
The Nasopharyngeal, Ruminal, and Vaginal Microbiota and the Core Taxa Shared across These Microbiomes in Virgin Yearling Heifers Exposed to Divergent In Utero Nutrition during Their First Trimester of Gestation and in Pregnant Beef Heifers in Response to Mineral Supplementation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Husbandry and Experimental Design
2.1.1. Virgin Yearling Heifers
2.1.2. Pregnant Heifers
2.2. Nasopharyngeal Swab, Ruminal Fluid, and Vaginal Swab Sampling
2.2.1. Nasopharyngeal Sampling
2.2.2. Rumen Fluid Sampling
2.2.3. Vaginal Sampling
2.3. Metagenomic DNA Extraction
2.4. 16S rRNA Gene Sequencing and Analysis
2.5. Statistical Analysis
3. Results
3.1. Sequencing Results
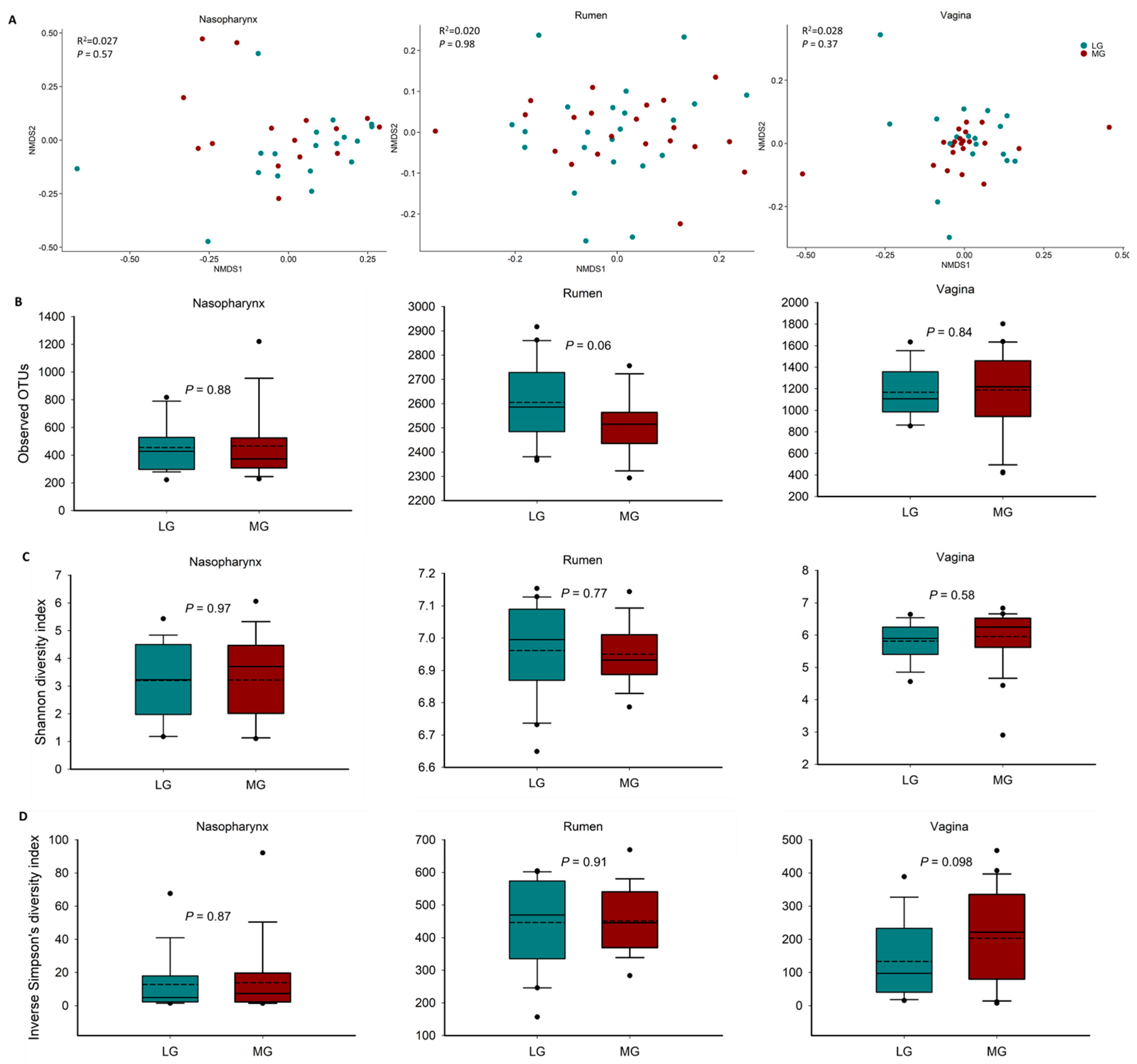
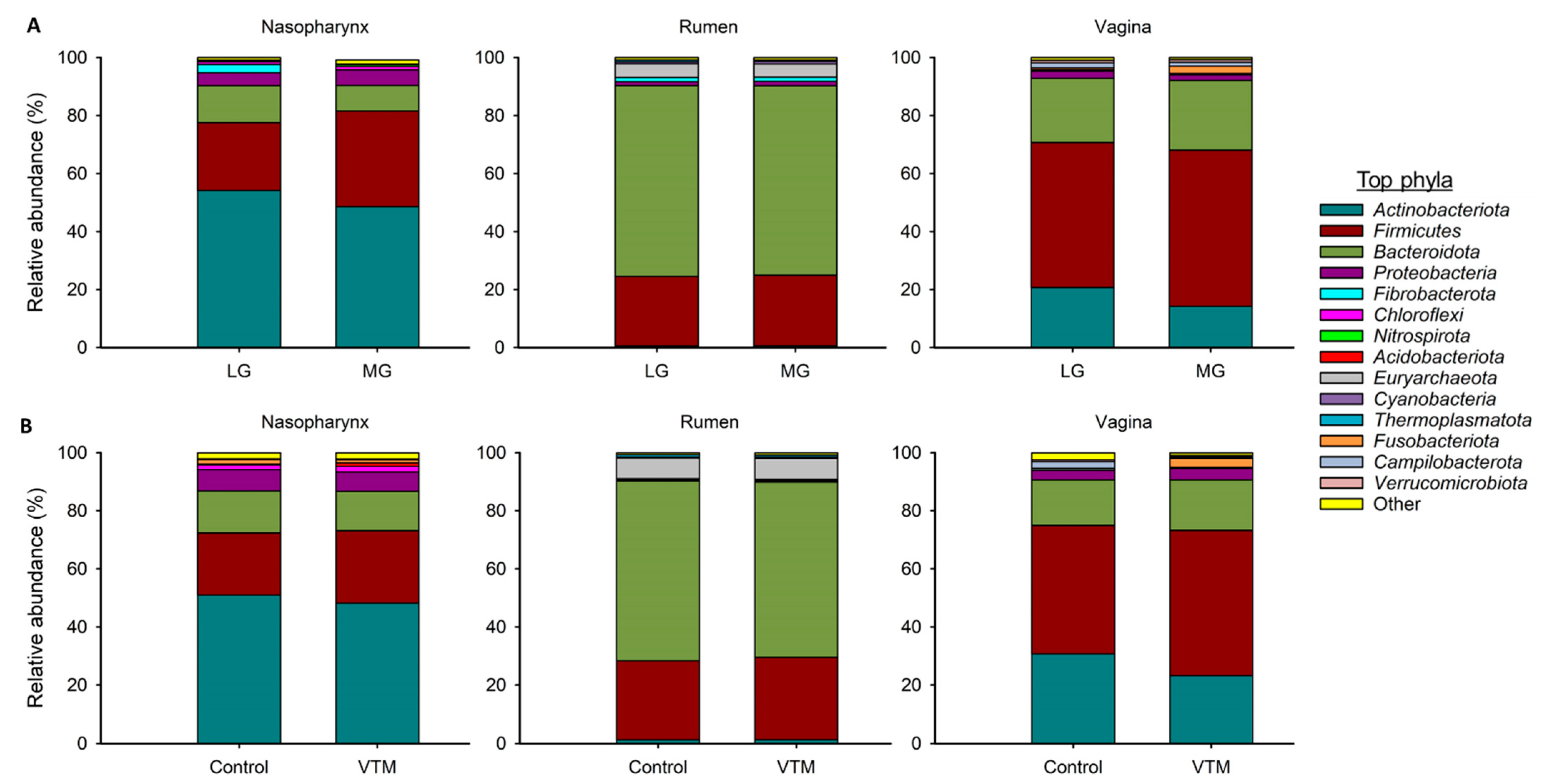
3.2. Effect of Maternal Restricted Gain during the First Trimester of Gestation on Offspring Microbiota Development
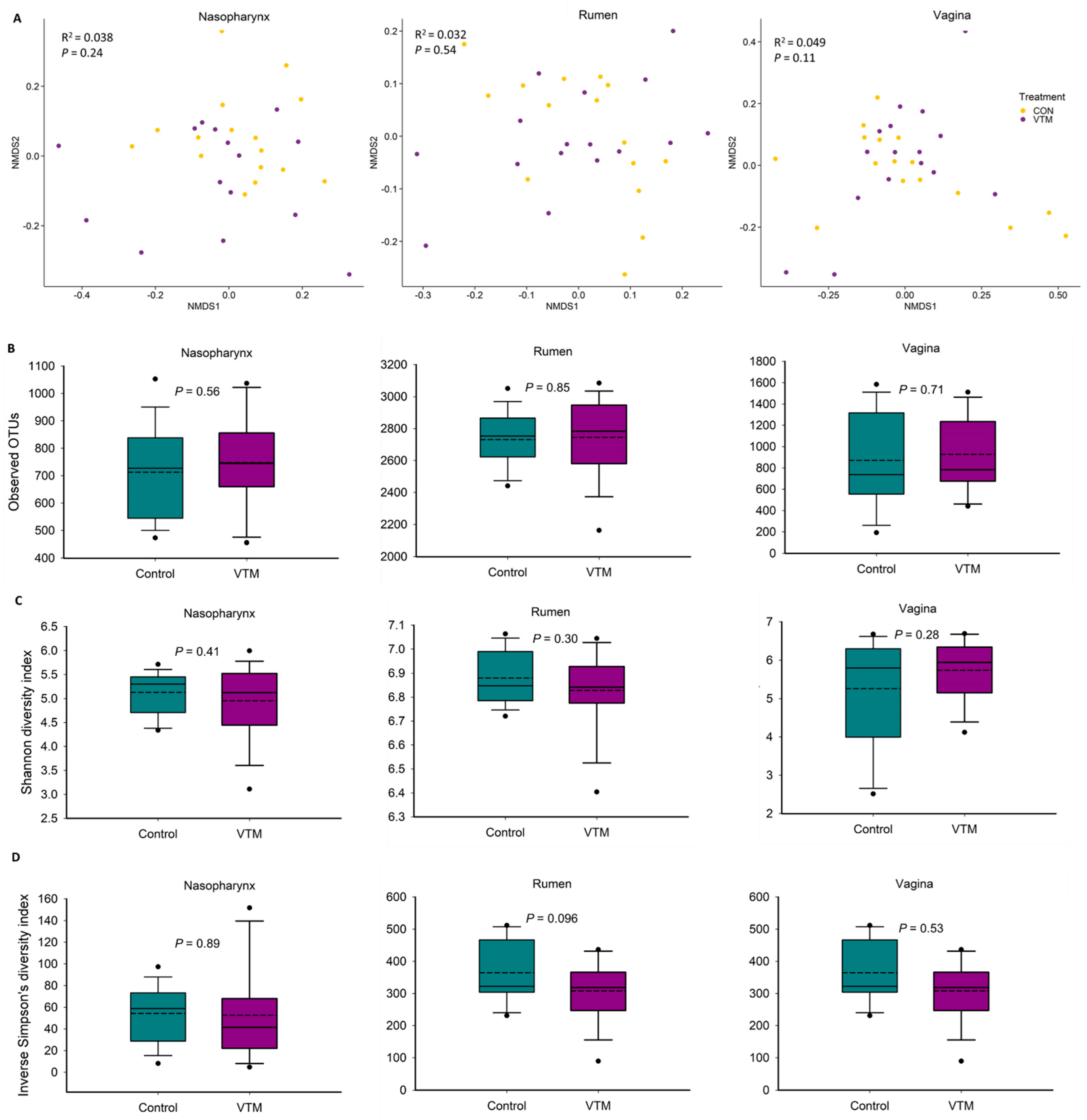
3.3. Effect of Vitamin and Mineral Supplementation during the First Six Months of Gestation on the Maternal Microbiota
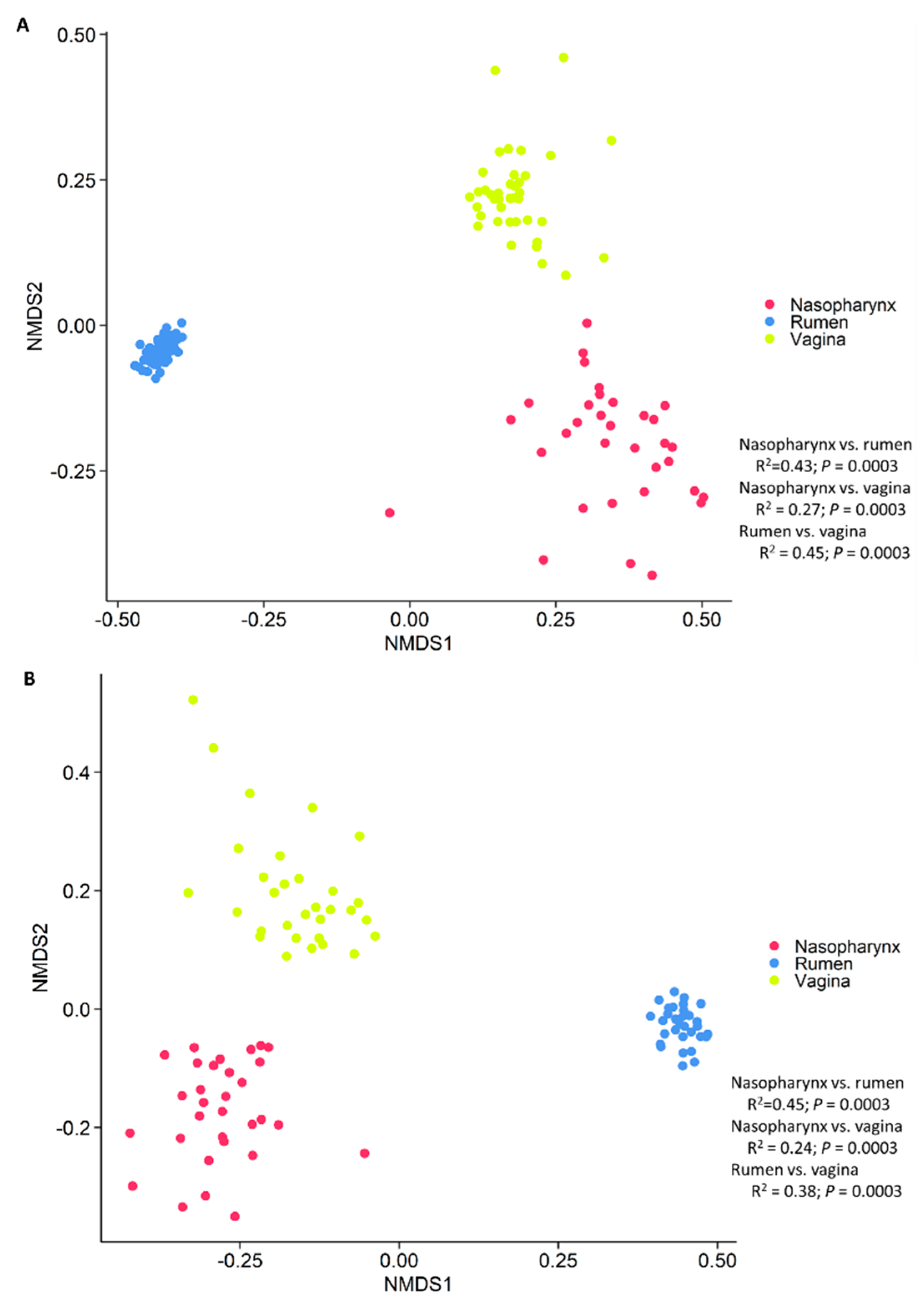
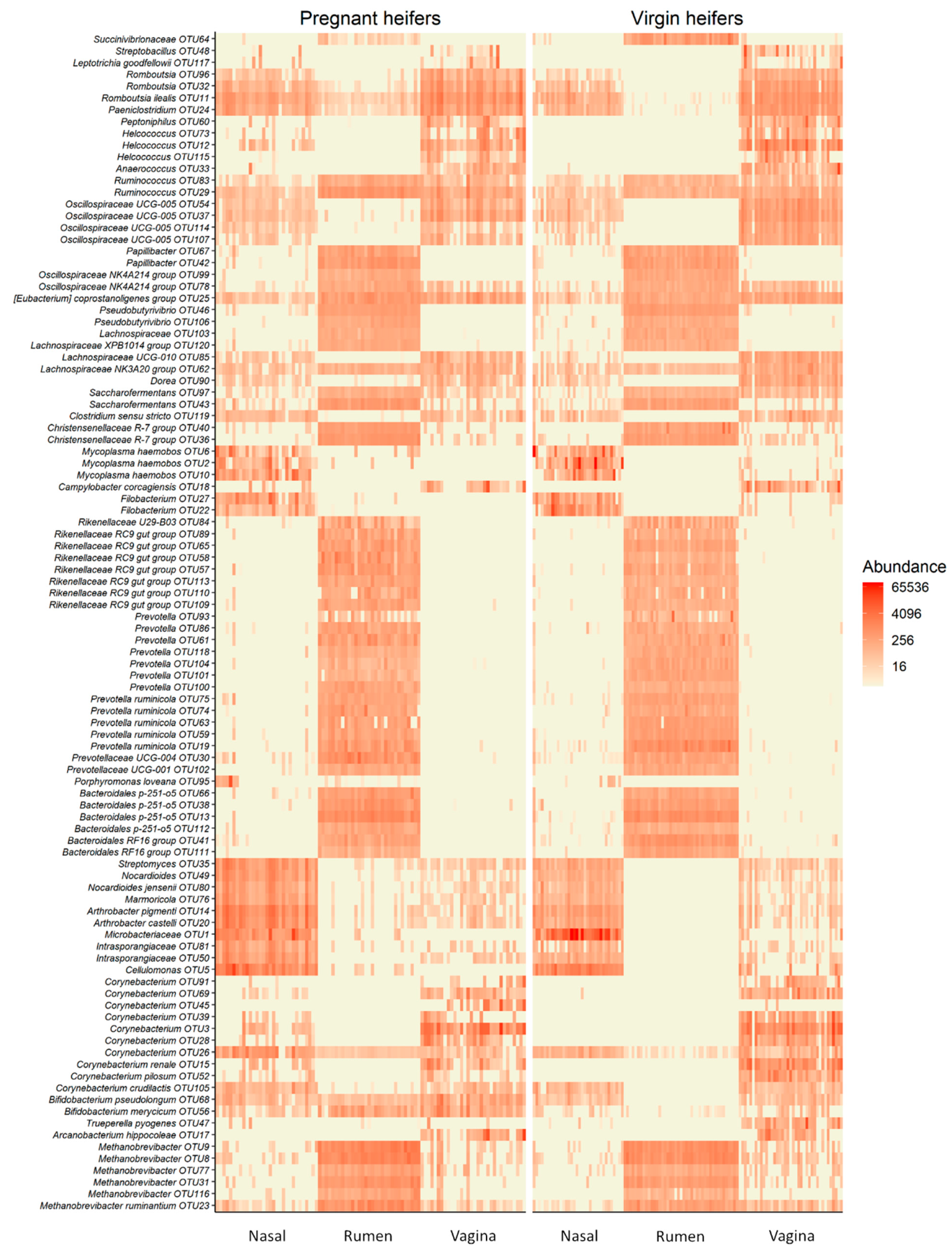
3.4. Holistic View of Nasopharyngeal, Ruminal and Vaginal Microbiota and Identification of Core Taxa Shared across These Microbiomes
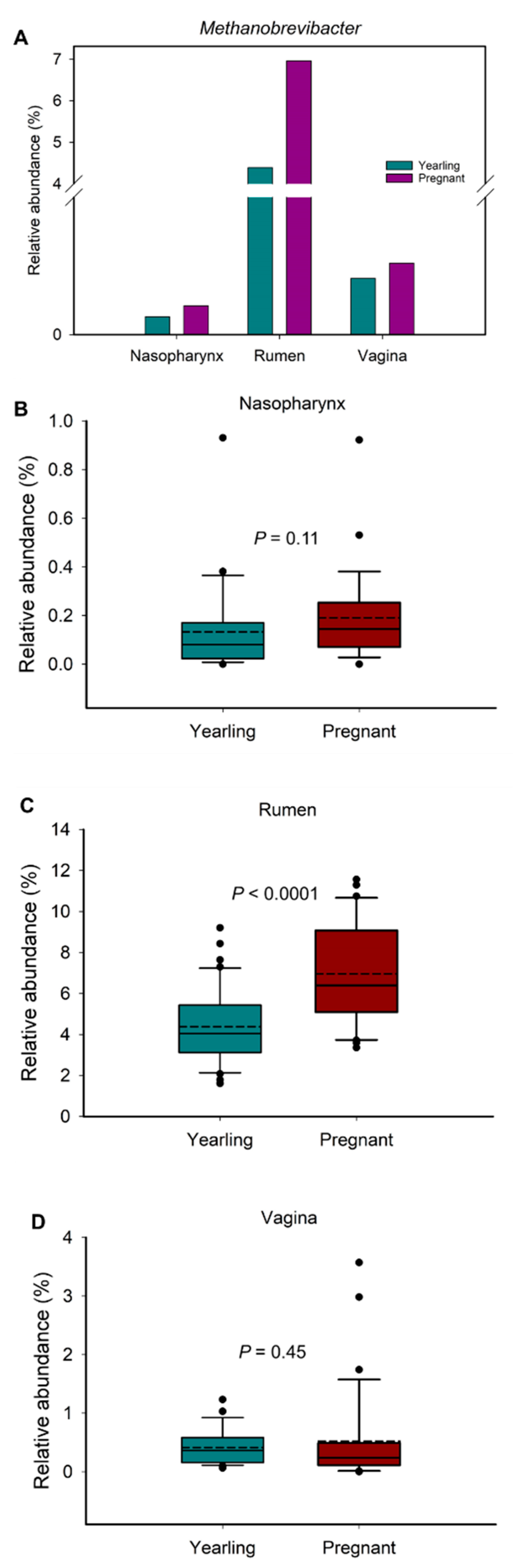
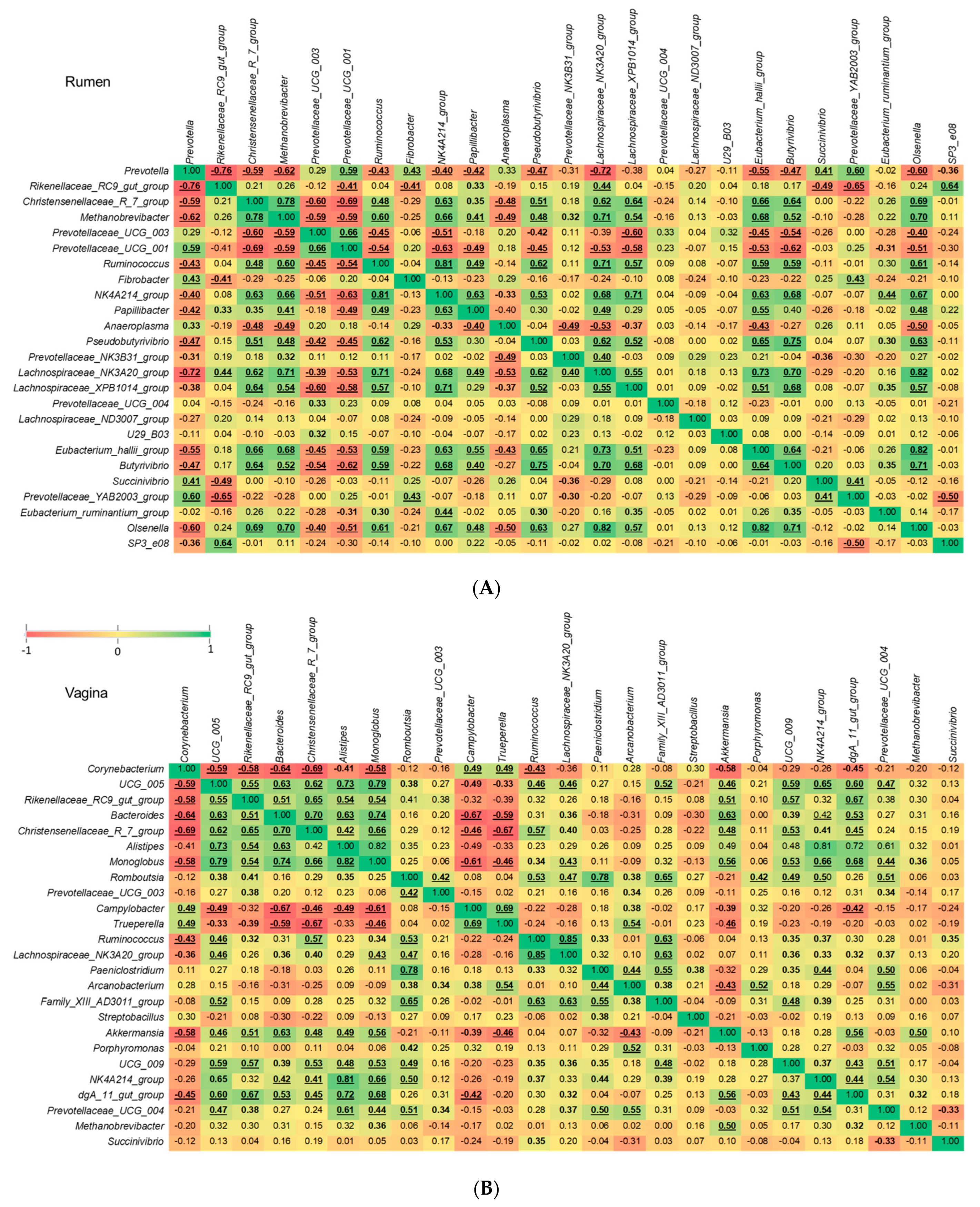
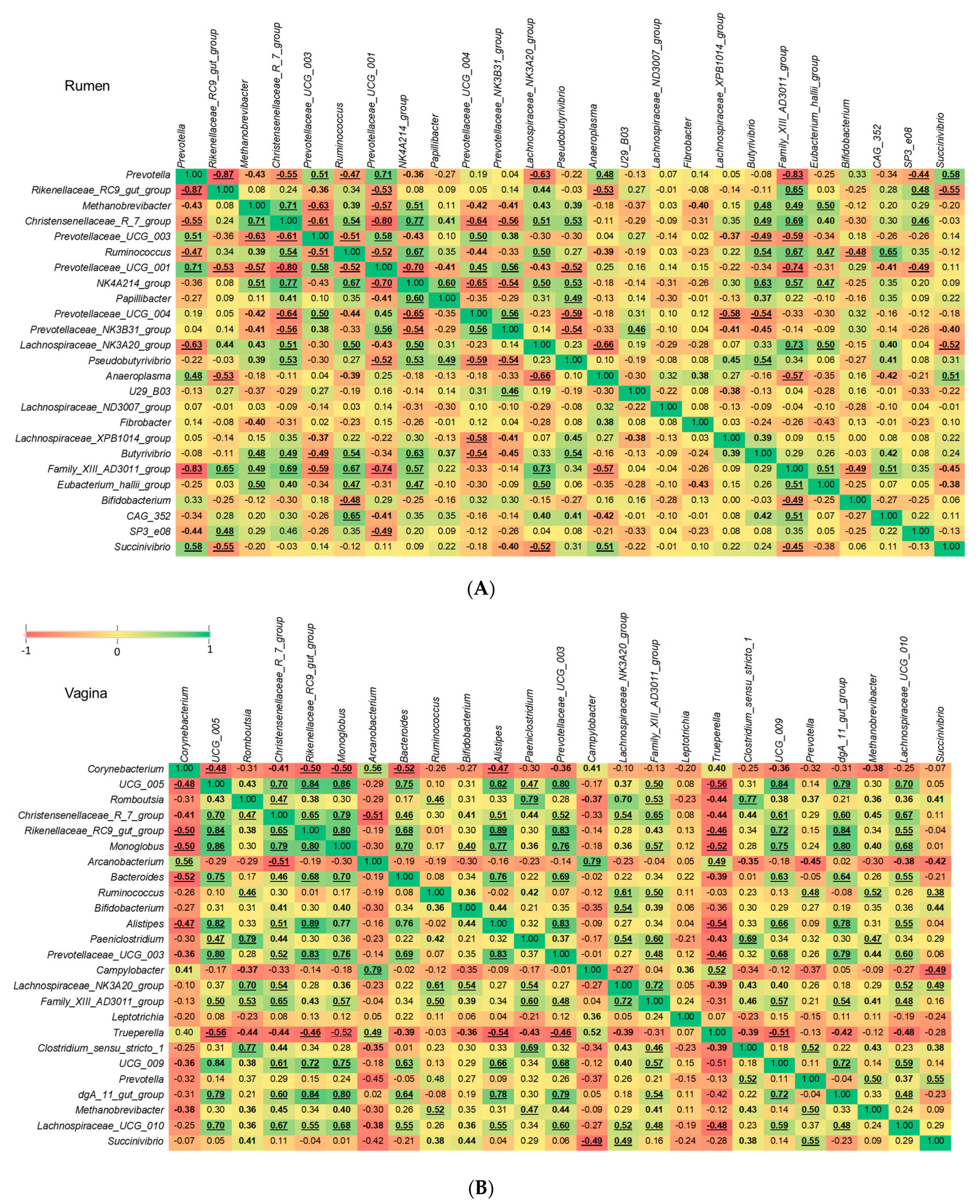
3.5. Comparison of Methanogenic Archaeal Relative Abundance between Virgin Yearling and Pregnant Heifers, and Identification of Bacterial Genera Associated with Methanobrevibacter
4. Discussion
4.1. Effect of Maternal Restricted Gain during the First Trimester of Gestation on Offspring Microbiota Development
4.2. Effect of Vitamin and Mineral Supplementation during the First 6 Months of Gestation on Maternal Microbiota
4.3. Holistic View of Microbial Communities across Respiratory, Gastrointestinal and Reproductive Tract and the Core Taxa Shared across These Habitats
4.4. Ruminal Methanobrevibacter Enriched in Pregnant Heifers and Associations of Methanobrevibacter with Predominant Ruminal and Vaginal Bacterial Genera
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huws, S.A.; Creevey, C.J.; Oyama, L.B.; Mizrahi, I.; Denman, S.E.; Popova, M.; Muñoz-Tamayo, R.; Forano, E.; Waters, S.M.; Hess, M.; et al. Addressing Global Ruminant Agricultural Challenges Through Understanding the Rumen Microbiome: Past, Present, and Future. Front. Microbiol. 2018, 9, 2161. [Google Scholar] [CrossRef]
- Matthews, C.; Crispie, F.; Lewis, E.; Reid, M.; O’Toole, P.W.; Cotter, P.D. The rumen microbiome: A crucial consideration when optimising milk and meat production and nitrogen utilisation efficiency. Gut Microbes 2018, 10, 115–132. [Google Scholar] [CrossRef]
- Galvão, K.N.; Bicalho, R.C.; Jeon, S.J. Symposium review: The uterine microbiome associated with the development of uterine disease in dairy cows. J. Dairy Sci. 2019, 102, 11786–11797. [Google Scholar] [CrossRef]
- Timsit, E.; McMullen, C.; Amat, S.; Alexander, T.W. Respiratory Bacterial Microbiota in Cattle. Veter- Clin. N. Am. Food Anim. Pr. 2020, 36, 297–320. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, E.; Neves, A.L.; Song, Y.; Guan, L.L. The Role of the Gut Microbiome in Cattle Production and Health: Driver or Passenger? Annu. Rev. Anim. Biosci. 2020, 8, 199–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Difford, G.F.; Plichta, D.R.; Løvendahl, P.; Lassen, J.; Noel, S.J.; Højberg, O.; Wright, A.-D.G.; Zhu, Z.; Kristensen, L.; Nielsen, H.B.; et al. Host genetics and the rumen microbiome jointly associate with methane emissions in dairy cows. PLoS Genet. 2018, 14, e1007580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Li, C.; Chen, Y.; Liu, J.; Zhang, C.; Irving, B.; Fitzsimmons, C.; Plastow, G.; Guan, L.L. Host genetics influence the rumen microbiota and heritable rumen microbial features associate with feed efficiency in cattle. Microbiome 2019, 7, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Weimer, P.J. Redundancy, resilience, and host specificity of the ruminal microbiota: Implications for engineering improved ruminal fermentations. Front. Microbiol. 2015, 6, 296. [Google Scholar] [CrossRef] [Green Version]
- Yáñez-Ruiz, D.R.; Abecia, L.; Newbold, C.J. Manipulating rumen microbiome and fermentation through interventions during early life: A review. Front. Microbiol. 2015, 6, 1133. [Google Scholar] [CrossRef] [Green Version]
- Saro, C.; Hohenester, U.M.; Bernard, M.; Lagrée, M.; Martin, C.; Doreau, M.; Boudra, H.; Popova, M.; Morgavi, D.P. Effectiveness of Interventions to Modulate the Rumen Microbiota Composition and Function in Pre-ruminant and Ruminant Lambs. Front. Microbiol. 2018, 9, 1273. [Google Scholar] [CrossRef]
- Belanche, A.; Palma-Hidalgo, J.; Nejjam, I.; Jiménez, E.; Martín-García, A.; Yáñez-Ruiz, D. Inoculation with rumen fluid in early life as a strategy to optimize the weaning process in intensive dairy goat systems. J. Dairy Sci. 2020, 103, 5047–5060. [Google Scholar] [CrossRef] [PubMed]
- Palma-Hidalgo, J.M.; Jiménez, E.; Popova, M.; Morgavi, D.P.; Martín-García, A.I.; Yáñez-Ruiz, D.R.; Belanche, A. Inoculation with rumen fluid in early life accelerates the rumen microbial development and favours the weaning process in goats. Anim. Microbiome 2021, 3, 11. [Google Scholar] [CrossRef] [PubMed]
- Guzman, C.E.; Wood, J.L.; Egidi, E.; White-Monsant, A.C.; Semenec, L.; Grommen, S.V.H.; Hill-Yardin, E.L.; De Groef, B.; Franks, A.E. A pioneer calf foetus microbiome. Sci. Rep. 2020, 10, 17712. [Google Scholar] [CrossRef] [PubMed]
- Amat, S.; Holman, D.B.; Schmidt, K.; McCarthy, K.L.; Dorsam, S.T.; Ward, A.K.; Borowicz, P.P.; Reynolds, L.P.; Caton, J.S.; Sedivec, K.K.; et al. Characterization of the microbiota associated with 12-week-old bovine fetuses exposed to divergent in utero nutrition. bioRxiv 2021. [Google Scholar] [CrossRef]
- Palmer, A.C. Nutritionally Mediated Programming of the Developing Immune System. Adv. Nutr. 2011, 2, 377–395. [Google Scholar] [CrossRef] [Green Version]
- Parlee, S.D.; MacDougald, O.A. Maternal nutrition and risk of obesity in offspring: The Trojan horse of developmental plasticity. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2013, 1842, 495–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanal, P.; Nielsen, M.O. Impacts of prenatal nutrition on animal production and performance: A focus on growth and metabolic and endocrine function in sheep. J. Anim. Sci. Biotechnol. 2017, 8, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caton, J.S.; Crouse, M.S.; Reynolds, L.P.; Neville, T.L.; Dahlen, C.R.; Ward, A.K.; Swanson, K.C. Maternal nutrition and programming of offspring energy requirements. Transl. Anim. Sci. 2019, 3, 976–990. [Google Scholar] [CrossRef] [Green Version]
- Stiemsma, L.T.; Michels, K.B. The Role of the Microbiome in the Developmental Origins of Health and Disease. Pediatrics 2018, 141, e20172437. [Google Scholar] [CrossRef] [Green Version]
- Calatayud, M.; Koren, O.; Collado, M.C. Maternal Microbiome and Metabolic Health Program Microbiome Development and Health of the Offspring. Trends Endocrinol. Metab. 2019, 30, 735–744. [Google Scholar] [CrossRef]
- Codagnone, M.; Spichak, S.; O’Mahony, S.M.; O’Leary, O.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Programming Bugs: Microbiota and the Developmental Origins of Brain Health and Disease. Biol. Psychiatry 2018, 85, 150–163. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Cai, X.; Chen, C.; Fang, H.; Zhao, Y.; Fei, W.; Chen, F.; Zheng, C. The Role of Microbiomes in Pregnant Women and Offspring: Research Progress of Recent Years. Front. Pharmacol. 2020, 11, 643. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; Miyamoto, J.; Ohue-Kitano, R.; Watanabe, K.; Yamada, T.; Onuki, M.; Aoki, R.; Isobe, Y.; Kashihara, D.; Inoue, D.; et al. Maternal gut microbiota in pregnancy influences offspring metabolic phenotype in mice. Science 2020, 367, eaaw8429. [Google Scholar] [CrossRef] [PubMed]
- Vuong, H.E.; Pronovost, G.N.; Williams, D.W.; Coley, E.J.L.; Siegler, E.L.; Qiu, A.; Kazantsev, M.; Wilson, C.J.; Rendon, T.; Hsiao, E.Y. The maternal microbiome modulates fetal neurodevelopment in mice. Nature 2020, 586, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Mee, J.F.; Rogers, A.P.; O’Farrell, K.J. Effect of feeding a mineral-vitamin supplement before calving on the calving performance of a trace element deficient dairy herd. Veter Rec. 1995, 137, 508–512. [Google Scholar] [CrossRef]
- Wilde, D. Influence of macro and micro minerals in the peri-parturient period on fertility in dairy cattle. Anim. Reprod. Sci. 2006, 96, 240–249. [Google Scholar] [CrossRef]
- Van Emon, M.; Sanford, C.; McCoski, S. Impacts of Bovine Trace Mineral Supplementation on Maternal and Offspring Production and Health. Animals 2020, 10, 2404. [Google Scholar] [CrossRef]
- Diniz, W.; Reynolds, L.; Borowicz, P.; Ward, A.; Sedivec, K.; McCarthy, K.; Kassetas, C.; Baumgaertner, F.; Kirsch, J.; Dorsam, S.; et al. Maternal Vitamin and Mineral Supplementation and Rate of Maternal Weight Gain Affects Placental Expression of Energy Metabolism and Transport-Related Genes. Genes 2021, 12, 385. [Google Scholar] [CrossRef]
- Menezes, A.C.B.; McCarthy, K.L.; Kassetas, C.J.; Baumgaertner, F.; Kirsch, J.D.; Dorsam, S.; Neville, T.L.; Ward, A.K.; Borowicz, P.P.; Reynolds, L.P.; et al. Vitamin and mineral supplementation and rate of gain during the first trimester of gestation affect concentrations of amino acids in maternal serum and allantoic fluid of beef heifers. J. Anim. Sci. 2021, 99, skab024. [Google Scholar] [CrossRef] [PubMed]
- NASEM. Nutrient Requirements of Beef Cattle, 8th ed.; The National Academies Press: Washington, DC, USA, 2016. [Google Scholar]
- Holman, D.B.; Timsit, E.; Amat, S.; Abbott, D.W.; Buret, A.G.; Alexander, T.W. The nasopharyngeal microbiota of beef cattle before and after transport to a feedlot. BMC Microbiol. 2017, 17, 70. [Google Scholar] [CrossRef] [Green Version]
- Amat, S.; Holman, D.; Timsit, E.; Schwinghamer, T.; Alexander, T.W. Evaluation of the Nasopharyngeal Microbiota in Beef Cattle Transported to a Feedlot, With a Focus on Lactic Acid-Producing Bacteria. Front. Microbiol. 2019, 10, 1988. [Google Scholar] [CrossRef]
- Paz, H.A.; Anderson, C.; Muller, M.J.; Kononoff, P.J.; Fernando, S.C. Rumen Bacterial Community Composition in Holstein and Jersey Cows Is Different under Same Dietary Condition and Is Not Affected by Sampling Method. Front. Microbiol. 2016, 7, 1206. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- McMurdie, P.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R. Package ‘Vegan’. Community Ecology Package, Version 2. 2013. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 15 September 2021).
- Mallick, H.; Rahnavard, A.; McIver, L.J.; Ma, S.; Zhang, Y.; Nguyen, L.H.; Tickle, T.L.; Weingart, G.; Ren, B.; Schwager, E.H. Multivariable association discovery in population-scale meta-omics studies. bioRxiv 2021. [Google Scholar] [CrossRef]
- Guzman, E.C.; Bereza-Malcolm, L.T.; De Groef, B.; Franks, A. Presence of Selected Methanogens, Fibrolytic Bacteria, and Proteobacteria in the Gastrointestinal Tract of Neonatal Dairy Calves from Birth to 72 Hours. PLoS ONE 2015, 10, e0133048. [Google Scholar] [CrossRef] [Green Version]
- Alipour, M.J.; Jalanka, J.; Pessa-Morikawa, T.; Kokkonen, T.; Satokari, R.; Hynönen, U.; Iivanainen, A.; Niku, M. The composition of the perinatal intestinal microbiota in cattle. Sci. Rep. 2018, 8, 10437. [Google Scholar] [CrossRef]
- McLean, K.J.; Crouse, M.; Crosswhite, M.R.; Pereira, N.N.; Dahlen, C.R.; Borowicz, P.P.; Reynolds, L.P.; Ward, A.K.; Neville, B.W.; Caton, J.S. Impacts of maternal nutrition on uterine and placental vascularity and mRNA expression of angiogenic factors during the establishment of pregnancy in beef heifers1. Transl. Anim. Sci. 2017, 1, 160–167. [Google Scholar] [CrossRef]
- Crouse, M.S.; Greseth, N.P.; McLean, K.J.; Crosswhite, M.R.; Pereira, N.N.; Ward, A.K.; Reynolds, L.P.; Dahlen, C.R.; Neville, B.W.; Borowicz, P.P.; et al. Maternal nutrition and stage of early pregnancy in beef heifers: Impacts on hexose and AA concentrations in maternal and fetal fluids1. J. Anim. Sci. 2019, 97, 1296–1316. [Google Scholar] [CrossRef] [PubMed]
- Baumgaertner, F.; Underdahl, S.R.; McCarthy, K.L.; Menezes, A.C.B.; Diniz, W.J.D.S.; Ward, A.K.; Sedivec, K.K.; Dorsam, S.T.; Kirsch, J.D.; Caton, J.; et al. 187 Effects of energy supplementation during early gestation in beef heifers on body weight, concentrations of IGF-1, and calf characteristics. J. Anim. Sci. 2020, 98, 163–164. [Google Scholar] [CrossRef]
- Yang, Q.; Liang, Q.; Balakrishnan, B.; Belobrajdic, D.P.; Feng, Q.-J.; Zhang, W. Role of Dietary Nutrients in the Modulation of Gut Microbiota: A Narrative Review. Nutrients 2020, 12, 381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslam, M.N.; Bassis, C.; Zhang, L.; Zaidi, S.; Varani, J.; Bergin, I.L. Calcium Reduces Liver Injury in Mice on a High-Fat Diet: Alterations in Microbial and Bile Acid Profiles. PLoS ONE 2016, 11, e0166178. [Google Scholar] [CrossRef]
- Trautvetter, U.; Camarinha-Silva, A.; Jahreis, G.; Lorkowski, S.; Glei, M. High phosphorus intake and gut-related parameters―results of a randomized placebo-controlled human intervention study. Nutr. J. 2018, 17, 23. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Tang, T.; Chang, X.; Fan, X.; Chen, X.; Wang, R.; Fan, C.; Qi, K. Abnormality in Maternal Dietary Calcium Intake During Pregnancy and Lactation Promotes Body Weight Gain by Affecting the Gut Microbiota in Mouse Offspring. Mol. Nutr. Food Res. 2018, 63, e1800399. [Google Scholar] [CrossRef]
- Neubauer, V.; Humer, E.; Mann, E.; Kröger, I.; Reisinger, N.; Wagner, M.; Zebeli, Q.; Petri, R.M. Effects of clay mineral supplementation on particle-associated and epimural microbiota, and gene expression in the rumen of cows fed high-concentrate diet. Anaerobe 2019, 59, 38–48. [Google Scholar] [CrossRef]
- Kelly, W.J.; Leahy, S.; Altermann, E.; Yeoman, C.J.; Dunne, J.C.; Kong, Z.; Pacheco, D.M.; Li, D.; Noel, S.J.; Moon, C.; et al. The Glycobiome of the Rumen Bacterium Butyrivibrio proteoclasticus B316T Highlights Adaptation to a Polysaccharide-Rich Environment. PLoS ONE 2010, 5, e11942. [Google Scholar] [CrossRef]
- Liu, C.; Li, X.H.; Chen, Y.X.; Cheng, Z.H.; Duan, Q.H.; Meng, Q.H.; Tao, X.P.; Shang, B.; Dong, H.M. Age-Related Response of Rumen Microbiota to Mineral Salt and Effects of Their Interactions on Enteric Methane Emissions in Cattle. Microb. Ecol. 2016, 73, 590–601. [Google Scholar] [CrossRef] [PubMed]
- Barden, M.; Richards-Rios, P.; Ganda, E.; Lenzi, L.; Eccles, R.; Neary, J.; Oultram, J.; Oikonomou, G. Maternal influences on oral and faecal microbiota maturation in neonatal calves in beef and dairy production systems. Anim. Microbiome 2020, 2, 31. [Google Scholar] [CrossRef]
- Lourenco, J.M.; Kieran, T.; Seidel, D.S.; Glenn, T.C.; Da Silveira, M.F.; Callaway, T.R.R.L.S., Jr. Comparison of the ruminal and fecal microbiotas in beef calves supplemented or not with concentrate. PLoS ONE 2020, 15, e0231533. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.A.; Isaiah, A.; Estill, C.T.; Pirelli, G.J.; Suchodolski, J.S. Weaned beef calves fed selenium-biofortified alfalfa hay have an enriched nasal microbiota compared with healthy controls. PLoS ONE 2017, 12, e0179215. [Google Scholar] [CrossRef]
- Hall, J.A.; Isaiah, A.; Bobe, G.; Estill, C.T.; Bishop-Stewart, J.K.; Davis, T.Z.; Suchodolski, J.S.; Pirelli, G.J. Feeding selenium-biofortified alfalfa hay during the preconditioning period improves growth, carcass weight, and nasal microbial diversity of beef calves. PLoS ONE 2020, 15, e0242771. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.K.; Pendell, D.L. Market Impacts of Reducing the Prevalence of Bovine Respiratory Disease in United States Beef Cattle Feedlots. Front. Veter-Sci. 2017, 4, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Xu, C.; Chen, X.; Cai, X.; Yang, S.; Sheng, Y.; Wang, T. Regulation of an antioxidant blend on intestinal redox status and major microbiota in early weaned piglets. Nutrition 2014, 30, 584–589. [Google Scholar] [CrossRef]
- Talsness, C.E.; Penders, J.; Jansen, E.H.J.M.; Damoiseaux, J.; Thijs, C.; Mommers, M. Influence of vitamin D on key bacterial taxa in infant microbiota in the KOALA Birth Cohort Study. PLoS ONE 2017, 12, e0188011. [Google Scholar] [CrossRef] [PubMed]
- Huda, M.N.; Ahmad, S.M.; Kalanetra, K.M.; Taft, D.; Alam, J.; Khanam, A.; Raqib, R.; Underwood, A.M.; Mills, A.; Stephensen, D.C.B. Neonatal Vitamin A Supplementation and Vitamin A Status Are Associated with Gut Microbiome Composition in Bangladeshi Infants in Early Infancy and at 2 Years of Age. J. Nutr. 2019, 149, 1075–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collado, M.C.; Isolauri, E.; Laitinen, K.; Salminen, S. Distinct composition of gut microbiota during pregnancy in overweight and normal-weight women. Am. J. Clin. Nutr. 2008, 88, 894–899. [Google Scholar] [CrossRef]
- Koren, O.; Goodrich, J.K.; Cullender, T.C.; Spor, A.; Laitinen, K.; Bäckhed, H.K.; Gonzalez, A.; Werner, J.J.; Angenent, L.; Knight, R.; et al. Host Remodeling of the Gut Microbiome and Metabolic Changes during Pregnancy. Cell 2012, 150, 470–480. [Google Scholar] [CrossRef] [Green Version]
- Nuriel-Ohayon, M.; Neuman, H.; Koren, O. Microbial Changes during Pregnancy, Birth, and Infancy. Front. Microbiol. 2016, 7, 1031. [Google Scholar] [CrossRef] [Green Version]
- Ricks, N.M.; Panzer, A.; McCoy, A.N.; Azcarate-Peril, M.A.; Keku, T.O.; Boggess, K.A.; Smid, M.C. Maternal Gut Microbiome Biodiversity in Pregnancy. Am. J. Perinatol. 2017, 35, 24–30. [Google Scholar] [CrossRef]
- Zeineldin, M.; Lowe, J.; Aldridge, B. Contribution of the Mucosal Microbiota to Bovine Respiratory Health. Trends Microbiol. 2019, 27, 753–770. [Google Scholar] [CrossRef] [PubMed]
- Cholewińska, P.; Górniak, W.; Wojnarowski, K. Impact of selected environmental factors on microbiome of the digestive tract of ruminants. BMC Vet. Res. 2021, 17, 25. [Google Scholar] [CrossRef] [PubMed]
- McMullen, C.; Alexander, T.W.; Léguillette, R.; Workentine, M.; Timsit, E. Topography of the respiratory tract bacterial microbiota in cattle. Microbiome 2020, 8, 91. [Google Scholar] [CrossRef] [PubMed]
- Derakhshani, H.; Fehr, K.B.; Sepehri, S.; Francoz, D.; De Buck, J.; Barkema, H.; Plaizier, J.C.; Khafipour, E. Invited review: Microbiota of the bovine udder: Contributing factors and potential implications for udder health and mastitis susceptibility. J. Dairy Sci. 2018, 101, 10605–10625. [Google Scholar] [CrossRef] [Green Version]
- Bartenslager, A.C.; Althuge, N.D.; Loy, J.D.; Hille, M.M.; Spangler, M.L.; Fernando, S.C. Longitudinal assessment of the bovine ocular bacterial community dynamics in calves. Anim. Microbiome 2021, 3, 16. [Google Scholar] [CrossRef]
- Zinicola, M.; Lima, F.; Lima, S.; Machado, V.; Gomez, M.; Döpfer, D.; Guard, C.; Bicalho, R. Altered Microbiomes in Bovine Digital Dermatitis Lesions, and the Gut as a Pathogen Reservoir. PLoS ONE 2015, 10, e0120504. [Google Scholar] [CrossRef]
- Lugli, G.A.; Duranti, S.; Albert, K.; Mancabelli, L.; Napoli, S.; Viappiani, A.; Anzalone, R.; Longhi, G.; Milani, C.; Turroni, F.; et al. Unveiling Genomic Diversity among Members of the Species Bifidobacterium pseudolongum, a Widely Distributed Gut Commensal of the Animal Kingdom. Appl. Environ. Microbiol. 2019, 85, e03065-18. [Google Scholar] [CrossRef] [Green Version]
- Abe, F.; Ishibashi, N.; Shimamura, S. Effect of administration of bifidobacteria and lactic acid bacteria to newborn calves and piglets. J. Dairy Sci. 1995, 78, 2838–2846. [Google Scholar] [CrossRef]
- Kissels, W.; Wu, X.; Santos, R. Short communication: Interaction of the isomers carvacrol and thymol with the antibiotics doxycycline and tilmicosin: In vitro effects against pathogenic bacteria commonly found in the respiratory tract of calves. J. Dairy Sci. 2017, 100, 970–974. [Google Scholar] [CrossRef] [Green Version]
- Hook, S.E.; Wright, A.-D.G.; McBride, B.W. Methanogens: Methane Producers of the Rumen and Mitigation Strategies. Archaea 2010, 2010, 945785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danielsson, R.; Dicksved, J.; Sun, L.; Gonda, H.; Müller, B.; Schnürer, A.; Bertilsson, J. Methane Production in Dairy Cows Correlates with Rumen Methanogenic and Bacterial Community Structure. Front. Microbiol. 2017, 8, 226. [Google Scholar] [CrossRef] [PubMed]
- Greening, C.; Geier, R.; Wang, C.; Woods, L.C.; Morales, S.E.; McDonald, M.J.; Rushton-Green, R.; Morgan, X.C.; Koike, S.; Leahy, S.C.; et al. Diverse hydrogen production and consumption pathways influence methane production in ruminants. ISME J. 2019, 13, 2617–2632. [Google Scholar] [CrossRef] [PubMed]
- Laguardia-Nascimento, M.; Branco, K.M.G.R.; Gasparini, M.R.; Giannattasio-Ferraz, S.; Leite, L.R.; Araújo, F.M.G.; Salim, A.C.D.M.; Nicoli, J.R.; De Oliveira, G.C.; Barbosa-Stancioli, E. Vaginal Microbiome Characterization of Nellore Cattle Using Metagenomic Analysis. PLoS ONE 2015, 10, e0143294. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Ungerfeld, E.; Eckard, R.J.; Wang, M. Review: Fifty years of research on rumen methanogenesis: Lessons learned and future challenges for mitigation. Animal 2020, 14, s2–s16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denman, S.E.; Fernandez, G.M.; Eshinkai, T.; Emitsumori, M.; McSweeney, C. Metagenomic analysis of the rumen microbial community following inhibition of methane formation by a halogenated methane analog. Front. Microbiol. 2015, 6, 1087. [Google Scholar] [CrossRef]
- Tapio, I.; Snelling, T.J.; Strozzi, F.; Wallace, R.J. The ruminal microbiome associated with methane emissions from ruminant livestock. J. Anim. Sci. Biotechnol. 2017, 8, 7. [Google Scholar] [CrossRef]
- Granja-Salcedo, Y.T.; Fernandes, R.M.; de Araujo, R.C.; Kishi, L.T.; Berchielli, T.T.; de Resende, F.D.; Berndt, A.; Siqueira, G.R. Long-Term Encapsulated Nitrate Supplementation Modulates Rumen Microbial Diversity and Rumen Fermentation to Reduce Methane Emission in Grazing Steers. Front. Microbiol. 2019, 10, 614. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet Composition, % DM | Dams of Virgin Yearling | Virgin Yearling | Pregnant Heifers | ||
|---|---|---|---|---|---|
| LG 1 | MG 2 | Control | VTM 3 | ||
| Corn silage | 37 | 29 | 20 | 30 | 30 |
| Prairie hay | 53 | 41 | 70 | 59 | 59 |
| Dried distillers grains plus solubles | 10 | 5 | 5 | 6 | 6 |
| Premix | − | − | 5 | 5 | 5 |
| Energy and protein supplement 1 | − | 25 | − | − | − |
| Nasopharynx | Rumen | Vagina | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genus | Rank | MG | LG | SEM | p-Value | Genus | Rank | MG | LG | SEM | p-Value | Genus | Rank | MG | LG | SEM | p-Value |
| Mycoplasma | 2 | 23.8 | 12.3 | 9.16 | 0.218 | Prevotella | 1 | 31.4 | 30.2 | 1.85 | 0.505 | Corynebacterium | 1 | 8.75 | 12.00 | 2.54 | 0.209 |
| Cellulomonas | 3 | 5.41 | 6.40 | 1.92 | 0.608 | Rikenellaceae RC9 gut group | 2 | 8.89 | 9.14 | 0.84 | 0.769 | Oscillospiraceae UCG-005 | 2 | 9.52 | 7.86 | 0.91 | 0.076 |
| Filobacterium | 4 | 2.62 | 8.03 | 3.00 | 0.081 | Christensenellaceae R-7 group | 4 | 4.70 | 4.81 | 0.48 | 0.804 | Rikenellaceae RC9 gut group | 4 | 4.30 | 3.87 | 0.51 | 0.406 |
| Arthrobacter | 5 | 2.36 | 2.05 | 0.51 | 0.546 | Methanobrevibacter | 5 | 4.30 | 4.47 | 0.55 | 0.756 | Bacteroides | 5 | 4.15 | 3.95 | 0.62 | 0.757 |
| Corynebacterium | 6 | 1.60 | 1.81 | 0.45 | 0.636 | Prevotellaceae UCG-003 | 7 | 3.82 | 3.77 | 0.19 | 0.803 | Christensenellaceae R-7 group | 8 | 3.25 | 3.22 | 0.37 | 0.952 |
| Nocardioides | 7 | 1.77 | 1.65 | 0.60 | 0.842 | Prevotellaceae UCG-001 | 8 | 2.77 | 2.85 | 0.19 | 0.660 | Alistipes | 9 | 3.55 | 2.55 | 0.40 | 0.017 |
| Oscillospiraceae UCG-005 | 9 | 1.35 | 1.36 | 0.58 | 0.975 | Ruminococcus | 9 | 2.55 | 2.36 | 0.18 | 0.305 | Monoglobus | 10 | 2.83 | 2.44 | 0.29 | 0.182 |
| Streptomyces | 13 | 1.06 | 0.76 | 0.34 | 0.389 | Fibrobacter | 12 | 1.36 | 1.45 | 0.30 | 0.773 | Romboutsia | 11 | 2.44 | 1.89 | 0.28 | 0.061 |
| Romboutsia | 15 | 0.85 | 0.80 | 0.22 | 0.809 | Oscillospiraceae NK4A214 group | 14 | 1.42 | 1.34 | 0.10 | 0.444 | Prevotellaceae UCG-003 | 13 | 1.71 | 1.79 | 0.29 | 0.771 |
| Christensenellaceae R-7 group | 16 | 0.83 | 0.71 | 0.31 | 0.702 | Papillibacter | 18 | 1.11 | 1.17 | 0.13 | 0.668 | Campylobacter | 16 | 1.24 | 1.69 | 1.03 | 0.668 |
| Ornithinimicrobium | 18 | 0.55 | 0.72 | 0.31 | 0.600 | Anaeroplasma | 21 | 0.96 | 0.90 | 0.10 | 0.551 | Trueperella | 17 | 0.66 | 2.30 | 1.14 | 0.160 |
| Rikenellaceae RC9 gut group | 19 | 0.49 | 0.56 | 0.18 | 0.697 | Pseudobutyrivibrio | 22 | 0.82 | 0.81 | 0.05 | 0.856 | Ruminococcus | 18 | 1.64 | 1.20 | 0.19 | 0.025 |
| Bacteroides | 21 | 0.52 | 0.49 | 0.16 | 0.865 | Prevotellaceae NK3B31 group | 26 | 0.69 | 0.79 | 0.08 | 0.193 | Lachnospiraceae NK3A20 group | 21 | 1.07 | 0.90 | 0.17 | 0.307 |
| Marmoricola | 22 | 0.58 | 0.41 | 0.13 | 0.187 | Lachnospiraceae NK3A20 group | 30 | 0.47 | 0.45 | 0.05 | 0.599 | Paeniclostridium | 22 | 1.13 | 0.79 | 0.19 | 0.092 |
| Ruminococcus | 24 | 0.43 | 0.50 | 0.18 | 0.658 | Lachnospiraceae XPB1014 group | 31 | 0.44 | 0.44 | 0.05 | 0.957 | Arcanobacterium | 24 | 0.49 | 1.35 | 0.50 | 0.090 |
| Lachnospiraceae NK3A20 group | 25 | 0.53 | 0.31 | 0.14 | 0.110 | Prevotellaceae UCG-004 | 32 | 0.44 | 0.41 | 0.05 | 0.547 | Family XIII AD3011 group | 25 | 0.97 | 0.81 | 0.10 | 0.116 |
| Prevotella | 26 | 0.58 | 0.24 | 0.26 | 0.196 | Lachnospiraceae ND3007 group | 33 | 0.39 | 0.44 | 0.06 | 0.429 | Streptobacillus | 26 | 1.58 | 0.07 | 1.00 | 0.139 |
| Ornithinicoccus | 29 | 0.33 | 0.42 | 0.13 | 0.527 | Rikenellaceae U29-B03 | 34 | 0.40 | 0.41 | 0.07 | 0.997 | Akkermansia | 27 | 0.85 | 0.79 | 0.20 | 0.754 |
| Clostridium sensu stricto 1 | 30 | 0.41 | 0.34 | 0.14 | 0.615 | [Eubacterium] hallii group | 36 | 0.40 | 0.38 | 0.03 | 0.573 | Porphyromonas | 28 | 0.34 | 1.23 | 1.01 | 0.382 |
| Olsenella | 31 | 0.32 | 0.39 | 0.14 | 0.618 | Butyrivibrio | 38 | 0.38 | 0.33 | 0.05 | 0.295 | Butyricicoccaceae UCG-009 | 30 | 0.74 | 0.66 | 0.09 | 0.395 |
| Bifidobacterium | 32 | 0.33 | 0.37 | 0.17 | 0.819 | Succinivibrio | 39 | 0.32 | 0.36 | 0.04 | 0.402 | Oscillospiraceae NK4A214 group | 31 | 0.81 | 0.56 | 0.09 | 0.010 |
| Paeniclostridium | 33 | 0.40 | 0.31 | 0.11 | 0.393 | Prevotellaceae YAB2003 group | 40 | 0.35 | 0.32 | 0.04 | 0.435 | Rikenellaceae dgA-11 gut group | 33 | 0.71 | 0.59 | 0.08 | 0.149 |
| Prevotellaceae UCG-003 | 35 | 0.43 | 0.22 | 0.13 | 0.111 | [Eubacterium] ruminantium group | 41 | 0.32 | 0.27 | 0.02 | 0.029 | Prevotellaceae UCG-004 | 37 | 0.68 | 0.54 | 0.11 | 0.209 |
| Monoglobus | 36 | 0.30 | 0.32 | 0.13 | 0.859 | Olsenella | 42 | 0.27 | 0.26 | 0.04 | 0.754 | Lachnospiraceae UCG-010 | 38 | 0.62 | 0.59 | 0.08 | 0.683 |
| Rhodococcus | 37 | 0.28 | 0.31 | 0.12 | 0.778 | Clostridium sensu stricto 1 | 39 | 0.54 | 0.68 | 0.21 | 0.514 | ||||||
| Brachybacterium | 38 | 0.31 | 0.27 | 0.12 | 0.777 | Olsenella | 40 | 0.30 | 0.92 | 0.55 | 0.263 | ||||||
| Mannheimia * | 0.01 | 0.00 | Alloprevotella | 42 | 0.57 | 0.42 | 0.13 | 0.246 | |||||||||
| Pasteurella * | 0.02 | 0.07 | Fusobacterium * | 0.01 | 0.02 | ||||||||||||
| Histophilus * | 0.00 | 0.01 | Trueperella * | 0.00 | 0.00 | Fusobacterium * | 0.00 | 0.00 | |||||||||
| Nasopharynx | Rumen | Vagina | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genus | Rank | CON | VTM | SEM | p-Value | Genus | Rank | CON | VTM | SEM | p-Value | Genus | Rank | CON | VTM | SEM | p-Value |
| Cellulomonas | 1 | 9.28 | 6.03 | 2.87 | 0.265 | Prevotella | 1 | 20.69 | 20.67 | 2.59 | 0.994 | Corynebacterium | 1 | 18.68 | 12.25 | 5.22 | 0.227 |
| Arthrobacter | 2 | 6.87 | 7.20 | 1.40 | 0.820 | Rikenellaceae RC9 gut group | 2 | 12.90 | 12.93 | 1.48 | 0.985 | Oscillospiraceae UCG-005 | 2 | 5.08 | 6.42 | 1.00 | 0.190 |
| Mycoplasma | 3 | 2.74 | 8.95 | 2.88 | 0.039 | Methanobrevibacter | 3 | 6.95 | 6.97 | 0.84 | 0.985 | Romboutsia | 4 | 4.39 | 3.62 | 0.77 | 0.330 |
| Corynebacterium | 5 | 5.23 | 4.26 | 1.01 | 0.342 | Christensenellaceae R-7 group | 5 | 5.62 | 5.98 | 0.38 | 0.352 | Christensenellaceae R-7 group | 5 | 3.16 | 3.86 | 0.60 | 0.244 |
| Nocardioides | 6 | 4.18 | 3.87 | 0.67 | 0.646 | Prevotellaceae UCG-003 | 7 | 3.22 | 3.00 | 0.23 | 0.348 | Rikenellaceae RC9 gut group | 7 | 3.09 | 2.97 | 0.75 | 0.875 |
| Filobacterium | 8 | 3.29 | 1.70 | 1.32 | 0.238 | Ruminococcus | 9 | 2.38 | 2.58 | 0.18 | 0.283 | Monoglobus | 8 | 2.51 | 3.08 | 0.56 | 0.316 |
| Streptomyces | 9 | 2.24 | 2.24 | 0.40 | 0.995 | Prevotellaceae UCG-001 | 10 | 2.52 | 2.28 | 0.29 | 0.414 | Arcanobacterium | 9 | 4.09 | 1.01 | 2.38 | 0.203 |
| Oscillospiraceae UCG-005 | 10 | 2.09 | 1.16 | 0.38 | 0.023 | Oscillospiraceae NK4A214 group | 11 | 1.41 | 1.73 | 0.13 | 0.024 | Bacteroides | 10 | 2.10 | 2.75 | 0.46 | 0.166 |
| Romboutsia | 11 | 1.56 | 1.30 | 0.19 | 0.194 | Papillibacter | 15 | 1.09 | 1.37 | 0.16 | 0.102 | Ruminococcus | 11 | 2.41 | 2.22 | 0.56 | 0.732 |
| Bacteroides | 12 | 1.45 | 1.20 | 0.54 | 0.644 | Prevotellaceae UCG-004 | 19 | 0.91 | 1.16 | 0.35 | 0.484 | Bifidobacterium | 13 | 2.27 | 1.93 | 0.54 | 0.538 |
| Porphyromonas | 13 | 1.65 | 0.77 | 1.29 | 0.498 | Prevotellaceae NK3B31 group | 21 | 1.06 | 0.95 | 0.16 | 0.515 | Alistipes | 14 | 1.89 | 2.06 | 0.45 | 0.709 |
| Marmoricola | 14 | 1.18 | 1.10 | 0.22 | 0.719 | Lachnospiraceae NK3A20 group | 23 | 0.86 | 0.84 | 0.10 | 0.841 | Paeniclostridium | 16 | 1.54 | 1.28 | 0.34 | 0.451 |
| Salinimicrobium | 16 | 0.72 | 1.48 | 0.40 | 0.068 | Pseudobutyrivibrio | 25 | 0.75 | 0.75 | 0.06 | 0.980 | Prevotellaceae UCG-003 | 17 | 1.52 | 1.20 | 0.42 | 0.460 |
| Brachybacterium | 19 | 1.08 | 0.93 | 0.21 | 0.485 | Anaeroplasma | 26 | 0.68 | 0.59 | 0.08 | 0.303 | Campylobacter | 18 | 2.17 | 0.42 | 1.43 | 0.231 |
| Ornithinicoccus | 21 | 0.92 | 0.88 | 0.17 | 0.835 | Rikenellaceae U29-B03 | 29 | 0.41 | 0.45 | 0.11 | 0.761 | Lachnospiraceae NK3A20 group | 20 | 0.98 | 1.20 | 0.28 | 0.451 |
| Rhodococcus | 22 | 0.84 | 0.78 | 0.16 | 0.683 | Lachnospiraceae ND3007 group | 30 | 0.46 | 0.40 | 0.09 | 0.520 | Family XIII AD3011 group | 21 | 0.95 | 1.09 | 0.19 | 0.477 |
| Rikenellaceae RC9 gut group | 24 | 0.86 | 0.59 | 0.17 | 0.114 | Fibrobacter | 31 | 0.38 | 0.46 | 0.08 | 0.296 | Leptotrichia | 26 | 0.01 | 1.70 | 1.20 | 0.171 |
| Christensenellaceae R-7 group | 25 | 0.84 | 0.56 | 0.12 | 0.033 | Lachnospiraceae XPB1014 group | 32 | 0.38 | 0.40 | 0.03 | 0.665 | Trueperella | 27 | 0.91 | 0.63 | 0.58 | 0.641 |
| Bifidobacterium | 26 | 0.53 | 0.81 | 0.19 | 0.146 | Butyrivibrio | 33 | 0.36 | 0.42 | 0.03 | 0.046 | Clostridium sensu stricto | 28 | 0.86 | 0.51 | 0.22 | 0.117 |
| Saccharopolyspora | 27 | 0.69 | 0.63 | 0.15 | 0.698 | Family XIII AD3011 group | 34 | 0.34 | 0.40 | 0.06 | 0.336 | Butyricicoccaceae UCG-009 | 30 | 0.54 | 0.73 | 0.13 | 0.178 |
| Alistipes | 28 | 0.71 | 0.61 | 0.16 | 0.510 | [Eubacterium] hallii group | 36 | 0.36 | 0.35 | 0.03 | 0.791 | Prevotella | 34 | 0.27 | 0.84 | 0.67 | 0.400 |
| Paeniclostridium | 29 | 0.75 | 0.55 | 0.10 | 0.061 | Bifidobacterium | 37 | 0.35 | 0.34 | 0.10 | 0.918 | Rikenellaceae dgA-11 gut group | 35 | 0.55 | 0.54 | 0.16 | 0.950 |
| Ruminococcus | 30 | 0.78 | 0.49 | 0.12 | 0.024 | Ruminococcaceae CAG-352 | 38 | 0.28 | 0.39 | 0.05 | 0.021 | Methanobrevibacter | 37 | 0.59 | 0.45 | 0.29 | 0.618 |
| Clostridium sensu stricto 1 | 31 | 0.58 | 0.58 | 0.15 | 0.998 | Rikenellaceae SP3-e08 | 40 | 0.29 | 0.34 | 0.08 | 0.537 | Lachnospiraceae UCG-010 | 38 | 0.43 | 0.56 | 0.08 | 0.114 |
| Lachnospiraceae NK3A20 group | 33 | 0.57 | 0.53 | 0.12 | 0.754 | Anaerovorax | 41 | 0.32 | 0.31 | 0.03 | 0.721 | Streptobacillus | 39 | 0.03 | 0.93 | 0.74 | 0.229 |
| Ornithinimicrobium | 34 | 0.65 | 0.44 | 0.10 | 0.049 | Monoglobus | 42 | 0.33 | 0.27 | 0.04 | 0.149 | Oscillospiraceae NK4A214 group | 41 | 0.47 | 0.48 | 0.10 | 0.932 |
| Altererythrobacter | 35 | 0.60 | 0.45 | 0.13 | 0.240 | Akkermansia | 42 | 0.43 | 0.51 | 0.19 | 0.667 | ||||||
| Microlunatus | 36 | 0.58 | 0.45 | 0.10 | 0.191 | ||||||||||||
| Monoglobus | 37 | 0.61 | 0.42 | 0.12 | 0.122 | ||||||||||||
| Mannheimia * | 0.01 | 0.02 | |||||||||||||||
| Pasteurella * | 0.00 | 0.00 | Fusobacterium * | 0.17 | 0.07 | ||||||||||||
| Histophilus * | 0.00 | 0.00 | Trueperella * | 0.00 | 0.00 | Fusobacterium * | 0.004 | 0.005 | |||||||||
| OTU | Taxa | 60% | 65% | 70% | 75% | 80% | 85% | 90% | 95% | 100% |
|---|---|---|---|---|---|---|---|---|---|---|
| OTU8 | [k__Archaea, p__Euryarchaeota, c__Methanobacteria, o__Methanobacteriales, f__Methanobacteriaceae, g__Methanobrevibacter, s__NA] | |||||||||
| OTU23 | [k__Archaea, p__Euryarchaeota, c__Methanobacteria, o__Methanobacteriales, f__Methanobacteriaceae, g__Methanobrevibacter, s__ruminantium] | |||||||||
| OTU56 | [k__Bacteria, p__Actinobacteriota, c__Actinobacteria, o__Bifidobacteriales, f__Bifidobacteriaceae, g__Bifidobacterium, s__merycicum] | |||||||||
| OTU68 | [k__Bacteria, p__Actinobacteriota, c__Actinobacteria, o__Bifidobacteriales, f__Bifidobacteriaceae, g__Bifidobacterium, s__pseudolongum] | |||||||||
| OTU105 | [k__Bacteria, p__Actinobacteriota, c__Actinobacteria, o__Corynebacteriales, f__Corynebacteriaceae, g__Corynebacterium, s__crudilactis] | |||||||||
| OTU147 | [k__Bacteria, p__Actinobacteriota, c__Actinobacteria, o__Corynebacteriales, f__Corynebacteriaceae, g__Corynebacterium, s__marinum] | |||||||||
| OTU26 | [k__Bacteria, p__Actinobacteriota, c__Actinobacteria, o__Corynebacteriales, f__Corynebacteriaceae, g__Corynebacterium, s__NA] | |||||||||
| OTU272 | [k__Bacteria, p__Actinobacteriota, c__Actinobacteria, o__Corynebacteriales, f__Corynebacteriaceae, g__Corynebacterium, s__provencense] | |||||||||
| OTU160 | [k__Bacteria, p__Actinobacteriota, c__Actinobacteria, o__Micrococcales, f__Intrasporangiaceae, g__Ornithinimicrobium, s__NA] | |||||||||
| OTU35 | [k__Bacteria, p__Actinobacteriota, c__Actinobacteria, o__Streptomycetales, f__Streptomycetaceae, g__Streptomyces, s__NA] | |||||||||
| OTU351 | [k__Bacteria, p__Actinobacteriota, c__Coriobacteriia, o__Coriobacteriales, f__Atopobiaceae, g__Atopobium, s__NA] | |||||||||
| OTU537 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Christensenellales, f__Christensenellaceae, g__Christensenellaceae R-7 group, s__NA] | |||||||||
| OTU370 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Clostridia UCG-014, f__NA, g__NA, s__NA] | |||||||||
| OTU927 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Clostridia UCG-014, f__NA, g__NA, s__NA] | |||||||||
| OTU43 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Hungateiclostridiaceae, f__Saccharofermentans, g__NA, s__NA] | |||||||||
| OTU97 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Hungateiclostridiaceae, f__Saccharofermentans, g__NA, s__NA] | |||||||||
| OTU133 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Lachnospirales, f__Lachnospiraceae, g__[Eubacterium] hallii group, s__NA] | |||||||||
| OTU158 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Lachnospirales, f__Lachnospiraceae, g__[Eubacterium] hallii group, s__NA] | |||||||||
| OTU489 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Lachnospirales, f__Lachnospiraceae, g__[Eubacterium] hallii group, s__NA] | |||||||||
| OTU62 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Lachnospirales, f__Lachnospiraceae, g__Lachnospiraceae NK3A20 group, s__NA] | |||||||||
| OTU373 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Lachnospirales, f__Lachnospiraceae, g__Lachnospiraceae NK3A20 group, s__NA] | |||||||||
| OTU657 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Lachnospirales, f__Lachnospiraceae, g__Lachnospiraceae NK3A20 group, s__NA] | |||||||||
| OTU1688 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Lachnospirales, f__Lachnospiraceae, g__Lachnospiraceae NK3A20 group, s__NA] | |||||||||
| OTU25 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Oscillospirales, f__[Eubacterium] coprostanoligenes group, g__NA, s__NA] | |||||||||
| OTU78 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Oscillospirales, f__Oscillospiraceae, g__NK4A214 group, s__NA] | |||||||||
| OTU37 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Oscillospirales, f__Oscillospiraceae, g__UCG-005, s__NA] | |||||||||
| OTU54 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Oscillospirales, f__Oscillospiraceae, g__UCG-005, s__NA] | |||||||||
| OTU360 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Oscillospirales, f__Ruminococcaceae, g__NA, s__NA] | |||||||||
| OTU29 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Oscillospirales, f__Ruminococcaceae, g__Ruminococcus, s__NA] | |||||||||
| OTU83 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Oscillospirales, f__Ruminococcaceae, g__Ruminococcus, s__NA] | |||||||||
| OTU188 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Oscillospirales, f__Ruminococcaceae, g__Ruminococcus, s__NA] | |||||||||
| OTU201 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Oscillospirales, f__Ruminococcaceae, g__Ruminococcus, s__NA] | |||||||||
| OTU307 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Oscillospirales, f__Ruminococcaceae, g__Ruminococcus, s__NA] | |||||||||
| OTU244 | k__Bacteria, p__Firmicutes, c__Clostridia, o__Oscillospirales, f__Ruminococcaceae, g__UCG-001, s__NA] | |||||||||
| OTU243 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Peptostreptococcales-Tissierellales, f__Anaerovoracaceae, g__Family XIII AD3011 group, s__NA] | |||||||||
| OTU518 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Peptostreptococcales-Tissierellales, f__Anaerovoracaceae, g__Family XIII AD3011 group, s__NA] | |||||||||
| OTU372 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Peptostreptococcales-Tissierellales, f__Anaerovoracaceae, g__Mogibacterium, s__NA] | |||||||||
| OTU24 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Peptostreptococcales-Tissierellales, f__Peptostreptococcaceae, g__Paeniclostridium, s__NA] | |||||||||
| OTU11 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Peptostreptococcales-Tissierellales, f__Peptostreptococcaceae, g__Romboutsia, s__ilealis] | |||||||||
| OTU32 | [k__Bacteria, p__Firmicutes, c__Clostridia, o__Peptostreptococcales-Tissierellales, f__Peptostreptococcaceae, g__Romboutsia, s__NA] | |||||||||
| OTU1655 | [k__Bacteria, p__Proteobacteria, c__Alphaproteobacteria, o__Acetobacterales, f__Acetobacteraceae, g__Acetobacter, s__pasteurianus] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amat, S.; Holman, D.B.; Schmidt, K.; Menezes, A.C.B.; Baumgaertner, F.; Winders, T.; Kirsch, J.D.; Liu, T.; Schwinghamer, T.D.; Sedivec, K.K.; et al. The Nasopharyngeal, Ruminal, and Vaginal Microbiota and the Core Taxa Shared across These Microbiomes in Virgin Yearling Heifers Exposed to Divergent In Utero Nutrition during Their First Trimester of Gestation and in Pregnant Beef Heifers in Response to Mineral Supplementation. Microorganisms 2021, 9, 2011. https://doi.org/10.3390/microorganisms9102011
Amat S, Holman DB, Schmidt K, Menezes ACB, Baumgaertner F, Winders T, Kirsch JD, Liu T, Schwinghamer TD, Sedivec KK, et al. The Nasopharyngeal, Ruminal, and Vaginal Microbiota and the Core Taxa Shared across These Microbiomes in Virgin Yearling Heifers Exposed to Divergent In Utero Nutrition during Their First Trimester of Gestation and in Pregnant Beef Heifers in Response to Mineral Supplementation. Microorganisms. 2021; 9(10):2011. https://doi.org/10.3390/microorganisms9102011
Chicago/Turabian StyleAmat, Samat, Devin B. Holman, Kaycie Schmidt, Ana Clara B. Menezes, Friederike Baumgaertner, Thomas Winders, James D. Kirsch, Tingting Liu, Timothy D. Schwinghamer, Kevin K. Sedivec, and et al. 2021. "The Nasopharyngeal, Ruminal, and Vaginal Microbiota and the Core Taxa Shared across These Microbiomes in Virgin Yearling Heifers Exposed to Divergent In Utero Nutrition during Their First Trimester of Gestation and in Pregnant Beef Heifers in Response to Mineral Supplementation" Microorganisms 9, no. 10: 2011. https://doi.org/10.3390/microorganisms9102011
APA StyleAmat, S., Holman, D. B., Schmidt, K., Menezes, A. C. B., Baumgaertner, F., Winders, T., Kirsch, J. D., Liu, T., Schwinghamer, T. D., Sedivec, K. K., & Dahlen, C. R. (2021). The Nasopharyngeal, Ruminal, and Vaginal Microbiota and the Core Taxa Shared across These Microbiomes in Virgin Yearling Heifers Exposed to Divergent In Utero Nutrition during Their First Trimester of Gestation and in Pregnant Beef Heifers in Response to Mineral Supplementation. Microorganisms, 9(10), 2011. https://doi.org/10.3390/microorganisms9102011