamed, Z.K.; Reis, V.M. Efeito Da Inoculação de Azospirillum e Herbaspirillum Na Produção de Compostos Indólicos Em Plântulas de Milho e Arroz. Pesqui. Agropecu. Bras. 2004, 39, 987–994. [Google Scholar] [CrossRef]
- van Puyvelde, S.; Cloots, L.; Engelen, K.; Das, F.; Marchal, K.; Vanderleyden, J.; Spaepen, S. Transcriptome Analysis of the Rhizosphere Bacterium Azospirillum brasilense Reveals an Extensive Auxin Response. Microb. Ecol. 2011, 61, 723–728. [Google Scholar] [CrossRef]
- Kim, J.; Park, W. Indole: A Signaling Molecule or a Mere Metabolic Byproduct That Alters Bacterial Physiology at a High Concentration? J. Microbiol. 2015, 53, 421–428. [Google Scholar] [CrossRef]
- Somers, E.; Ptacek, D.; Gysegom, P.; Srinivasan, M.; Vanderleyden, J. Azospirillum brasilense Produces the Auxin-Like Phenylacetic Acid by Using. Appl. Environ. Microbiol. 2005, 71, 1803–1810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naseem, H.; Ahsan, M.; Shahid, M.A.; Khan, N. Exopolysaccharides Producing Rhizobacteria and Their Role in Plant Growth and Drought Tolerance. J. Basic Microbiol. 2018, 58, 1009–1022. [Google Scholar] [CrossRef] [PubMed]
- Bashan, Y.; Levanony, H.; Whitmoyer, R.E. Root Surface Colonization of Non-Cereal Crop Plants by Pleomorphic Azospirillum brasilense Cd. J. Gen. Microbiol. 1991, 137, 187–196. [Google Scholar] [CrossRef] [Green Version]
- Viruega-góngora, V.I.; Acatitla-jácome, I.S.; Reyes-carmona, S.R.; Baca, B.E.; Ramírez-mata, A. Spatio-Temporal Formation of Biofilms and Extracellular Matrix Analysis in Azospirillum brasilense. FEMS Microbiol. Lett. 2020, 367, fnaa037. [Google Scholar] [CrossRef] [PubMed]
- Konnova, S.A.; Brykova, O.S.; Sachkova, O.A.; Egorenkova, I.V.; Ignatov, V.V. Protective Role of the Polysaccharide-Containing Capsular Components of Azospirillum brasilense. Microbiology 2001, 70, 436–440. [Google Scholar] [CrossRef]
- El-Komy, H.M.; Hamdia, M.A.; Abd El-Baki, G.K. Nitrate Reductase in Wheat Plants Grown under Water Stress and Inoculated with Azospirillum spp. Biologia Plantarum. 2003, 281–287. [Google Scholar] [CrossRef]
- Oliveira, A.L.M.; Santos, O.J.A.P.; Marcelino, P.R.F.; Milani, K.M.L.; Zuluaga, M.Y.A.; Zucareli, C.; Gonçalves, L.S.A. Maize Inoculation with Azospirillum brasilense Ab-V5 Cells Enriched with Exopolysaccharides and Polyhydroxybutyrate Results in High Productivity under Low N Fertilizer Input. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Arzanesh, M.H.; Alikhani, H.A.; Khavazi, K.; Rahimian, H.A.; Miransari, M. Wheat (Triticum aestivum L.) Growth Enhancement by Azospirillum sp. under Drought Stress. World J. Microbiol. Biotechnol. 2011, 27, 197–205. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A. Exopolysaccharide Producing Rhizobacteria and Their Impact on Growth and Drought Tolerance of Wheat Grown under Rainfed Conditions. PLoS ONE 2019, 14, e0222302. [Google Scholar] [CrossRef] [Green Version]
- Passos da Silva, D.; Matwichuk, M.L.; Townsend, D.O.; Reichhardt, C.; Lamba, D.; Wozniak, D.J.; Parsek, M.R. The Pseudomonas aeruginosa Lectin LecB Binds to the Exopolysaccharide Psl and Stabilizes the Biofilm Matrix. Nat. Commun. 2019, 10, 2183. [Google Scholar] [CrossRef]
- Kanchiswamy, C.N.; Malnoy, M.; Maffei, M.E. Bioprospecting Bacterial and Fungal Volatiles for Sustainable Agriculture. Trends Plant Sci. 2015, 20, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Solís, D.; Zetter-Salmón, E.; Contreras-Pérez, M.; del Rocha-Granados, M.C.; Macías-Rodríguez, L.; Santoyo, G. Pseudomonas stutzeri E25 and Stenotrophomonas maltophilia CR71 Endophytes Produce Antifungal Volatile Organic Compounds and Exhibit Additive Plant Growth-Promoting Effects. Biocatal. Agric. Biotechnol. 2018, 13, 46–52. [Google Scholar] [CrossRef]
- Sheoran, N.; Kumar, A.; Munjal, V.; Nadakkakath, A.V.; Eapen, S.J. Pseudomonas Putida BP25 Alters Root Phenotype and Triggers Salicylic Acid Signaling as a Feedback Loop in Regulating Endophytic Colonization in Arabidopsis Thaliana. Physiol. Mol. Plant. Pathol. 2016, 93, 99–111. [Google Scholar] [CrossRef]
- Ossowicki, A.; Jafra, S.; Garbeva, P. The Antimicrobial Volatile Power of the Rhizospheric Isolate Pseudomonas donghuensis P482. PLoS ONE 2017, 12, e0174362. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, T.; Liu, Y.; Li, X.; Zhang, C.; Feng, Z.; Peng, X.; Li, Z.; Qin, S.; Xing, K. Volatile Organic Compounds Produced by Pseudomonas chlororaphis Subsp. Aureofaciens SPS-41 as Biological Fumigants to Control Ceratocystis Fimbriata in Postharvest Sweet Potatoes. J. Agric. Food Chem. 2019, 67, 3702–3710. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| PGP Trait | Gene | Protein | AP02 | AP21 | D7 | N7 | Reference |
|---|---|---|---|---|---|---|---|
| ACC deaminase | acdS | 1-aminocyclopropane-1-carboxylate deaminase | + | + | − | + | Glick et al. [44] |
| Indolic compounds production | iaaM | Monoamine oxidase | - | - | - | + | Wisniewski-Dyé et al. [45] |
| ipdC | Indole-3-pyruvate decarboxylase | + | - | + | - | Wisniewski-Dyé et al. [45] | |
| Exopolysaccharides (EPS) and Lypopolysaccharides production (LPS) | noeJ | Mannose-1-phosphate guanylyltransferase | - | - | - | - | Lerner et al. [46] |
| noeL | GDP-mannose 4,6-dehydratase | - | - | - | - | Lerner et al. [46] | |
| rfbB | dTDP-glucose 4,6-dehydratase | - | - | + | + | Balsanelli et al. [43] | |
| rfbC | dTDP-4-dehydrorhamnose 3,5-epimerase | - | - | + | - | Balsanelli et al. [43] | |
| Colonization: Biofilm formation | flmA | Putative sugar nucleotide epimerase/dehydratase | - | - | + | - | Rossi et al. [47] |
| flmB | Flagellin modification protein FlmB | - | - | + | - | Rossi et al. [47] | |
| Volatile Organic Compounds | gacA | Response regulator GacA | - | - | - | + | Cheng et al. [48] |
| gacS | Histidine kinase | - | - | - | + | Cheng et al. [48] |
| Treatment | Indolic Compounds | ||
|---|---|---|---|
| μg mL−1 | μg LogCFU−1 | Morphology Observed | |
| AP02 | 42.9 ± 3.0 b | 13.5 ± 1.1 bc | Herbaspirillum |
| AP21 | 45.5 ± 1.8 b | 11.9 ± 0.5 b | Herbaspirillum |
| D7 | 69.7 ± 1.0 c | 12.4 ± 1.3 bc | Azospirillum |
| N7 | 11.9 ± 1.6 a | 1.7 ± 0.2 a | Pseudomonas |
| AP02 + AP21 | 39.7 ± 0.3 b | 7.5 ± 0.3 b | Herbaspirillum |
| AP02 + N7 | 14.8 ± 3.7 a | 2.3 ± 0.5 a | Pseudomonas |
| AP21 + D7 | 90.8 ± 3.9 d | 14.5 ± 1.0 c | Azospirillum |
| Treatment | Exopolysaccharides | ||
| mg mL−1 | mg LogCFU−1 | Morphology Observed | |
| AP02 | 4.8 ± 0.5 b | 1.0 ± 0.1 b | Herbaspirillum |
| AP21 | 5.4 ± 0.6 b | 1.0 ± 0.1 b | Herbaspirillum |
| D7 | 24.5 ± 1.0 d | 3.7 ± 0.3 d | Azospirillum |
| N7 | 2.4 ± 0.7 a | 0.3 ± 0.0 a | Pseudomonas |
| AP02 + AP21 | 4.6 ± 1.0 b | 0.8 ± 0.2 b | Herbaspirillum |
| AP02 + N7 | 2.6 ± 0.3 a | 0.3 ± 0.0 a | Pseudomonas |
| AP21 + D7 | 14.9 ± 1.1 c | 3.0 ± 0.2 c | Azospirillum + Herbaspirillum |
| Treatment | Seed Germination | Length (mm) | |
|---|---|---|---|
| (%) | Coleoptile | Radicle | |
| Uninoculated | 70.0 ± 5.7 a | 4.6 ± 1.3 a | 8.2 ± 0.6 a |
| AP02 | 67.5 ± 4.2 a | 6.0 ± 1.3 a | 12.0 ± 1.1 ab |
| AP21 | 78.3± 8.4 a | 4.6 ± 0.5 a | 10.3 ± 1.7ab |
| D7 | 74.1 ± 15.7 a | 6.0 ± 0.7 a | 12.4 ± 1.3 ab |
| N7 | 63.3 ± 11.5 a | 7.4 ± 1.9 a | 14.5 ± 1.8 b |
| AP02 + AP21 | 62.5 ± 6.8 a | 6.2 ± 1.2 a | 13.2 ± 2.5 b |
| AP02 + N7 | 63.3 ± 14.1 a | 5.0 ± 0.6 a | 11.2 ± 1.8 ab |
| AP21 + D7 | 65.8 ± 2.6 a | 5.2 ± 1.8 a | 11.7 ± 0.7 ab |
| Treatment | Cell Count (CFU g−1) | |
|---|---|---|
| Shoot | Root | |
| Uninoculated | 2.5 × 103 | 4.0 × 102 |
| AP02 | 7.5 × 105 | 7.0 × 105 |
| AP21 | 9.5 × 105 | 2.0 × 107 |
| D7 | 1.1 × 107 | 1.5 × 102 |
| N7 | 1.5 × 109 | 7.0 × 106 |
| AP21 + D7 | 4.5 × 106 | 4.5 × 104 |
| AP02 + D7 | 4.5 × 106 | 4.5 × 106 |
| AP02 + N7 | 7.0 × 106 | 1.5 × 107 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cortés-Patiño, S.; Vargas, C.; Álvarez-Flórez, F.; Bonilla, R.; Estrada-Bonilla, G. Potential of Herbaspirillum and Azospirillum Consortium to Promote Growth of Perennial Ryegrass under Water Deficit. Microorganisms 2021, 9, 91. https://doi.org/10.3390/microorganisms9010091
Cortés-Patiño S, Vargas C, Álvarez-Flórez F, Bonilla R, Estrada-Bonilla G. Potential of Herbaspirillum and Azospirillum Consortium to Promote Growth of Perennial Ryegrass under Water Deficit. Microorganisms. 2021; 9(1):91. https://doi.org/10.3390/microorganisms9010091
Chicago/Turabian StyleCortés-Patiño, Sandra, Christian Vargas, Fagua Álvarez-Flórez, Ruth Bonilla, and German Estrada-Bonilla. 2021. "Potential of Herbaspirillum and Azospirillum Consortium to Promote Growth of Perennial Ryegrass under Water Deficit" Microorganisms 9, no. 1: 91. https://doi.org/10.3390/microorganisms9010091
APA StyleCortés-Patiño, S., Vargas, C., Álvarez-Flórez, F., Bonilla, R., & Estrada-Bonilla, G. (2021). Potential of Herbaspirillum and Azospirillum Consortium to Promote Growth of Perennial Ryegrass under Water Deficit. Microorganisms, 9(1), 91. https://doi.org/10.3390/microorganisms9010091