
Molecular Characterization of Fungal Biodiversity in Long-Term Polychlorinated Biphenyl-Contaminated Soils

Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sampling
2.2. Soils’ Chemical Properties
2.3. DNA Extraction, PCR Amplification and Illumina-Based Sequencing
2.4. Bioinformatic Analysis
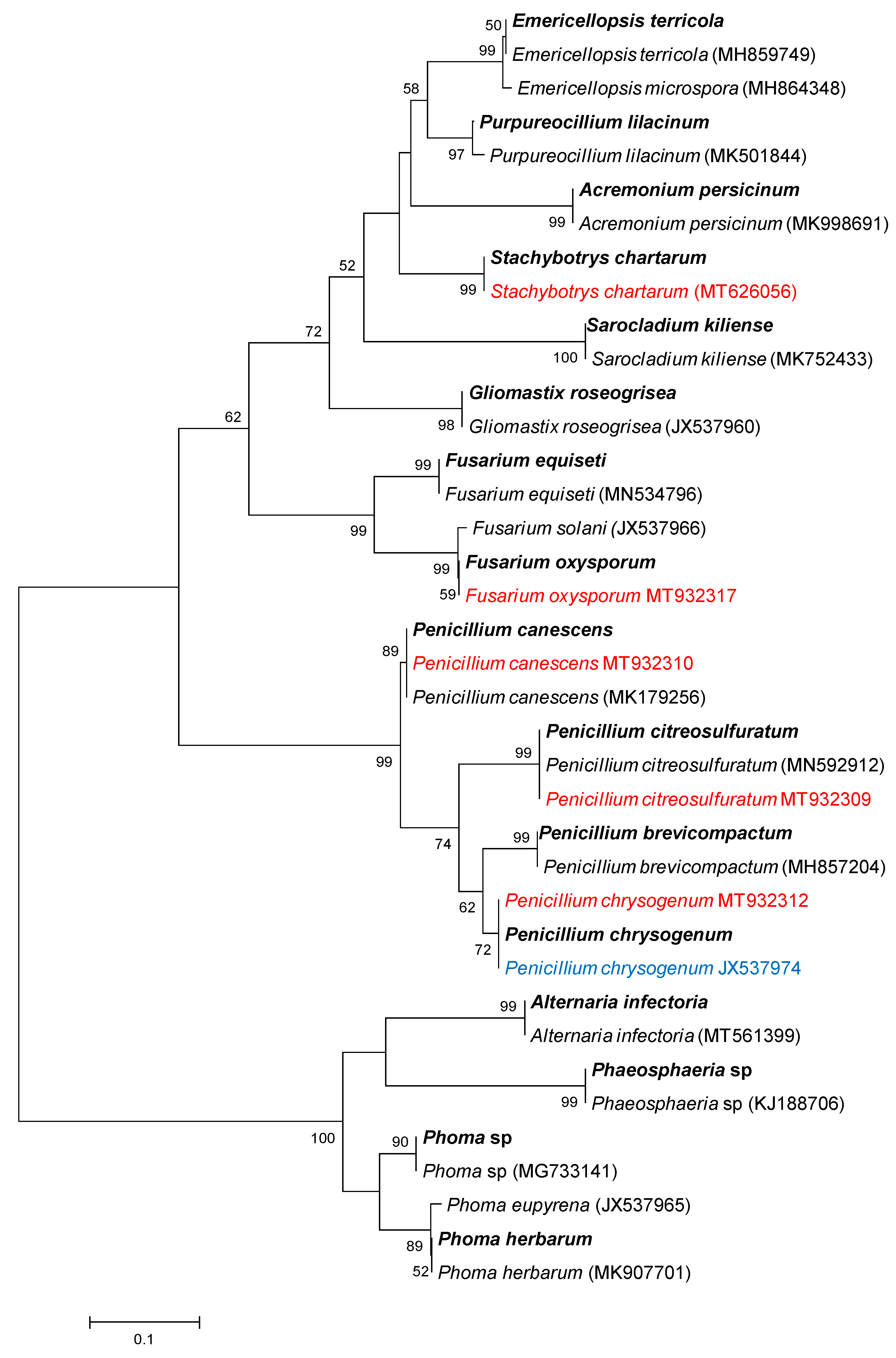
2.5. Phylogenetic Analysis
2.6. Statistical Analysis
3. Results
3.1. Alpha Diversity of Fungal Communities
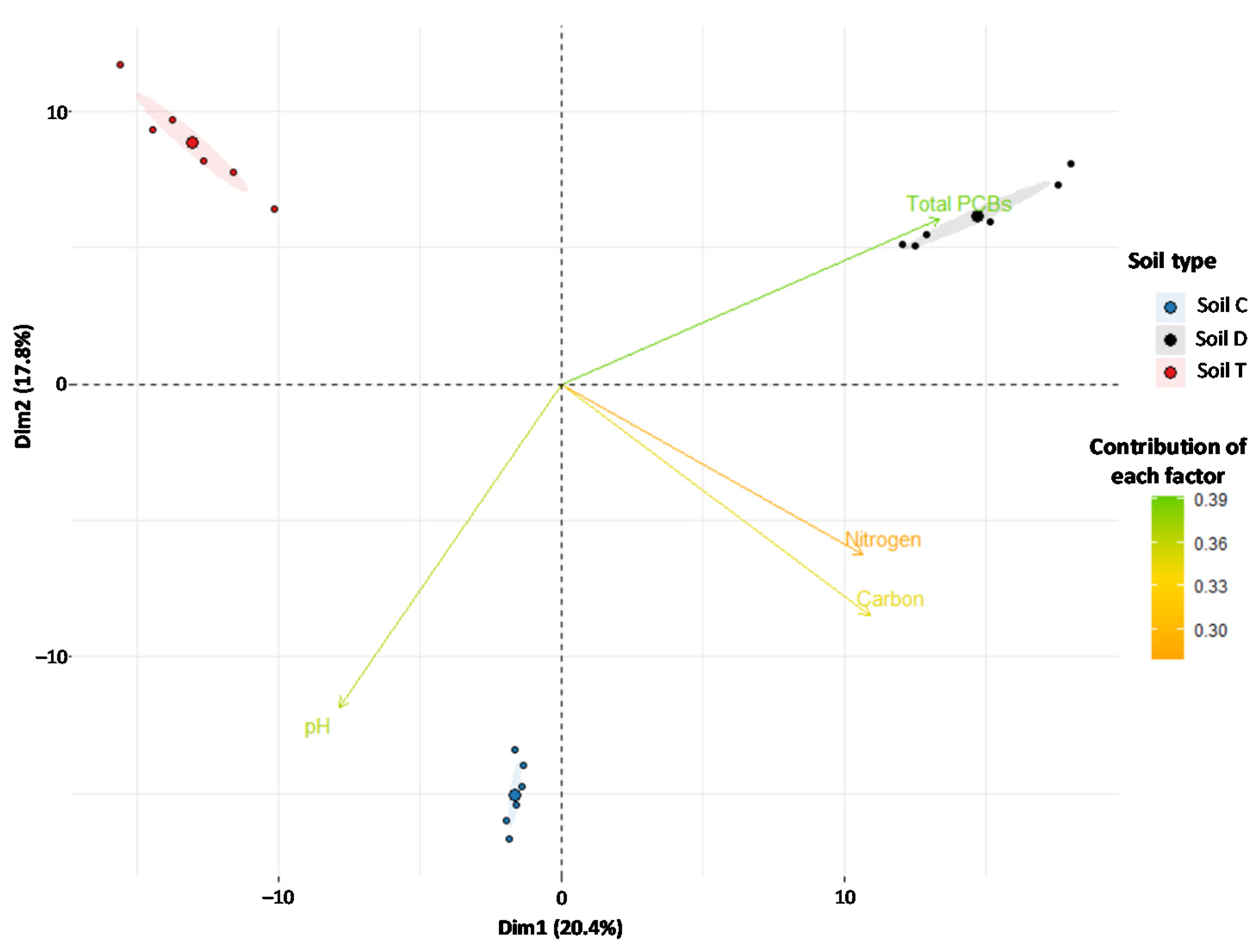
3.2. Patterns of Soil Fungal Communities
3.3. Composition of Soils’ Fungal Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Edwards, C.A. Assessing the Effects of Environmental Pollutants on Soil Organisms, Communities, Processes and Ecosystems. Eur. J. Soil Biol. 2002, 38, 225–231. [Google Scholar] [CrossRef]
- Kowalchuk, G.A.; Stephen, J.R. Ammonia-Oxidizing Bacteria: A Model for Molecular Microbial Ecology. Annu. Rev. Microbiol. 2001, 55, 485–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Heijden, M.G.A.; Bardgett, R.D.; van Straalen, N.M. The Unseen Majority: Soil Microbes as Drivers of Plant Diversity and Productivity in Terrestrial Ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Wang, P.; Devlin, A.T.; Xiao, S.; Shu, W.; Zhang, H.; Ding, M. Influence of Soil and Water Conservation Measures on Soil Microbial Communities in a Citrus Orchard of Southeast China. Microorganisms 2021, 9, 319. [Google Scholar] [CrossRef] [PubMed]
- Shuaib, M.; Azam, N.; Bahadur, S.; Romman, M.; Yu, Q.; Xuexiu, C. Variation and Succession of Microbial Communities under the Conditions of Persistent Heavy Metal and Their Survival Mechanism. Microb. Pathog. 2021, 150, 104713. [Google Scholar] [CrossRef]
- Shaoping, K.; Zhiwei, D.; Bingchen, W.; Huihui, W.; Jialiang, L.; Hongbo, S. Changes of Sensitive Microbial Community in Oil Polluted Soil in the Coastal Area in Shandong, China for Ecorestoration. Ecotoxicol. Environ. Saf. 2021, 207, 111551. [Google Scholar] [CrossRef]
- Chun, S.C.; Muthu, M.; Hasan, N.; Tasneem, S.; Gopal, J. Mycoremediation of PCBs by Pleurotus Ostreatus: Possibilities and Prospects. Appl. Sci. 2019, 9, 4185. [Google Scholar] [CrossRef] [Green Version]
- Passatore, L.; Rossetti, S.; Juwarkar, A.A.; Massacci, A. Phytoremediation and Bioremediation of Polychlorinated Biphenyls (PCBs): State of Knowledge and Research Perspectives. J. Hazard. Mater. 2014, 278, 189–202. [Google Scholar] [CrossRef]
- Helmfrid, I.; Berglund, M.; Löfman, O.; Wingren, G. Health Effects and Exposure to Polychlorinated Biphenyls (PCBs) and Metals in a Contaminated Community. Environ. Int. 2012, 44, 53–58. [Google Scholar] [CrossRef]
- Lauby-Secretan, B.; Loomis, D.; Grosse, Y.; Ghissassi, F.E.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Baan, R.; Mattock, H.; Straif, K. Carcinogenicity of Polychlorinated Biphenyls and Polybrominated Biphenyls. Lancet Oncol. 2013, 14, 287–288. [Google Scholar] [CrossRef] [Green Version]
- El Majidi, N.; Bouchard, M.; Carrier, G. Systematic Analysis of the Relationship between Standardized Prenatal Exposure to Polychlorinated Biphenyls and Mental and Motor Development during Follow-up of Nine Children Cohorts. Regul. Toxicol. Pharmacol. 2013, 66, 130–146. [Google Scholar] [CrossRef]
- Dave, P.N.; Chaturvedi, S.; Sahu, L.K. Impact of polychlorinated biphenyls on environment and public health. In Handbook of Advanced Approaches towards Pollution Prevention and Control; Rahman, R.O.A., Hussain, C.M., Eds.; Elsevier: Netherlands, Amsterdam, 2021; Volume 1, pp. 261–280. ISBN 978-0-12-822121-1. [Google Scholar]
- Harms, H.; Schlosser, D.; Wick, L.Y. Untapped Potential: Exploiting Fungi in Bioremediation of Hazardous Chemicals. Nat. Rev. Microbiol. 2011, 9, 177–192. [Google Scholar] [CrossRef]
- Sharma, J.K.; Gautam, R.K.; Nanekar, S.V.; Weber, R.; Singh, B.K.; Singh, S.K.; Juwarkar, A.A. Advances and Perspective in Bioremediation of Polychlorinated Biphenyl-Contaminated Soils. Environ. Sci. Pollut. Res. 2018, 25, 16355–16375. [Google Scholar] [CrossRef]
- Horváthová, H.; Lászlová, K.; Dercová, K. Bioremediation of PCB-Contaminated Shallow River Sediments: The Efficacy of Biodegradation Using Individual Bacterial Strains and Their Consortia. Chemosphere 2018, 193, 270–277. [Google Scholar] [CrossRef]
- Stella, T.; Covino, S.; Čvančarová, M.; Filipová, A.; Petruccioli, M.; D’Annibale, A.; Cajthaml, T. Bioremediation of Long-Term PCB-Contaminated Soil by White-Rot Fungi. J. Hazard. Mater. 2017, 324, 701–710. [Google Scholar] [CrossRef]
- Sadañoski, M.A.; Benítez, S.F.; Fonseca, M.I.; Velázquez, J.E.; Zapata, P.D.; Levin, L.N.; Villalba, L.L. Mycoremediation of High Concentrations of Polychlorinated Biphenyls with Pleurotus Sajor-Caju LBM 105 as an Effective and Cheap Treatment. J. Environ. Chem. Eng. 2019, 7, 103453. [Google Scholar] [CrossRef]
- Sadañoski, M.A.; Benitez, S.F.; Velázquez, J.E.; Fonseca, M.I.; Zapata, P.D.; Levin, L.N.; Villalba, L.L. Bioprocess Conditions for Treating Mineral Transformer Oils Contaminated with Polychlorinated Biphenyls (PCBs). J. Environ. Chem. Eng. 2020, 8, 104068. [Google Scholar] [CrossRef]
- Tigini, V.; Prigione, V.; Di Toro, S.; Fava, F.; Varese, G.C. Isolation and Characterisation of Polychlorinated Biphenyl (PCB) Degrading Fungi from a Historically Contaminated Soil. Microb. Cell Factories 2009, 8, 5. [Google Scholar] [CrossRef] [Green Version]
- Mouhamadou, B.; Faure, M.; Sage, L.; Marçais, J.; Souard, F.; Geremia, R.A. Potential of Autochthonous Fungal Strains Isolated from Contaminated Soils for Degradation of Polychlorinated Biphenyls. Fungal Biol. 2013, 117, 268–274. [Google Scholar] [CrossRef]
- Sage, L.; Périgon, S.; Faure, M.; Gaignaire, C.; Abdelghafour, M.; Mehu, J.; Geremia, R.A.; Mouhamadou, B. Autochthonous Ascomycetes in Depollution of Polychlorinated Biphenyls Contaminated Soil and Sediment. Chemosphere 2014, 110, 62–69. [Google Scholar] [CrossRef]
- Germain, J.; Raveton, M.; Binet, M.N.; Mouhamadou, B. Screening and Metabolic Potential of Fungal Strains Isolated from Contaminated Soil and Sediment in the Polychlorinated Biphenyl Degradation. Ecotoxicol. Environ. Saf. 2021, 208, 111703. [Google Scholar] [CrossRef]
- Germain, J.; Raveton, M.; Binet, M.-N.; Mouhamadou, B. Potentiality of Native Ascomycete Strains in Bioremediation of Highly Polychlorinated Biphenyl Contaminated Soils. Microorganisms 2021, 9, 612. [Google Scholar] [CrossRef]
- Baldrian, P. Wood-Inhabiting Ligninolytic Basidiomycetes in Soils: Ecology and Constraints for Applicability in Bioremediation. Fungal Ecol. 2008, 1, 4–12. [Google Scholar] [CrossRef]
- Nogales, B.; Moore, E.R.B.; Llobet-Brossa, E.; Rossello-Mora, R.; Amann, R.; Timmis, K.N. Combined Use of 16S Ribosomal DNA and 16S RRNA To Study the Bacterial Community of Polychlorinated Biphenyl-Polluted Soil. Appl. Environ. Microbiol. 2001, 67, 1874–1884. [Google Scholar] [CrossRef] [Green Version]
- Petrić, I.; Bru, D.; Udiković-Kolić, N.; Hršak, D.; Philippot, L.; Martin-Laurent, F. Evidence for Shifts in the Structure and Abundance of the Microbial Community in a Long-Term PCB-Contaminated Soil under Bioremediation. J. Hazard. Mater. 2011, 195, 254–260. [Google Scholar] [CrossRef]
- Du, J.; Hou, F.; Zhou, Q. Response of Soil Enzyme Activity and Soil Bacterial Community to PCB Dissipation across Different Soils. Chemosphere 2021, 283, 131229. [Google Scholar] [CrossRef]
- Ding, N.; Hayat, T.; Wang, J.; Wang, H.; Liu, X.; Xu, J. Responses of Microbial Community in Rhizosphere Soils When Ryegrass Was Subjected to Stress from PCBs. J. Soils Sediments 2011, 11, 1355–1362. [Google Scholar] [CrossRef]
- Robertson, G.P.; Coleman, D.C.; Bledsoe, C.S.; Sollins, P. Standard Soil Methods for Long-Term Ecological Research; Long-Term Ecological Research Network Series; Oxford University Press: New York, NY, USA, 1999; ISBN 978-0-19-512083-7. [Google Scholar]
- Piton, G.; Legay, N.; Arnoldi, C.; Lavorel, S.; Clément, J.-C.; Foulquier, A. Using Proxies of Microbial Community-weighted Means Traits to Explain the Cascading Effect of Management Intensity, Soil and Plant Traits on Ecosystem Resilience in Mountain Grasslands. J. Ecol. 2020, 108, 876–893. [Google Scholar] [CrossRef]
- AFNOR Groupe. Available online: https://www.boutique.afnor.org/norme/nf-en-16167/sols-biodechets-traites-et-boues-dosage-des-polychlorobiphenyles-pcbs-par-chromatographie-en-phase-gazeuse-spectrometrie-gazeuse/article/906803/fa190960 (accessed on 11 January 2021).
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. AMPLIFICATION AND DIRECT SEQUENCING OF FUNGAL RIBOSOMAL RNA GENES FOR PHYLOGENETICS. In PCR Protocols; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc: San Diego, CA, USA, 1990; pp. 315–322. ISBN 978-0-12-372180-8. [Google Scholar]
- Epp, L.S.; Boessenkool, S.; Bellemain, E.P.; Haile, J.; Esposito, A.; Riaz, T.; Erséus, C.; Gusarov, V.I.; Edwards, M.E.; Johnsen, A.; et al. New environmental metabarcodes for analysing soil DNA: Potential for Studying Past and Present Ecosystems. Mol. Ecol. 2012, 21, 1821–1833. [Google Scholar] [CrossRef]
- Mouhamadou, B.; Puissant, J.; Personeni, E.; Desclos-Theveniau, M.; Kastl, E.M.; Schloter, M.; Zinger, L.; Roy, J.; Geremia, R.A.; Lavorel, S. Effects of Two Grass Species on the Composition of Soil Fungal Communities. Biol. Fertil. Soils 2013, 49, 1131–1139. [Google Scholar] [CrossRef]
- Boyer, F.; Mercier, C.; Bonin, A.; Le Bras, Y.; Taberlet, P.; Coissac, E. OBITOOLS: A UNIX-inspired software package for DNA metabarcoding. Mol. Ecol. Resour. 2016, 16, 176–182. [Google Scholar] [CrossRef]
- van Dongen, S. Graph Clustering by Flow Simulation, Center for Math and Computer Science. Ph.D. Thesis, Utrecht University, Amsterdam, The Netherlands, 2000. [Google Scholar]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Thompson, L.A. R (and S-PLUS) Manual to Accompany Agresti’s Categorical Data Analysis (2002) 2nd Edition. Available online: http://users.stat.ufl.edu/~aa/cda/Thompson_manual.pdf (accessed on 11 January 2021).
- Vegan: Community Ecology Package; R Package Version 1.15-4; Ordination methods, diversity analysis and other functions for community and vegetation ecologists; The R Project for Statistical Computing: Vienna, Austria, 2009.
- Ventorino, V.; Pascale, A.; Adamo, P.; Rocco, C.; Fiorentino, N.; Mori, M.; Faraco, V.; Pepe, O.; Fagnano, M. Comparative Assessment of Autochthonous Bacterial and Fungal Communities and Microbial Biomarkers of Polluted Agricultural Soils of the Terra Dei Fuochi. Sci. Rep. 2018, 8, 14281. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Axelsson, E.P.; Girona, M.M.; Senior, J.K. Continuous-Cover Forestry Maintains Soil Fungal Communities in Norway Spruce Dominated Boreal Forests. For. Ecol. Manag. 2021, 480, 118659. [Google Scholar] [CrossRef]
- Oehl, F.; Laczko, E.; Bogenrieder, A.; Stahr, K.; Bösch, R.; van der Heijden, M.; Sieverding, E. Soil Type and Land Use Intensity Determine the Composition of Arbuscular Mycorrhizal Fungal Communities. Soil Biol. Biochem. 2010, 42, 724–738. [Google Scholar] [CrossRef]
- Wubet, T.; Christ, S.; Schöning, I.; Boch, S.; Gawlich, M.; Schnabel, B.; Fischer, M.; Buscot, F. Differences in soil fungal communities between european beech (Fagus sylvatica L.) dominated forests are related to soil and understory vegetation. PLoS ONE 2012, 7, e47500. [Google Scholar] [CrossRef]
- Nagati, M.; Roy, M.; Manzi, S.; Richard, F.; Desrochers, A.; Gardes, M.; Bergeron, Y. Impact of Local Forest Composition on Soil Fungal Communities in a Mixed Boreal Forest. Plant Soil 2018, 432, 345–357. [Google Scholar] [CrossRef]
- Zeng, Q.; Liu, Y.; Xiao, L.; An, S. Climate and Soil Properties Regulate Soil Fungal Communities on the Loess Plateau. Pedobiologia 2020, 81–82, 150668. [Google Scholar] [CrossRef]
- Périgon, S.; Massier, M.; Germain, J.; Binet, M.-N.; Legay, N.; Mouhamadou, B. Metabolic Adaptation of Fungal Strains in Response to Contamination by Polychlorinated Biphenyls. Environ. Sci. Pollut. Res. 2019, 26, 14943–14950. [Google Scholar] [CrossRef]
- Bourceret, A.; Cébron, A.; Tisserant, E.; Poupin, P.; Bauda, P.; Beguiristain, T.; Leyval, C. The Bacterial and Fungal Diversity of an Aged PAH- and Heavy Metal-Contaminated Soil Is Affected by Plant Cover and Edaphic Parameters. Microb. Ecol. 2016, 71, 711–724. [Google Scholar] [CrossRef] [PubMed]
- Egidi, E.; Delgado-Baquerizo, M.; Plett, J.M.; Wang, J.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K. A Few Ascomycota Taxa Dominate Soil Fungal Communities Worldwide. Nat. Commun. 2019, 10, 2369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stajich, J.E.; Berbee, M.L.; Blackwell, M.; Hibbett, D.S.; James, T.Y.; Spatafora, J.W.; Taylor, J.W. The Fungi. Curr. Biol. 2009, 19, R840–R845. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Soil Properties | Soil PCB Concentrations | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| pH | Organic Matter (g kg−1) | Nitrogen (g kg−1) | Carbon (g kg−1) | PCB28 (mg kg−1) | PCB52 (mg kg−1) | PCB101 (mg kg−1) | PCB118 (mg kg−1) | PCB138 (mg kg−1) | PCB153 (mg kg−1) | PCB180 (mg kg−1) | Total PCB (mg kg−1) | |
| Soil C | 9.16 ± 0.02 | 42.25 ± 1.00 | 109.75 ± 5.09 | 57.08 ± 0.77 | 0.03 ± 0.01 | 0.22 ± 0.04 | 0.24 ± 0.04 | 0.13 ± 0.03 | 0.45 ± 0.16 | 0.48 ± 0.13 | 0.28 ± 0.10 | 1.81 ± 0.47 |
| Soil T | 8.70 ± 0.03 | 34.58 ± 0.34 | 48.08 ± 14.02 | 38.65 ± 2.78 | 0.05 ± 0.00 | 0.22 ± 0.02 | 0.78 ± 0.19 | 0.41 ± 0.05 | 4.57 ± 1.32 | 5.16 ± 1.42 | 4.93 ± 0.28 | 15.98 ± 2.02 |
| Soil D | 8.32 ± 0.10 | 36.18 ± 0.94 | 117.48 ± 3.94 | 56.80 ± 1.76 | 1.32 ± 0.57 | 37.73 ± 4.80 | 148.50 ± 13.34 | 69.28 ± 10.45 | 222.75 ± 20.25 | 226.25 ± 17.75 | 134.78 ± 14.62 | 843.50 ± 66.78 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marchal, C.; Germain, J.; Raveton, M.; Lyonnard, B.; Arnoldi, C.; Binet, M.-N.; Mouhamadou, B. Molecular Characterization of Fungal Biodiversity in Long-Term Polychlorinated Biphenyl-Contaminated Soils. Microorganisms 2021, 9, 2051. https://doi.org/10.3390/microorganisms9102051
Marchal C, Germain J, Raveton M, Lyonnard B, Arnoldi C, Binet M-N, Mouhamadou B. Molecular Characterization of Fungal Biodiversity in Long-Term Polychlorinated Biphenyl-Contaminated Soils. Microorganisms. 2021; 9(10):2051. https://doi.org/10.3390/microorganisms9102051
Chicago/Turabian StyleMarchal, Camille, Joaquim Germain, Muriel Raveton, Blandine Lyonnard, Cindy Arnoldi, Marie-Noëlle Binet, and Bello Mouhamadou. 2021. "Molecular Characterization of Fungal Biodiversity in Long-Term Polychlorinated Biphenyl-Contaminated Soils" Microorganisms 9, no. 10: 2051. https://doi.org/10.3390/microorganisms9102051
APA StyleMarchal, C., Germain, J., Raveton, M., Lyonnard, B., Arnoldi, C., Binet, M. -N., & Mouhamadou, B. (2021). Molecular Characterization of Fungal Biodiversity in Long-Term Polychlorinated Biphenyl-Contaminated Soils. Microorganisms, 9(10), 2051. https://doi.org/10.3390/microorganisms9102051