
Independence of a Marine Unicellular Diazotroph to the Presence of NO3−

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
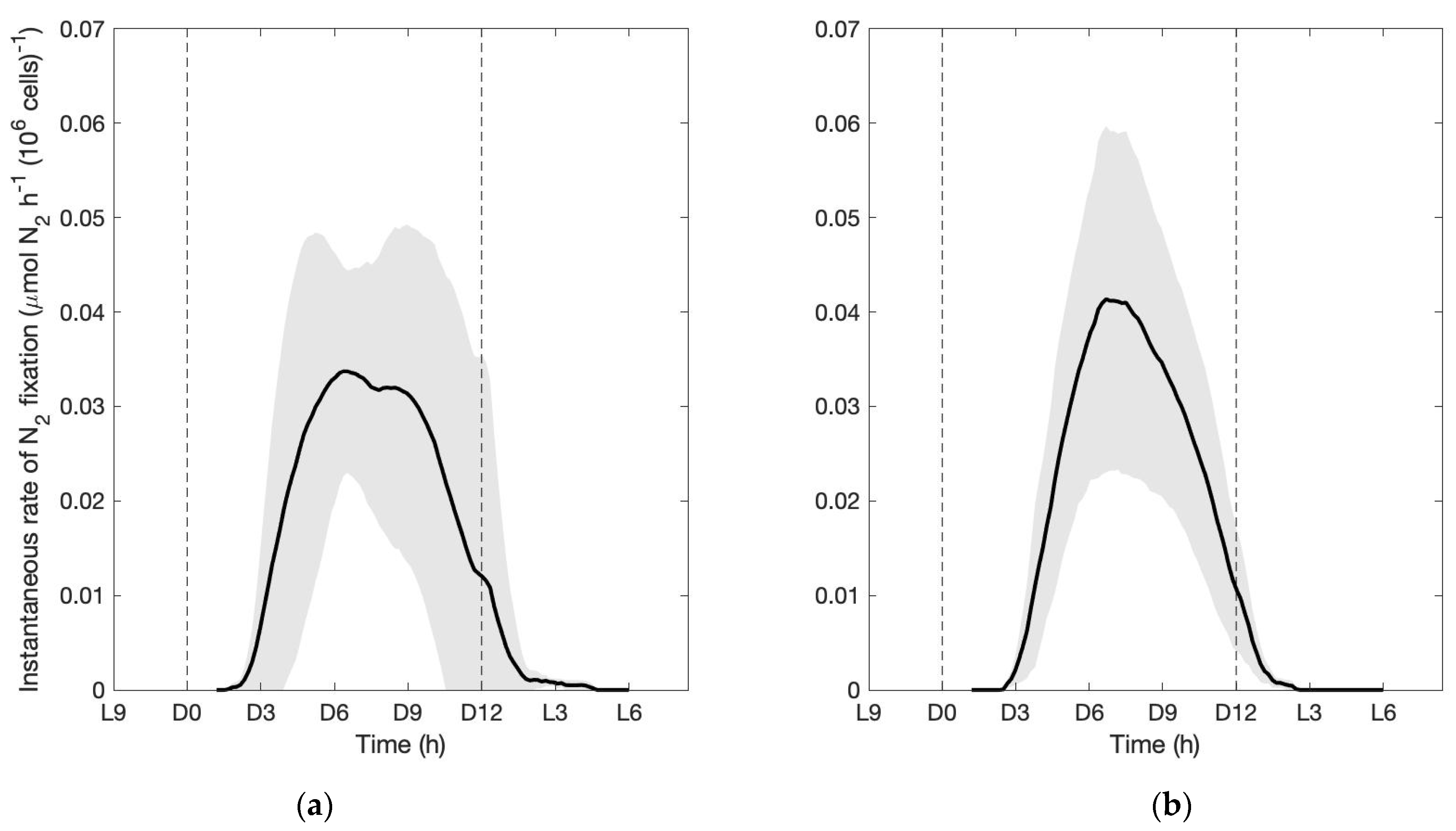
3. Results
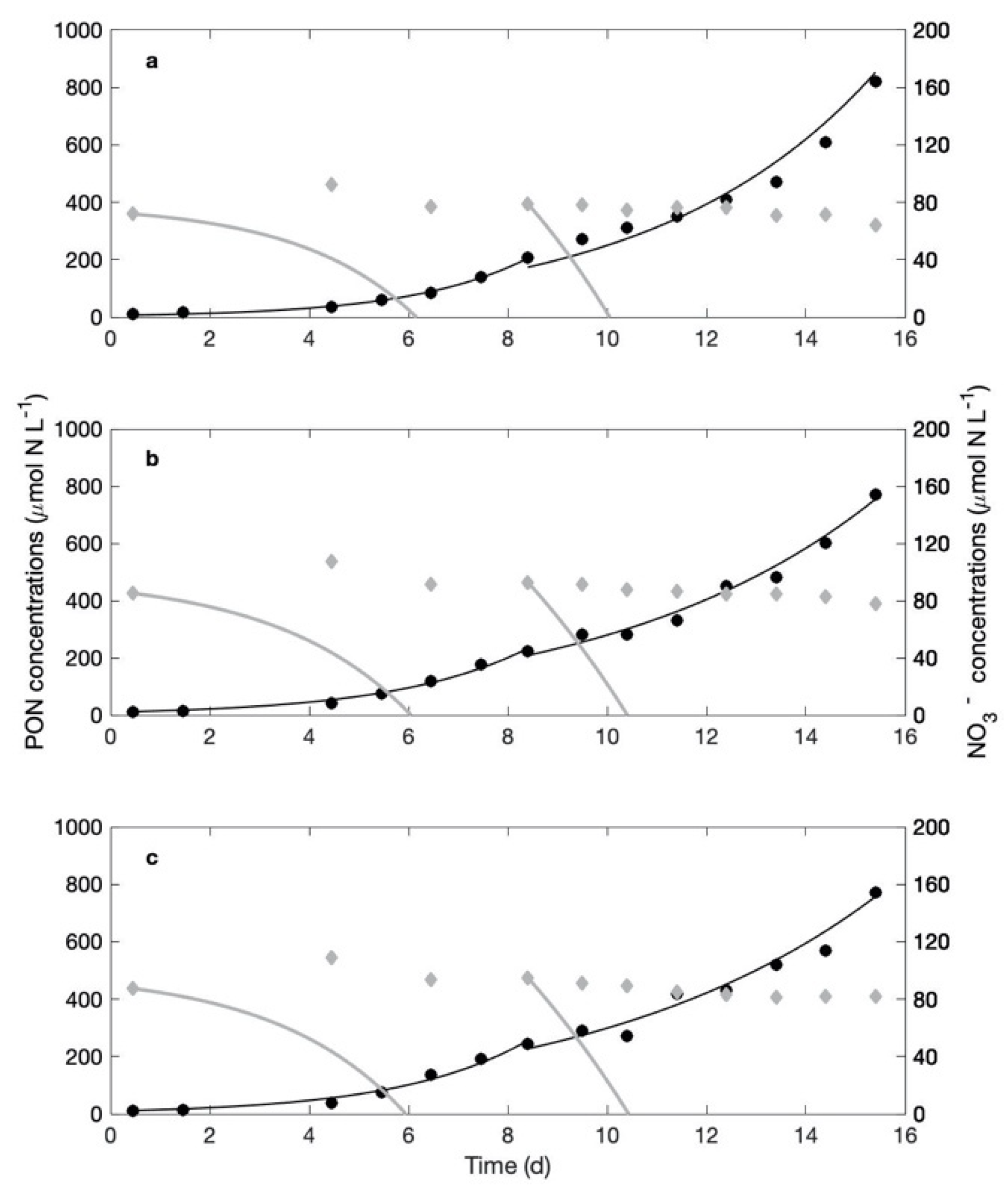
3.1. Control
3.2. Culture Experiments
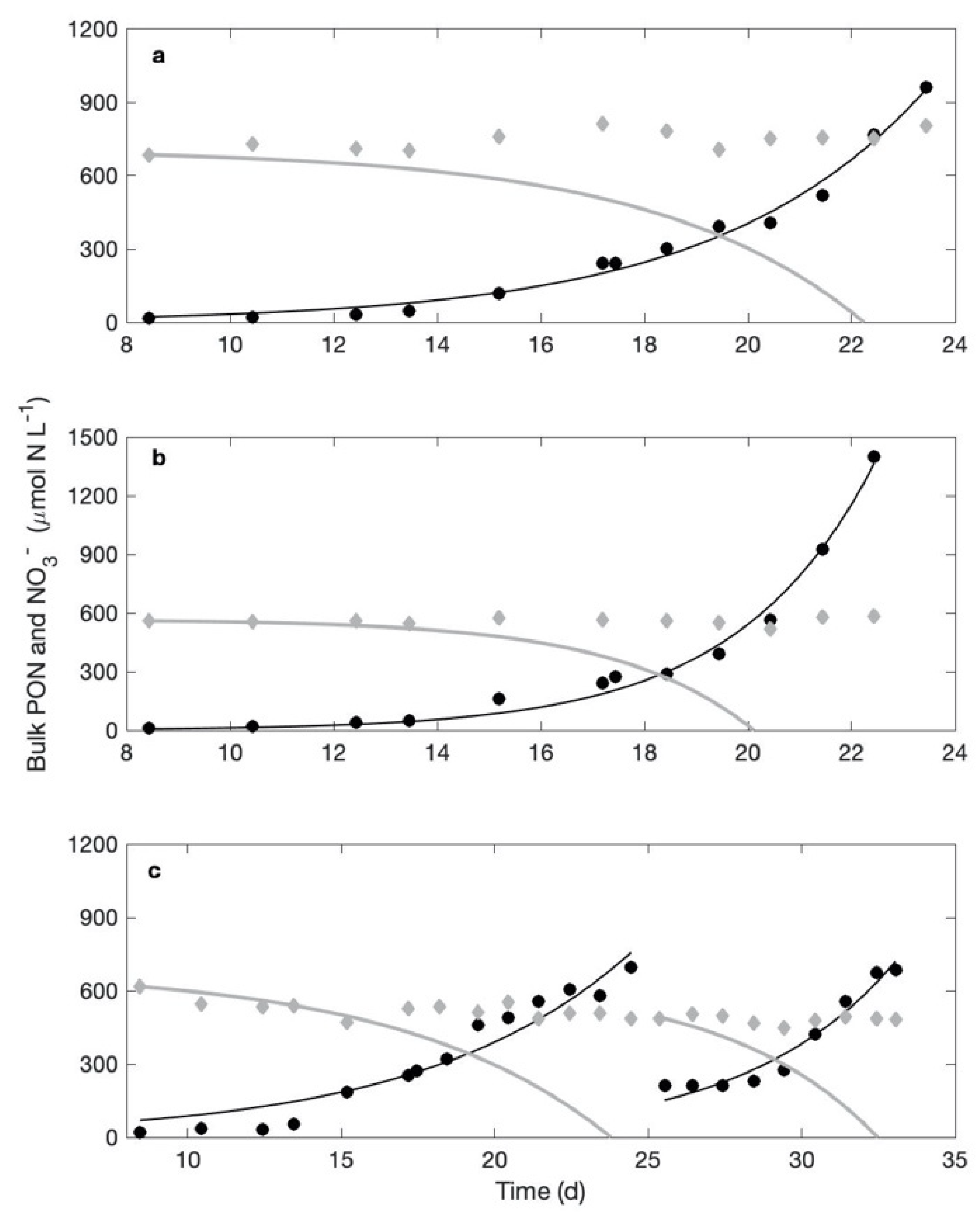
3.2.1. Experiment #1
3.2.2. Experiment #2
3.2.3. Experiment #3
Experiment #3-1
Experiment #3-2
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Falkowski, P.G. Enzymology of nitrogen assimilation. In Nitrogen in the Marine Environment; Carpenter, E.J., Capone, D.G., Eds.; Academic Press: San Diego, CA, USA, 1983; pp. 839–868. ISBN 978-0-12-160280-2. [Google Scholar]
- Großkopf, T.; LaRoche, J. Direct and Indirect Costs of Dinitrogen Fixation in Crocosphaera watsonii WH8501 and Possible Implications for the Nitrogen Cycle. Front. Microbiol. 2012, 3, 236. [Google Scholar] [CrossRef] [Green Version]
- Rabouille, S.; Campbell, D.A.; Masuda, T.; Zavřel, T.; Bernát, G.; Polerecky, L.; Halsey, K.; Eichner, M.; Kotabová, E.; Stephan, S.; et al. Electron and Biomass Dynamics of Cyanothece under Interacting Nitrogen and Carbon Limitations. Front. Microbiol. 2021, 12, 620. [Google Scholar] [CrossRef]
- Agawin, N.S.R.; Rabouille, S.; Veldhuis, M.J.W.; Servatius, L.; Hol, S.; van Overzee, H.M.J.; Huisman, J. Competition and Facilitation between Unicellular Nitrogen-Fixing Cyanobacteria and Non-Nitrogen-Fixing Phytoplankton Species. Limnol. Oceanogr. 2007, 52, 2233–2248. [Google Scholar] [CrossRef] [Green Version]
- Zehr, J.P.; Mellon, M.T.; Zani, S. New Nitrogen-Fixing Microorganisms Detected in Oligotrophic Oceans by Amplification of Nitrogenase (NifH) Genes. Appl. Environ. Microbiol. 1998, 64, 3444–3450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zehr, J.P.; Bench, S.R.; Carter, B.J.; Hewson, I.; Niazi, F.; Shi, T.; Tripp, H.J.; Affourtit, J.P. Globally Distributed Uncultivated Oceanic N2-Fixing Cyanobacteria Lack Oxygenic Photosystem II. Science 2008, 322, 1110–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripp, H.J.; Bench, S.R.; Turk, K.A.; Foster, R.A.; Desany, B.A.; Niazi, F.; Affourtit, J.P.; Zehr, J.P. Metabolic Streamlining in an Open-Ocean Nitrogen-Fixing Cyanobacterium. Nature 2010, 464, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Bombar, D.; Paerl, R.W.; Riemann, L. Marine Non-Cyanobacterial Diazotrophs: Moving beyond Molecular Detection. Trends Microbiol. 2016, 24, 916–927. [Google Scholar] [CrossRef] [PubMed]
- Moisander, P.H.; Beinart, R.A.; Hewson, I.; White, A.E.; Johnson, K.S.; Carlson, C.A.; Montoya, J.P.; Zehr, J.P. Unicellular Cyanobacterial Distributions Broaden the Oceanic N2 Fixation Domain. Science 2010, 327, 1512–1514. [Google Scholar] [CrossRef] [PubMed]
- Capone, D.G.; Zehr, J.P.; Paerl, H.W.; Bergman, B.; Carpenter, E.J. Trichodesmium, a Globally Significant Marine Cyanobacterium. Science 1997, 276, 1221–1229. [Google Scholar] [CrossRef] [Green Version]
- Knapp, A. The Sensitivity of Marine N2 Fixation to Dissolved Inorganic Nitrogen. Front. Microbiol. 2012, 3, 374. [Google Scholar] [CrossRef] [Green Version]
- Ohki, K.; Zehr, J.P.; Falkowski, P.G.; Fujita, Y. Regulation of Nitrogen Fixation by Different Nitrogen Sources in the Marine Non-Heterocystous Cyanobacterium Trichodesmium Sp. Nibb1067. Arch. Microbiol. 1991, 156, 335–337. [Google Scholar] [CrossRef]
- Mulholland, M.R.; Ohki, K.; Capone, D.G. Nutrient Controls on Nitrogen Uptake and Metabolism by Natural Populations and Cultures of Trichodesmium (Cyanobacteria). J. Phycol. 2001, 37, 1001–1009. [Google Scholar] [CrossRef]
- Fu, F.X.; Bell, P.R.F. Factors Affecting N2 Fixation by the Cyanobacterium Trichodesmium Sp. GBR-TRLI101. FEMS Microbiol. Ecol. 2003, 45, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Holl, C.M.; Montoya, J.P. Interactions between Nitrate Uptake and Nitrogen Fixation in Continuous Cultures of the Marine Diazotroph Trichodesmium (Cyanobacteria). J. Phycol. 2005, 41, 1178–1183. [Google Scholar] [CrossRef]
- Sandh, G.; Ran, L.; Xu, L.; Sundqvist, G.; Bulone, V.; Bergman, B. Comparative Proteomic Profiles of the Marine Cyanobacterium Trichodesmium Erythraeum IMS101 under Different Nitrogen Regimes. Proteomics 2011, 11, 406–419. [Google Scholar] [CrossRef]
- Knapp, A.N.; Dekaezemacker, J.; Bonnet, S.; Sohm, J.A.; Capone, D.G. Sensitivity of Trichodesmium Erythraeum and Crocosphaera watsonii Abundance and N2 Fixation Rates to Varying NO3− and PO43− Concentrations in Batch Cultures. Aquat. Microb. Ecol. 2012, 66, 223–236. [Google Scholar] [CrossRef]
- Aumont, O.; Ethe, C.; Tagliabue, A.; Bopp, L.; Gehlen, M. PISCES-v2: An Ocean Biogeochemical Model for Carbon and Ecosystem Studies. Geosci. Model Dev. 2015, 8, 2465–2513. [Google Scholar] [CrossRef] [Green Version]
- Dunne, J.P.; John, J.G.; Shevliakova, E.; Stouffer, R.J.; Krasting, J.P.; Malyshev, S.L.; Milly, P.C.D.; Sentman, L.T.; Adcroft, A.J.; Cooke, W.; et al. GFDL’s ESM2 Global Coupled Climate–Carbon Earth System Models. Part II: Carbon System Formulation and Baseline Simulation Characteristics. J. Clim. 2013, 26, 2247–2267. [Google Scholar] [CrossRef] [Green Version]
- Schmittner, A.; Oschlies, A.; Matthews, H.D.; Galbraith, E.D. Future Changes in Climate, Ocean Circulation, Ecosystems, and Biogeochemical Cycling Simulated for a Business-as-Usual CO2 Emission Scenario until Year 4000 AD. Glob. Biogeochem. Cycles 2008, 22. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.W.; Doney, S.C.; Anderson, L.A.; Benavides, M.; Berman-Frank, I.; Bode, A.; Bonnet, S.; Bostrom, K.H.; Bottjer, D.; Capone, D.G.; et al. Database of Diazotrophs in Global Ocean: Abundance, Biomass and Nitrogen Fixation Rates. Earth Syst. Sci. Data 2012, 4, 47–73. [Google Scholar] [CrossRef] [Green Version]
- Zehr, J.P.; Waterbury, J.B.; Turner, P.J.; Montoya, J.P.; Omoregie, E.; Steward, G.F.; Hansen, A.; Karl, D.M. Unicellular Cyanobacteria Fix N2 in the Subtropical North Pacific Ocean. Nature 2001, 412, 635–638. [Google Scholar] [CrossRef] [PubMed]
- Zehr, J.P. Nitrogen Fixation by Marine Cyanobacteria. Trends Microbiol. 2011, 19, 162–173. [Google Scholar] [CrossRef]
- Zehr, J.P.; Capone, D.G. Changing Perspectives in Marine Nitrogen Fixation. Science 2020, 368, eaay9514. [Google Scholar] [CrossRef]
- Dekaezemacker, J.; Bonnet, S. Sensitivity of N2 Fixation to Combined Nitrogen Forms (NO3− and NH4+) in Two Strains of the Marine Diazotroph Crocosphaera watsonii (Cyanobacteria). Mar. Ecol.-Prog. Ser. 2011, 438, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Garcia, N.S.; Hutchins, D.A. Light-Limited Growth Rate Modulates Nitrate Inhibition of Dinitrogen Fixation in the Marine Unicellular Cyanobacterium Crocosphaera watsonii. PLoS ONE 2014, 9, e114465. [Google Scholar] [CrossRef]
- Waterbury, J.B.; Watson, S.W.; Valois, F.W. Temporal Separation of Photosynthesis and Dinitrogen Fixation in the Marine Unicellular Cyanobacterium Erythrosphaera marina. EOS 1988, 69, 1089. [Google Scholar]
- Waterbury, J.B.; Rippka, R. The order Chroococcales. In Bergey’s Manual of Systematic Bacteriology; Krieg, N.R., Holt, J.B., Eds.; Williams & Wilkins: Baltimore, MD, USA, 1989; Volume 3, pp. 1728–1746. [Google Scholar]
- Rabouille, S.; Cabral, G.S.; Pedrotti, M.L. Towards a Carbon Budget of the Diazotrophic Cyanobacterium Crocosphaera: Effect of Irradiance. Mar. Ecol.-Prog. Ser. 2017, 570, 29–40. [Google Scholar] [CrossRef]
- Chen, Y.B.; Zehr, J.P.; Mellon, M. Growth and Nitrogen Fixation of the Diazotrophic Filamentous Non Heterocystous Cyanobacterium Trichodesmium Sp IMS 101 in Defined Media: Evidence for a Circadian Rhythm. J. Phycol. 1996, 32, 916–923. [Google Scholar] [CrossRef]
- Dron, A.; Rabouille, S.; Claquin, P.; Le Roy, B.; Talec, A.; Sciandra, A. Light-Dark (12:12) Cycle of Carbon and Nitrogen Metabolism in Crocosphaera watsonii WH8501: Relation to the Cell Cycle. Environ. Microbiol. 2012, 14, 967–981. [Google Scholar] [CrossRef]
- Malara, G.; Sciandra, A. A Multiparameter Phytoplankton Culture System Driven by Microcomputer. J. Appl. Phycol. 1991, 3, 235–241. [Google Scholar] [CrossRef]
- Montoya, J.P.; Voss, M.; Kahler, P.; Capone, D.G. A Simple, High-Precision, High-Sensitivity Tracer Assay for N2 Fixation. Appl. Environ. Microbiol. 1996, 62, 986–993. [Google Scholar] [CrossRef] [Green Version]
- Raimbault, P.; Garcia, N. Evidence for Efficient Regenerated Production and Dinitrogen Fixation in Nitrogen-Deficient Waters of the South Pacific Ocean: Impact on New and Export Production Estimates. Biogeosciences 2008, 5, 323–338. [Google Scholar] [CrossRef] [Green Version]
- Dron, A.; Rabouille, S.; Claquin, P.; Talec, A.; Raimbault, V.; Sciandra, A. Photoperiod Length Paces the Temporal Orchestration of Cell Cycle and Carbon-Nitrogen Metabolism in Crocosphaera watsonii. Environ. Microbiol. 2013, 15, 3292–3304. [Google Scholar] [CrossRef] [PubMed]
- Glibert, P.M.; Bronk, D.A. Release of Dissolved Organic Nitrogen by Marine Diazotrophic Cyanobacteria, Trichodesmium spp. Appl. Environ. Microbiol. 1994, 60, 3996–4000. [Google Scholar] [CrossRef] [Green Version]
- Mulholland, M.R.; Bernhardt, P.W. The Effect of Growth Rate, Phosphorus Concentration, and Temperature on N2 Fixation, Carbon Fixation, and Nitrogen Release in Continuous Cultures of Trichodesmium IMS101. Limnol. Oceanogr. 2005, 50, 839–849. [Google Scholar] [CrossRef]
- Großkopf, T.; Mohr, W.; Baustian, T.; Schunck, H.; Gill, D.; Kuypers, M.M.M.; Lavik, G.; Schmitz, R.A.; Wallace, D.W.R.; LaRoche, J. Doubling of Marine Dinitrogen-Fixation Rates Based on Direct Measurements. Nature 2012, 488, 361–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, T.; Ilikchyan, I.; Rabouille, S.; Zehr, J.P. Genome-Wide Analysis of Diel Gene Expression in the Unicellular N-2-Fixing Cyanobacterium Crocosphaera watsonii WH 8501. ISME J. 2010, 4, 621–632. [Google Scholar] [CrossRef] [Green Version]
- Gallon, J.R. Reconciling the Incompatible—N2 Fixation and O2. New Phytol. 1992, 122, 571–609. [Google Scholar] [CrossRef]
- Hood, R.R.; Bates, N.R.; Capone, D.G.; Olson, D.B. Modeling the Effect of Nitrogen Fixation on Carbon and Nitrogen Fluxes at BATS. Deep-Sea Res. Part II 2001, 48, 1609–1648. [Google Scholar] [CrossRef]
- Fennel, K.; Spitz, Y.H.; Letelier, R.M.; Abbott, M.R.; Karl, D.M. A Deterministic Model for N2 Fixation at Stn. ALOHA in the Subtropical North Pacific Ocean. Deep-Sea Res. Part II 2002, 49, 149–174. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.K.; Doney, S.C.; Kleypas, J.A.; Glover, D.M.; Fung, I.Y. An Intermediate Complexity Marine Ecosystem Model for the Global Domain. Deep-Sea Res. Part II-Top. Stud. Oceanogr. 2002, 49, 403–462. [Google Scholar] [CrossRef] [Green Version]
- Deutsch, C.; Sarmiento, J.L.; Sigman, D.M.; Gruber, N.; Dunne, J.P. Spatial Coupling of Nitrogen Inputs and Losses in the Ocean. Nature 2007, 445, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Maier-Reimer, E.; Kriest, I.; Segschneider, J.; Wetzel, P. The HAMburg Ocean Carbon Cycle Model HAMOCC5.1—Technical Description Release 1.1. Berichte zur Erdsystemforschung 2004, 14. [Google Scholar]
- Ilyina, T.; Six, K.D.; Segschneider, J.; Maier-Reimer, E.; Li, H.; Núñez-Riboni, I. Global Ocean Biogeochemistry Model HAMOCC: Model Architecture and Performance as Component of the MPI-Earth System Model in Different CMIP5 Experimental Realizations. J. Adv. Model. Earth Syst. 2013, 5, 287–315. [Google Scholar] [CrossRef]
- Kriest, I.; Oschlies, A. MOPS-1.0: Towards a Model for the Regulation of the Global Oceanic Nitrogen Budget by Marine Biogeochemical Processes. Geosci. Model Dev. 2015, 8, 2929–2957. [Google Scholar] [CrossRef] [Green Version]
- Redfield, A.C. On the Proportions of Organic Derivations in Sea Water and Their Relation to the Composition of Plankton. In James Johnstone Memorial Volume; Daniel, R.J., Ed.; University Press of Liverpool: Merseyside, UK, 1934; pp. 177–192. [Google Scholar]
- Redfield, A.C.; Ketchum, B.H.; Richards, F.A. The Influence of Organisms on the Composition of Sea-Water. In The Composition of Seawater. Comparative and Descriptive Oceanography. The Sea: Ideas and Observations on Progress in the Study of the Seas; Hill, M.N., Ed.; Wiley-Interscience: New York, NY, USA, 1963; Volume 2, pp. 26–77. [Google Scholar]
- Moisander, P.H.; Serros, T.; Paerl, R.W.; Beinart, R.A.; Zehr, J.P. Gammaproteobacterial Diazotrophs and NifH Gene Expression in Surface Waters of the South Pacific Ocean. ISME J. 2014, 8, 1962–1973. [Google Scholar] [CrossRef] [PubMed]
- Blais, M.; Tremblay, J.-É.; Jungblut, A.D.; Gagnon, J.; Martin, J.; Thaler, M.; Lovejoy, C. Nitrogen Fixation and Identification of Potential Diazotrophs in the Canadian Arctic. Glob. Biogeochem. Cycles 2012, 26. [Google Scholar] [CrossRef]
- Shiozaki, T.; Fujiwara, A.; Ijichi, M.; Harada, N.; Nishino, S.; Nishi, S.; Nagata, T.; Hamasaki, K. Diazotroph Community Structure and the Role of Nitrogen Fixation in the Nitrogen Cycle in the Chukchi Sea (Western Arctic Ocean). Limnol. Oceanogr. 2018, 63, 2191–2205. [Google Scholar] [CrossRef] [Green Version]
- Shiozaki, T.; Fujiwara, A.; Inomura, K.; Hirose, Y.; Hashihama, F.; Harada, N. Biological Nitrogen Fixation Detected under Antarctic Sea Ice. Nat. Geosci. 2020, 13, 729–732. [Google Scholar] [CrossRef]
- Hewson, I.; Moisander, P.H.; Achilles, K.M.; Carlson, C.A.; Jenkins, B.D.; Mondragon, E.A.; Morrison, A.E.; Zehr, J.P. Characteristics of Diazotrophs in Surface to Abyssopelagic Waters of the Sargasso Sea. Aquat. Microb. Ecol. 2007, 46, 15–30. [Google Scholar] [CrossRef]
- Moisander, P.H.; Benavides, M.; Bonnet, S.; Berman-Frank, I.; White, A.E.; Riemann, L. Chasing after Non-Cyanobacterial Nitrogen Fixation in Marine Pelagic Environments. Front. Microbiol. 2017, 8, 1736. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Experiment #1 | Experiment #2 | Experiment #3 | ||||
|---|---|---|---|---|---|---|---|
| 3.1 | 3.2 | ||||||
| Light regime | LD | LL | LD | LD | |||
| irradiance (µE m−2 s−1) | 240 | 120 | 220 | 260 | |||
| NO3− (μmol N·L−1) | (+) | (−) | (+) | (+) | (−) | (+) | (−) |
| Growth rate (d−1) | 0.30, 0.31 | 0.43, 0.35 ∅ | 0.19, 0.22 | 0.32 ± 0.02 | 0.34 ± 0.02 ∅ | 0.39 ± 0.03 | 0.37 ± 0.06 ∅ |
| N content (fmolN cell−1) | 33.22 | 48.52 | 62.47 ± 9.74 | 55.4 ± 1.44 | |||
| Max NO3− support (%) | 10.4, 2.3 | NA | 24.55, 27.54 | 0, 10.5, 1.6 | 1.90 ± 0.45 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rabouille, S.; Randall, B.; Talec, A.; Raimbault, P.; Blasco, T.; Latifi, A.; Oschlies, A. Independence of a Marine Unicellular Diazotroph to the Presence of NO3−. Microorganisms 2021, 9, 2073. https://doi.org/10.3390/microorganisms9102073
Rabouille S, Randall B, Talec A, Raimbault P, Blasco T, Latifi A, Oschlies A. Independence of a Marine Unicellular Diazotroph to the Presence of NO3−. Microorganisms. 2021; 9(10):2073. https://doi.org/10.3390/microorganisms9102073
Chicago/Turabian StyleRabouille, Sophie, Benjamin Randall, Amélie Talec, Patrick Raimbault, Thierry Blasco, Amel Latifi, and Andreas Oschlies. 2021. "Independence of a Marine Unicellular Diazotroph to the Presence of NO3−" Microorganisms 9, no. 10: 2073. https://doi.org/10.3390/microorganisms9102073
APA StyleRabouille, S., Randall, B., Talec, A., Raimbault, P., Blasco, T., Latifi, A., & Oschlies, A. (2021). Independence of a Marine Unicellular Diazotroph to the Presence of NO3−. Microorganisms, 9(10), 2073. https://doi.org/10.3390/microorganisms9102073