
Intraspecies Variation in Tetrahymena rostrata

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture
2.2. Morphological and Phenotypic Characterisation
2.3. Genome Sequencing
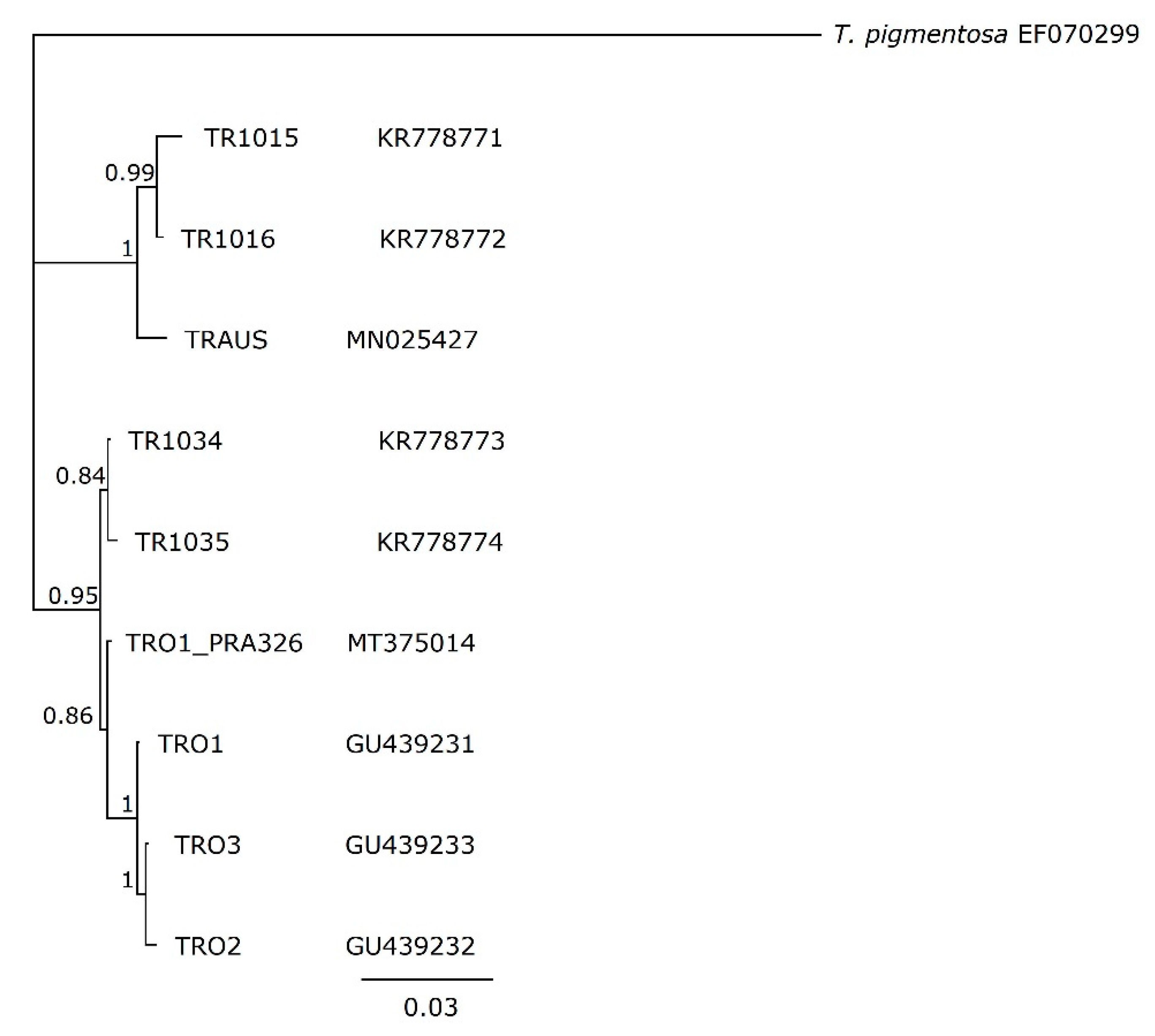
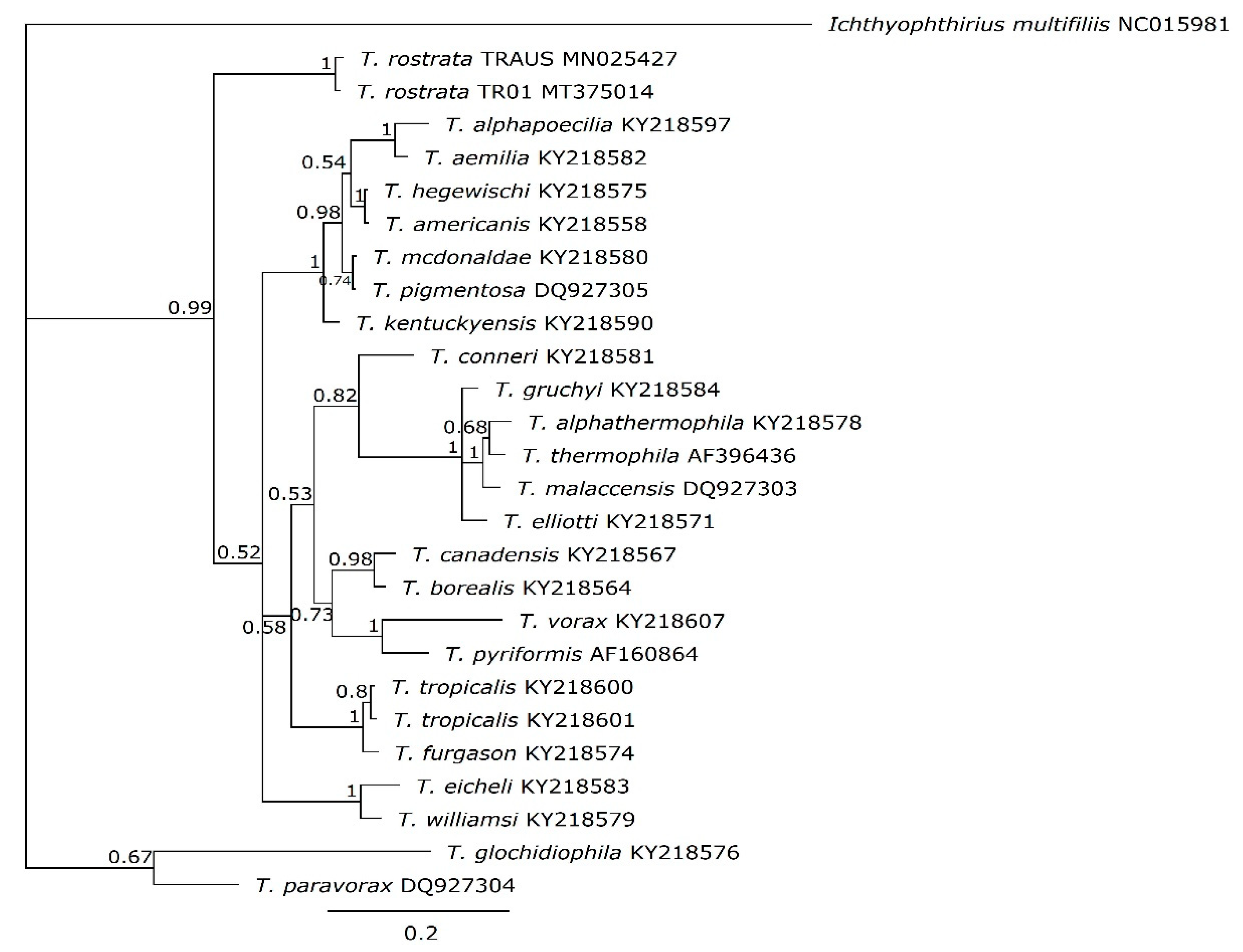
2.4. Phylogenetic Analysis
3. Results and Discussion
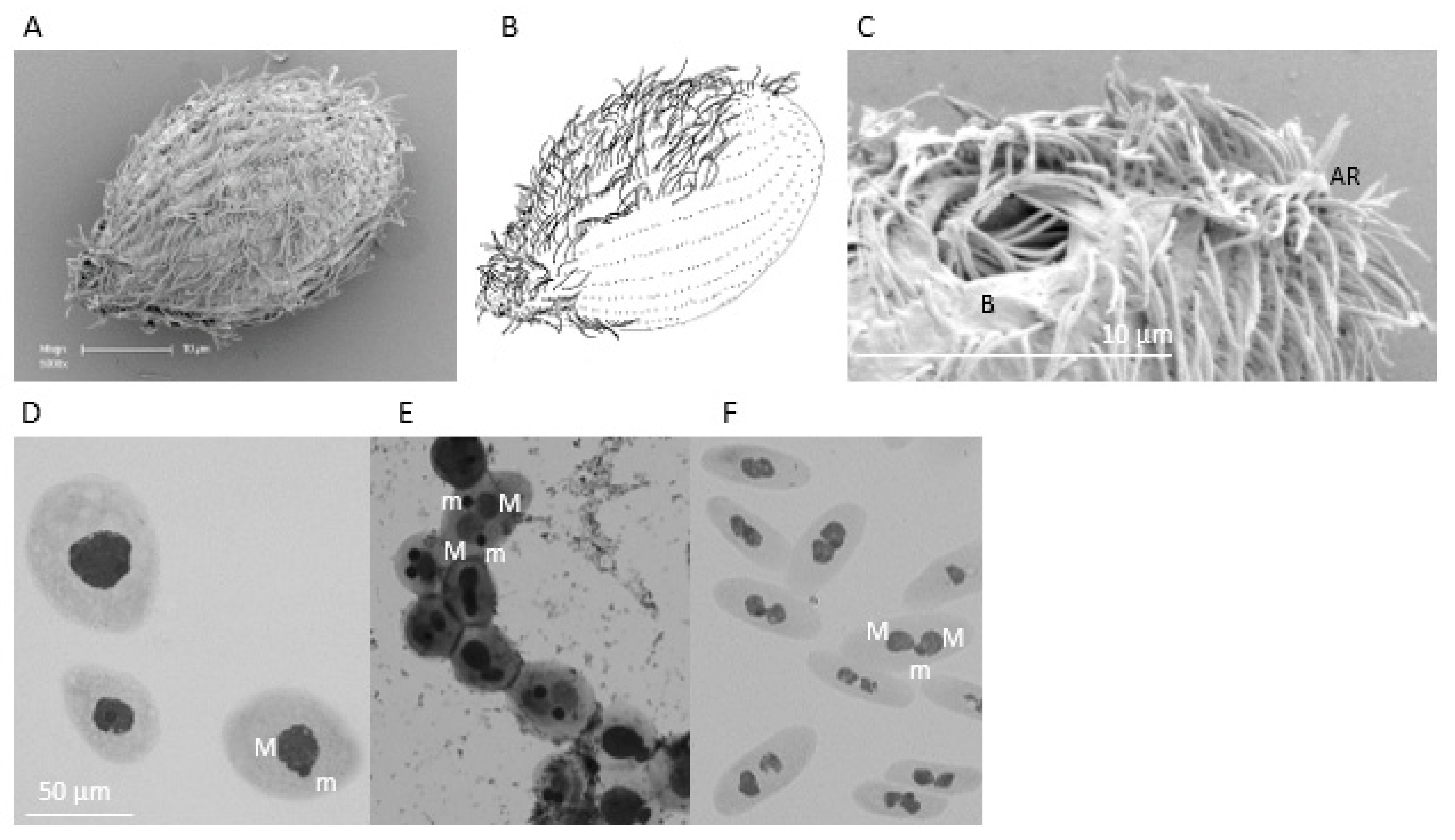
3.1. Morphological and Phenotypic Examination of Isolate TRAUS
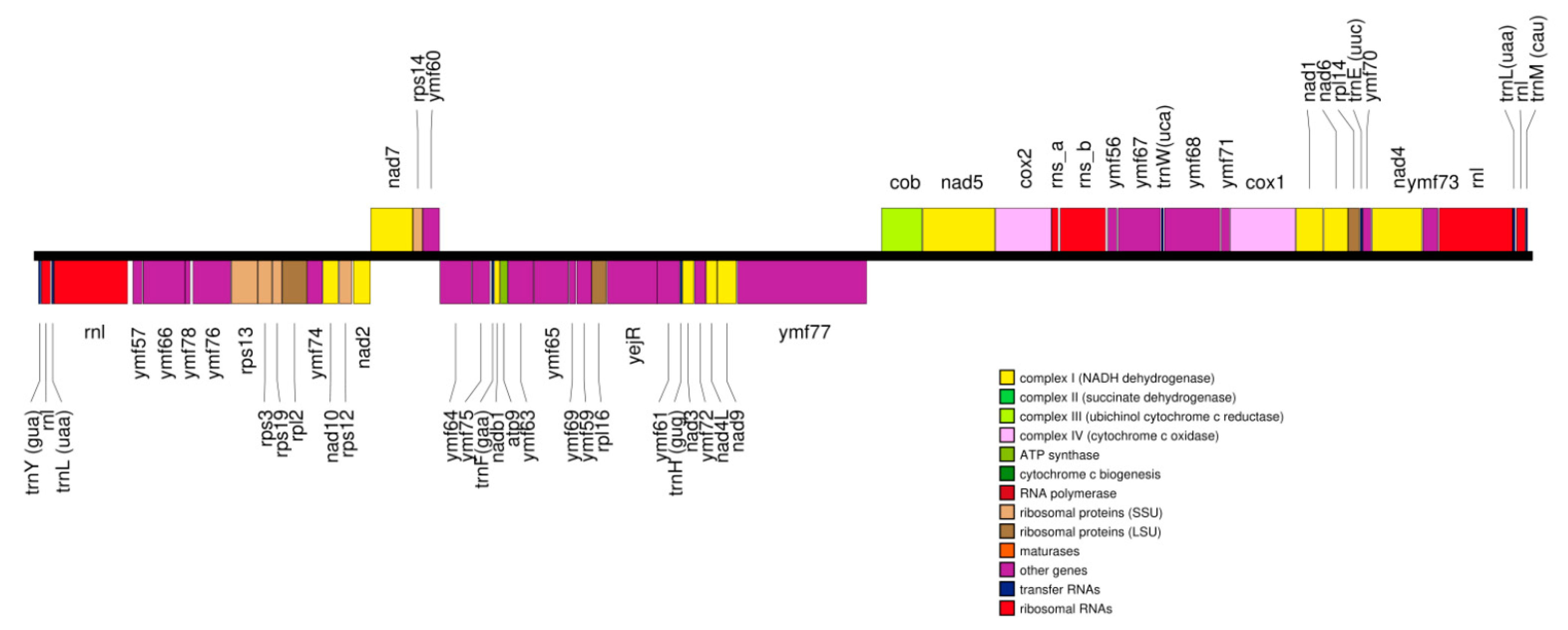
3.2. Mitogenomes of TR01 and TRAUS
3.3. Cytochrome Oxidase 1 Gene
3.4. HCEs and Central Repeat Region
3.5. mtSSUrRNA
3.6. Nuclear Genes
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Corliss, J.O. Le cycle autogamique de Tetrahymena rostrata. Comptes Rendus Hebd. Seances L’acad. Sci. 1952, 235, 399–402. [Google Scholar]
- Kozloff, E.N. A species of Tetrahymena parasitic in the renal organ of the slug Deroceras reticulatum. J. Protozool. 1957, 4, 75–79. [Google Scholar] [CrossRef]
- McArdle, E.W.; Bergquist, B.L.; Ehret, C.F. Strucural changes in Tetrahymena rostrata during induced encystment. J. Protozool. 1980, 27, 388–397. [Google Scholar] [CrossRef]
- Stout, J. The ecology, life history and parasitism of Tetrahymena [Paraglaucoma] rostrata (Kahl) Corliss. J. Protozool. 1954, 1, 211–215. [Google Scholar] [CrossRef]
- Smith, D.G.S.; Gawryluk, R.M.R.; Spencer, D.F.; Pearlman, R.E.; Siu, K.W.M.; Gray, M.W. Exploring the mitochondrial proteome of the ciliate protozoon Tetrahymena thermophila: Direct analysis by tandem mass spectrometry. J. Mol. Biol. 2007, 374, 837–863. [Google Scholar] [CrossRef]
- Kaczanowski, A.; Brunk, C.F.; Kazubski, S.L. Cohesion of clonal life history, senescence and rejuvenation induced by autogamy of the histophagous ciliate Tetrahymena rostrata. Protist 2016, 167, 490–510. [Google Scholar] [CrossRef]
- Segade, P.; Garcia-Estevez, J.; Arias, C.; Iglesias, R. Parasitic infections in mixed system-based heliciculture farms: Dynamics and key epidemiological factors. Parasitology 2013, 140, 482–497. [Google Scholar] [CrossRef]
- Van As, J.; Basson, I. Ciliophoran (Ciliophora) Parasites of Terrestrial Gastropods; CABI Publishing: Wallingford, UK, 2004; pp. 559–578. [Google Scholar]
- Brooks, W. Tetrahymenid ciliates as parasites of the gray garden slug. Hilgardia 1968, 39, 205–276. [Google Scholar] [CrossRef] [Green Version]
- Segade, P.; Kher, C.P.; Lynn, D.H.; Iglesias, R. Morphological and molecular characterization of renal ciliates infecting farmed snails in Spain. Parasitology 2009, 136, 771–782. [Google Scholar] [CrossRef]
- Kher, C.P.; Doerder, F.P.; Cooper, J.; Ikonomi, P.; Achilles-Day, U.; Küpper, F.C.; Lynn, D.H. Barcoding Tetrahymena: Discriminating species and identifying unknowns using the cytochrome c oxidase subunit 1 (cox-1) barcode. Protist 2011, 162, 2–13. [Google Scholar] [CrossRef]
- Doerder, F.P. Barcodes reveal 48 new species of Tetrahymena, Dexiostoma, and Glaucoma: Phylogeny, ecology, and biogeography of new and established species. J. Eukaryot. Microbiol. 2019, 66, 182–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- South, A. Terrestrial Slugs: Biology, Ecology and Control; Chapman and Hall: London, UK, 1992. [Google Scholar]
- Watt, A.; Haites, R.; Young, N.; Billman-Jacobe, H. The mitochondrial genome of Tetrahymena rostrata. Mitochondrial DNA Part B-Resour. 2020, 5, 53–54. [Google Scholar] [CrossRef]
- Gao, F.; Li, J.; Song, W.; Xu, D.; Warren, A.; Yi, Z.; Gao, S. Multi-gene-based phylogenetic analysis of oligotrich ciliates with emphasis on two dominant groups: Cyrtostrombidiids and Strombidiids (Protozoa, Ciliophora). Mol. Phylogenet. Evol. 2016, 105, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. Nih image to imagej: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Brunk, C.F.; Lee, L.C.; Tran, A.B.; Li, J. Complete sequence of the mitochondrial genome of Tetrahymena thermophila and comparative methods for identifying highly divergent genes. Nucleic Acids Res. 2003, 31, 1673–1682. [Google Scholar] [CrossRef] [Green Version]
- Burger, G.; Zhu, Y.; Littlejohn, T.G.; Greenwood, S.J.; Schnare, M.N.; Lang, B.F.; Gray, M.W. Complete sequence of the mitochondrial genome of Tetrahymena pyriformis and comparison with Paramecium aurelia mitochondrial DNA. J. Mol. Biol. 2000, 297, 365–380. [Google Scholar] [CrossRef] [Green Version]
- Moradian, M.M.; Beglaryan, D.; Skozylas, J.M.; Kerikorian, V. Complete mitochondrial genome sequence of three Tetrahymena species reveals mutation hot spots and accelerated non-synonymous substitutions in ymf genes. PLoS ONE 2007, 2, e650. [Google Scholar] [CrossRef]
- Chantangsi, C.; Struder-Kypke, M.C.; Lynn, D.H. Barcoding protists: A trailblazing study on a ciliate genus of closely related species, Tetrahymena. J. Eukaryot. Microbiol. 2007, 54, 10S. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. Mega x: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Gershgorin, R.A.; Gorbunov, K.Y.; Zverkov, O.A.; Rubanov, L.I.; Seliverstov, A.V.; Lyubetsky, V.A. Highly conserved elements and chromosome structure evolution in mitochondrial genomes in ciliates. Life 2017, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Li, C.; Zhang, X.; Wang, C.; Roger, A.J.; Gao, F. Characterization and comparative analyses of mitochondrial genomes in single-celled eukaryotes to shed light on the diversity and evolution of linear molecular architecture. Int. J. Mol. Sci. 2021, 22, 2546. [Google Scholar] [CrossRef] [PubMed]
- Segade, P.; Garcia, N.; Garcia Estevez, J.M.; Arias, C.; Iglesias, R. Encystment/excystment response and serotypic variation in the gastropod parasite Tetrahymena rostrata (Ciliophora, Tetrahymenidae). Parasitol. Res. 2016, 115, 771–777. [Google Scholar] [CrossRef]
- Nielsen, H.; Engberg, J. Sequence comparison of the rdna introns from six different species of Tetrahymena. Nucleic Acids Res. 1985, 13, 7445–7455. [Google Scholar] [CrossRef] [Green Version]
- Katz, L.A.; Snoeyenbos-West, O.; Doerder, F.P. Patterns of protein evolution in Tetrahymena thermophila: Implications for estimates of effective population size. Mol. Biol. Evol. 2006, 23, 608–614. [Google Scholar] [CrossRef] [Green Version]
- Yi, Z.; Huang, L.; Yang, R.; Lin, X.; Song, W. Actin evolution in ciliates (Protist, alveolata) is characterized by high diversity and three duplication events. Mol. Phylogenet. Evol. 2016, 96, 45–54. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain ID | Host | Tissue | Accession Numbers for Each Isolate | Ref. |
|---|---|---|---|---|
| TR01 | Helix aspersa aspersa | renal organ | GU439231 (cox1), JQ045342 (18S rRNA) | [9] |
| MT375014 (mtDNA), MT506240 (histone H3 H4), MT420428 (18S-5.8S-28S rRNA), SRR12315381(short read archive) | This study | |||
| TR02 | Helix aspersa maxima | renal organ | GU439232 (cox1) | [9] |
| TR03 | Deroceras reticulatum | renal organ | GU439233 (cox1) | [9] |
| TR1015 | Zonitoides nitidus | renal organ | KR778771 (cox1); KR778775 (18S rRNA) | [5] |
| TR1016 | Zonitoides nitidus | renal organ | KR778772 (cox1), KR778776 (18S rRNA) | [5] |
| TR1034 | Cochlicopa lubrica | renal organ | KR778773 (cox1), KR778777 (18S rRNA) | [5] |
| TR1035 | Cochlicopa lubrica | renal organ | KR778774 (cox1), KR778778 (18S rRNA) | [5] |
| TRAUS | Deroceras reticulatum | egg | MN025427 (mtDNA), MN167836 (histone H3 | |
| H4), MN158348 (18S, 5.8S, 28S rRNA), SRR12315411 (short read archive) | [12] and this study |
| CDS | bp | DNA Seq Identity (%) | Amino Acid Seq Similarity (%) |
|---|---|---|---|
| Atp9 | 228 | 99.12 | 100 |
| Rps14 | 306 | 99.02 | 100 |
| Ymf57 | 303 | 98.68 | 100 |
| Ymf78 | 168 | 98.21 | 100 |
| Ymf56 | 294 | 97.96 | 100 |
| Ymf72 | 357 | 97.76 | 99.16 |
| Nad4L | 351 | 97.44 | 100 |
| RpS12 | 402 | 97.26 | 100 |
| Ymf69 | 216 | 97.22 | 100 |
| Ymf61 | 720 | 97.08 | 99.58 |
| Ymf65 | 2652 | 97.05 | 100 |
| Nad2 | 537 | 97.02 | 100 |
| Ymf64 | 999 | 97 | 99.7 |
| Nad10 | 489 | 96.93 | 100 |
| Ymf76 | 1194 | 96.82 | 99.75 |
| RpS3 | 456 | 96.77 | 100 |
| RpL16 | 465 | 96.77 | 100 |
| Ymf74 | 492 | 96.75 | 100 |
| Cob | 1287 | 96.74 | 99.77 |
| Nad1b | 180 | 96.67 | 100 |
| Ymf70 | 270 | 96.67 | 98.89 |
| Nad7 | 1329 | 96.61 | 99.77 |
| RpS13 | 834 | 96.52 | 99.28 |
| Nad3 | 366 | 96.45 | 100 |
| Ymf75 | 567 | 96.3 | 98.41 |
| Ymf63 | 816 | 96.2 | 100 |
| Cox1 | 2067 | 96.18 | 99.71 |
| RpL2 | 789 | 96.07 | 100 |
| RpS19 | 297 | 95.96 | 98.99 |
| Ymf66 | 1329 | 95.71 | 100 |
| Nad1a | 855 | 95.67 | 100 |
| Ymf68 | 1761 | 95.57 | 99.32 |
| RpL14 | 360 | 95.56 | 100 |
| Nad4 | 1578 | 95.56 | 97.72 |
| YejR | 1581 | 95.51 | 98.67 |
| Nad6 | 762 | 95.41 | 98.92 |
| Ymf60 | 534 | 95.13 | 99.44 |
| Ymf67 | 1344 | 95.09 | 98.44 |
| Ymf73 | 483 | 95.03 | 100 |
| Nad9 | 597 | 94.97 | 98.99 |
| Nad5 | 2280 | 94.91 | 99.21 |
| Cox2 | 588 | 94.9 | 98.64 |
| Ymf59 | 471 | 94.69 | 98.73 |
| Ymf71 | 264 | 94.32 | 97.73 |
| Ymf77 | 4077 | 92.62 | 96.98 |

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watt, A.; Young, N.; Haites, R.; Dunse, K.; Russell, D.; Billman-Jacobe, H. Intraspecies Variation in Tetrahymena rostrata. Microorganisms 2021, 9, 2100. https://doi.org/10.3390/microorganisms9102100
Watt A, Young N, Haites R, Dunse K, Russell D, Billman-Jacobe H. Intraspecies Variation in Tetrahymena rostrata. Microorganisms. 2021; 9(10):2100. https://doi.org/10.3390/microorganisms9102100
Chicago/Turabian StyleWatt, Anne, Neil Young, Ruth Haites, Kerry Dunse, Derek Russell, and Helen Billman-Jacobe. 2021. "Intraspecies Variation in Tetrahymena rostrata" Microorganisms 9, no. 10: 2100. https://doi.org/10.3390/microorganisms9102100
APA StyleWatt, A., Young, N., Haites, R., Dunse, K., Russell, D., & Billman-Jacobe, H. (2021). Intraspecies Variation in Tetrahymena rostrata. Microorganisms, 9(10), 2100. https://doi.org/10.3390/microorganisms9102100