
Development of Plant–Fungal Endophyte Associations to Suppress Phoma Stem Canker in Brassica

 , , ,
, , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Endophytes Isolated from Wild Brassica
2.2. Development of Novel Brassica–Endophyte Associations
2.3. Inoculation of Brassica Seed with Fungal Spore Suspensions
2.4. Assessment of Endophyte Colonisation
2.5. Bioactivity of Fungal Endophytes towards Leptosphaeria Maculans
3. Results
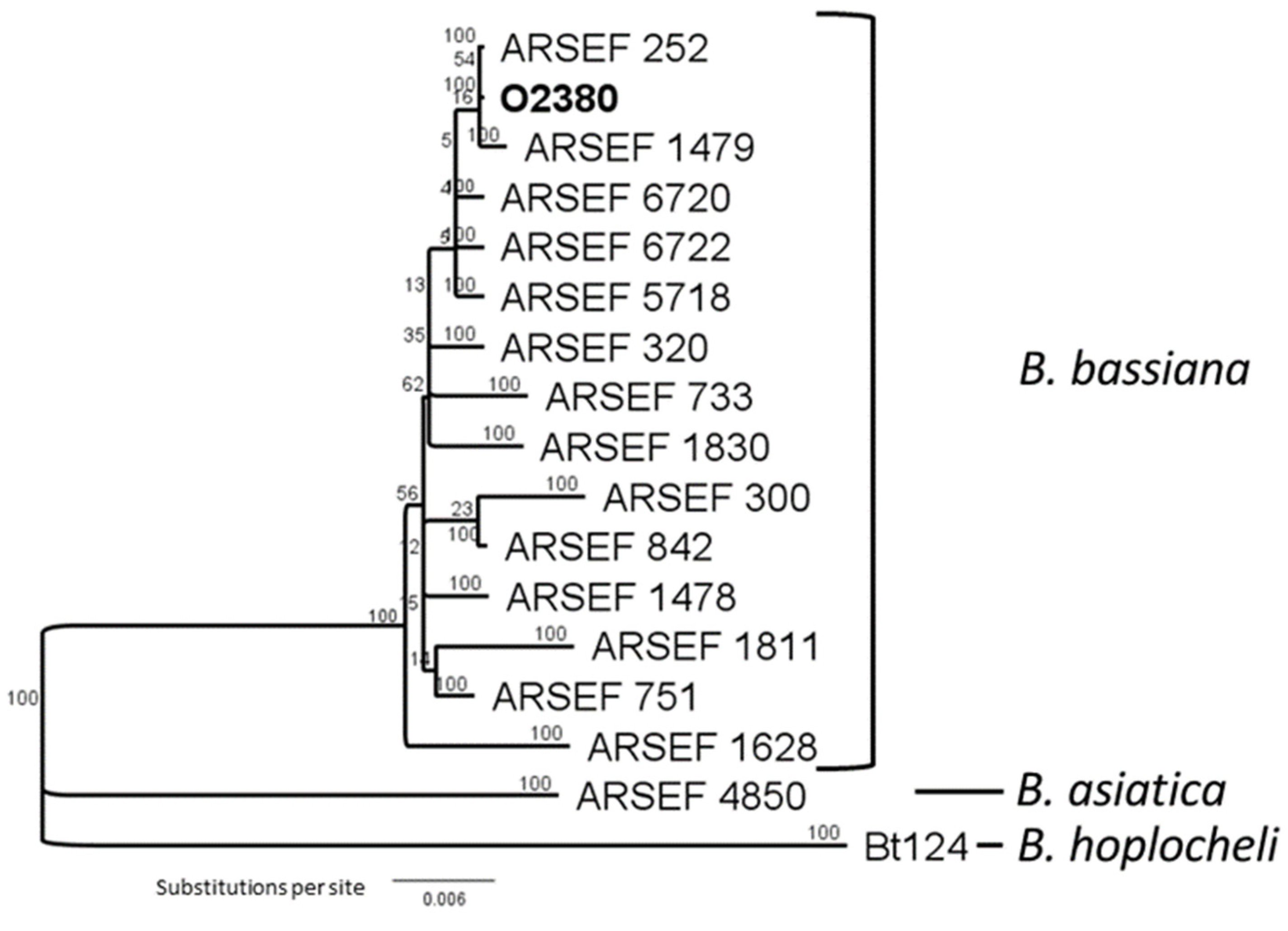
3.1. Fungal Endophytes Isolated from Wild Brassica
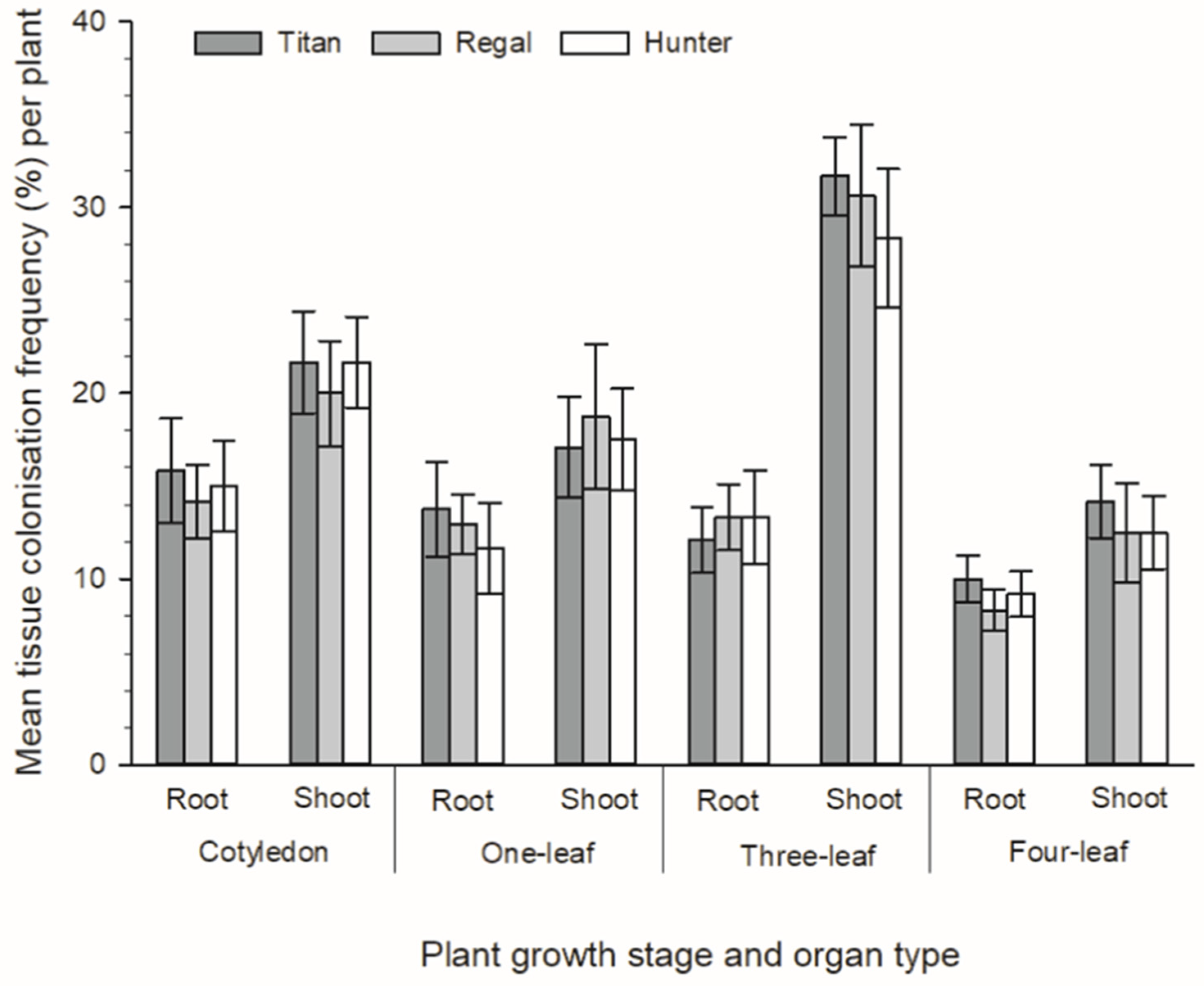
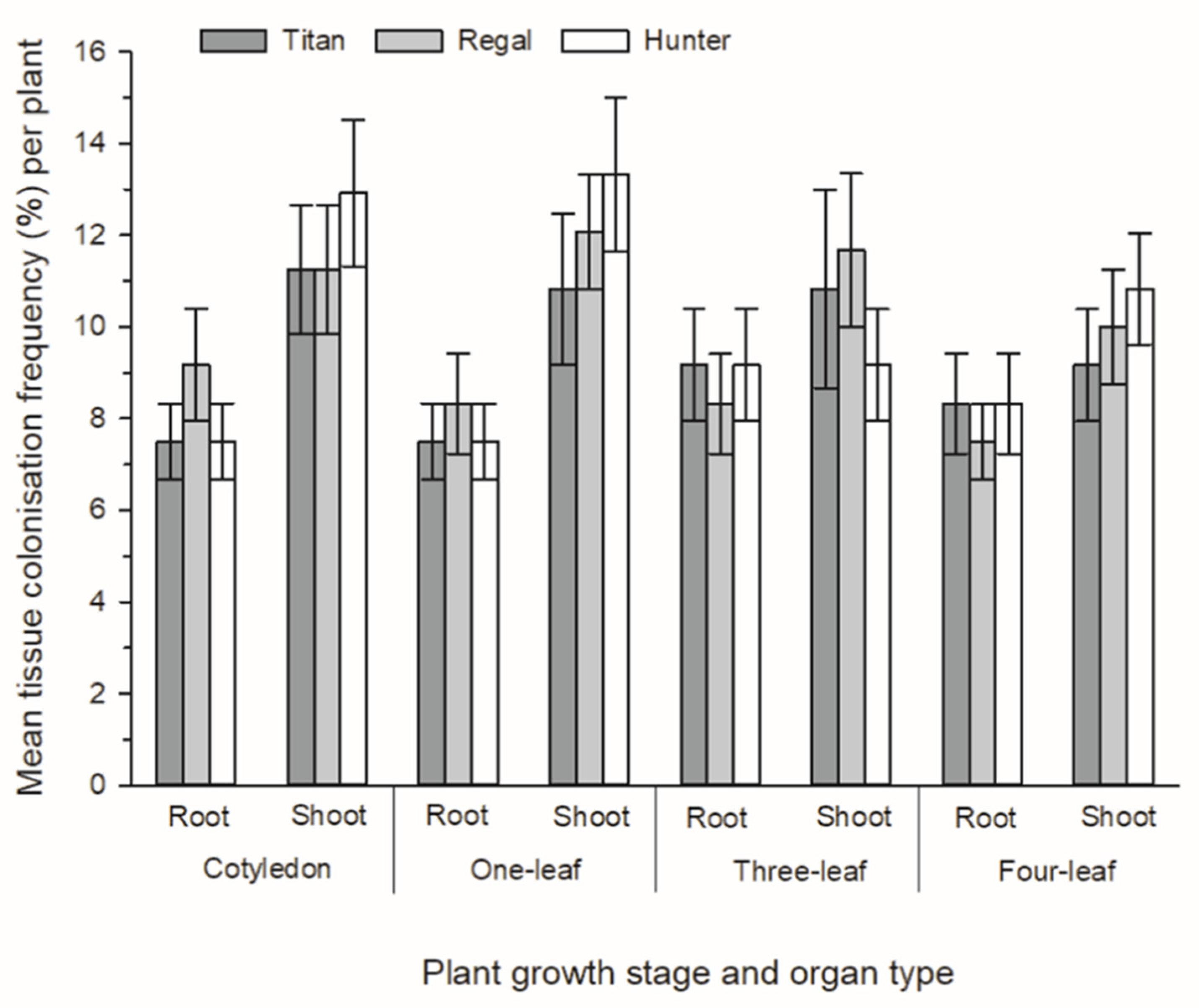
3.2. Development of Novel Plant–Endophyte Associations
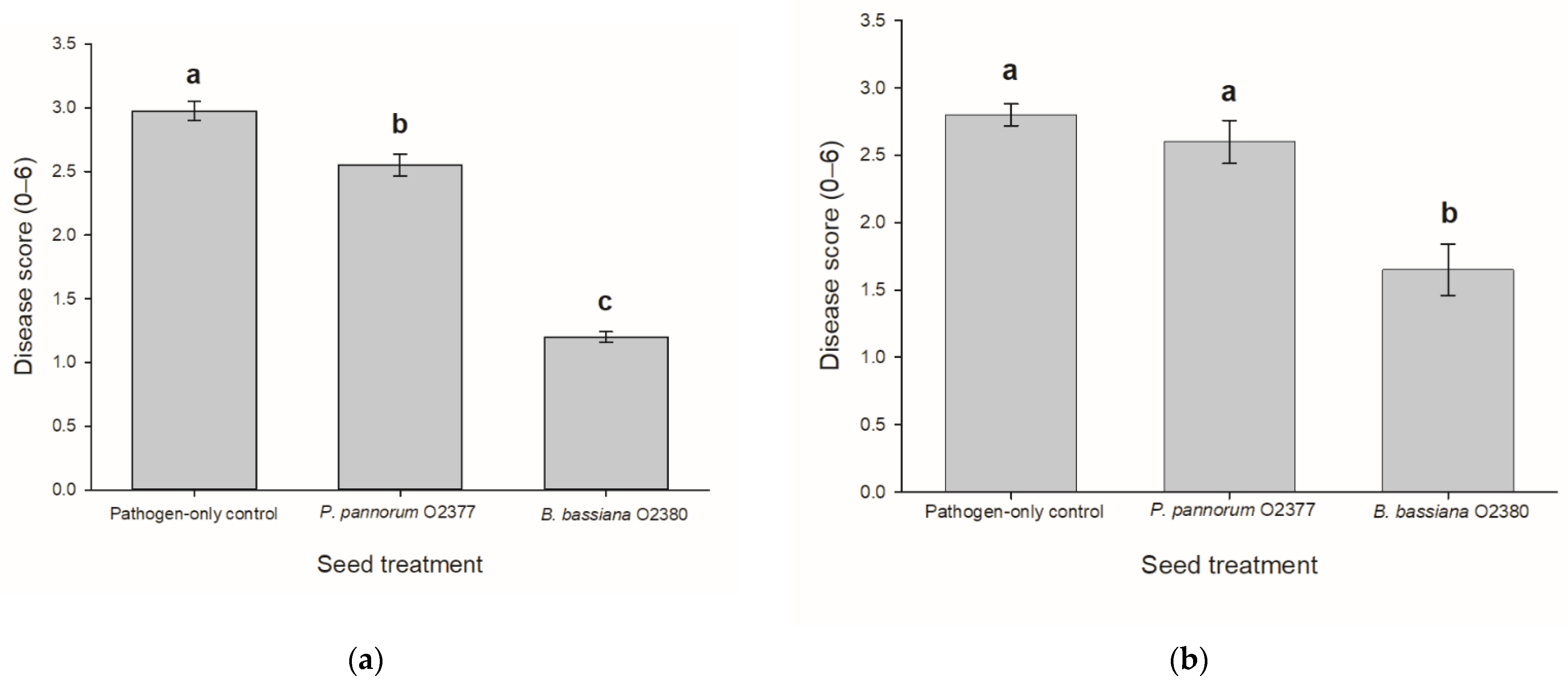
3.3. Bioactivity of Fungal Endophytes towards Leptosphaeria Maculans
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Snowdon, R.; Lühs, W.; Friedt, W. Oilseed Rape. In Genome Mapping and Molecular Breeding in Plants—Oilseeds; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 54–114. [Google Scholar]
- Dixon, G.R. Vegetable Brassicas and Related Crucifers; CABI: Wallingford, UK, 2007. [Google Scholar]
- Gómez-Campo, C. Morphology and morpho-taxonomy of the tribe Brassiceae. In Brassica crops and Wild Allies. Biology and Breeding; Tsunoda, S., Hinata, K., Gomez-Campo, C., Eds.; Japan Scientific Societies Press: Tokyo, Japan, 1980; pp. 3–31. [Google Scholar]
- Warwick, S.I.; Francis, A.; Gugel, R.K. Guide to Wild Germplasm: Brassica and Allied Crops (Tribe Brassiceae, Brassicaceae); Agriculture and Agri-Food Canada: Ottawa, ON, Canada, 2009; pp. 1–6.
- Rao, S.; Horn, F. Cereals and Brassicas for forage. In Forages: An Introduction to Grassland Agriculture; Barnes, R., Miller, D., Nelson, C., Eds.; Iowa State University Press: Ames, IA, USA, 1995; Volume 5, pp. 451–462. [Google Scholar]
- Rakow, G. Species origin and economic importance of Brassica. In Brassica. Biotechnology in Agriculture and Forestry; Pua, E.C., Douglas, C.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; Volume 54, pp. 3–11. [Google Scholar]
- USDA. World Agricultural Production; USDA: Washington, DC, USA, 2019; p. 33.
- Lamb, R.J. Entomology of oilseed Brassica crops. Annu. Rev. Entomol. 1989, 34, 211–229. [Google Scholar] [CrossRef]
- Dixelius, C.; Bohman, S.; Wretblad, S. Disease Resistance. In Biotechnology in Agriculture and Forestry; Douglas, C., Pua, E., Eds.; Brassica; Springer: Berlin/Heidelberg, Germany, 2004; Volume 54, pp. 253–271. [Google Scholar]
- Kimber, D.; McGregor, D. Brassica Oilseeds: Production and Utilization; CAB International: Wallingford, UK, 1995. [Google Scholar]
- West, J.S.; Kharbanda, P.D.; Barbetti, M.J.; Fitt, B.D.L. Epidemiology and management of Leptosphaeria maculans (phoma stem canker) on oilseed rape in Australia, Canada and Europe. Plant Pathol. 2001, 50, 10–27. [Google Scholar] [CrossRef] [Green Version]
- Salisbury, P.; Ballinger, D.; Wratten, N.; Plummer, K.; Howlett, B. Blackleg disease on oilseed Brassica in Australia: A review. Aust. J. Exp. Agric. 1995, 35, 665–672. [Google Scholar] [CrossRef]
- Fitt, B.D.; Brun, H.; Barbetti, M.; Rimmer, S. World-wide importance of phoma stem canker (Leptosphaeria maculans and L. biglobosa) on oilseed rape (Brassica napus). In Sustainable Strategies for Managing Brassica napus (Oilseed Rape) Resistance to Leptosphaeria maculans (Phoma Stem Canker); Springer: Berlin/Heidelberg, Germany, 2006; pp. 3–15. [Google Scholar]
- Fitt, B.D.; Hu, B.; Li, Z.; Liu, S.; Lange, R.; Kharbanda, P.; Butterworth, M.; White, R. Strategies to prevent spread of Leptosphaeria maculans (phoma stem canker) onto oilseed rape crops in China; costs and benefits. Plant Pathol. 2008, 57, 652–664. [Google Scholar] [CrossRef]
- Zhang, X.; Fernando, W.D. Insights into fighting against blackleg disease of Brassica napus in Canada. Crop Pasture Sci. 2018, 69, 40–47. [Google Scholar] [CrossRef]
- Lawrence, F. Agrichemicals and Ever More Intensive Farming Will not Feed the World. The Guardian 2016. Available online: https://www.theguardian.com/commentisfree/2016/oct/02/agrichemicals-intensive-farming-food-production-biodiversity (accessed on 5 April 2021).
- Keswani, C.; Singh, H.B.; Hermosa, R.; García-Estrada, C.; Caradus, J.; He, Y.-W.; Mezaache-Aichour, S.; Glare, T.R.; Borriss, R.; Vinale, F.; et al. Antimicrobial secondary metabolites from agriculturally important fungi as next biocontrol agents. Appl. Microbiol. Biotechnol. 2019, 103, 9287–9303. [Google Scholar] [CrossRef]
- Stokstad, E. European Union Expands Ban of Three Neonicotinoid Pesticides. ScienceInsider 2018. Available online: https://www.science.org/content/article/european-union-expands-ban-three-neonicotinoid-pesticides (accessed on 19 November 2020).
- Neik, T.X.; Barbetti, M.J.; Batley, J. Current status and challenges in identifying disease resistance genes in Brassica napus. Front. Plant Sci. 2017, 8, 1788. [Google Scholar] [CrossRef]
- Card, S.; Johnson, L.; Teasdale, S.; Caradus, J. Deciphering endophyte behaviour: The link between endophyte biology and efficacious biological control agents. FEMS Microbiol. Ecol. 2016, 92, fiw114. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, J.L.; Maccheroni Jr, W.; Pereira, J.O.; de Araújo, W.L. Endophytic microorganisms: A review on insect control and recent advances on tropical plants. Electron. J. Biotechnol. 2000, 3, 15–16. [Google Scholar] [CrossRef]
- Card, S.D.; Hume, D.E.; Roodi, D.; McGill, C.R.; Millner, J.P.; Johnson, R.D. Beneficial endophytic microorganisms of Brassica–A review. Biol. Control 2015, 90, 102–112. [Google Scholar] [CrossRef]
- Finkel, O.M.; Castrillo, G.; Herrera Paredes, S.; Salas González, I.; Dangl, J.L. Understanding and exploiting plant beneficial microbes. Curr. Opin. Plant Biol. 2017, 38, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Roodi, D.; Millner, J.P.; McGill, C.; Johnson, R.D.; Jauregui, R.; Card, S.D. Methylobacterium, a major component of the culturable bacterial endophyte community of wild Brassica seed. PeerJ 2020, 8, e9514. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Ali, H.; Ali, Z.; Ali, H.; Mehmood, S.; Ali, W. In vitro regeneration of Brassica napus L., cultivars (Star, Cyclone and Westar) from hypocotyls and cotyledonary leaves. Pak. J. Bot. 2007, 39, 1251. [Google Scholar]
- Card, S.D. Biological Control of Botrytis cinerea in Lettuce & Strawberry Crops. Ph.D. Thesis, Lincoln University, Canterbury, New Zealand, 2005. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Sanger, F.; Coulson, A.R. A rapid method for determining sequences in DNA by primed synthesis with DNA polymerase. J. Mol. Biol. 1975, 94, 441–448. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2019. [Google Scholar]
- Lenth, R.V. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 1.6.1. 2021. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 17 December 2020).
- Lob, S. Leptosphaeria Diseases of Oilseed Rape and Swede: Identification and Epidemiology. Ph.D. Thesis, Lincoln University, Canterbury, New Zealand, 2014. [Google Scholar]
- Hammoudi, O.; Salman, M.; Abuamsha, R.; Ehlers, R.-U. Effectiveness of bacterial and fungal isolates to control Phoma lingam on oilseed rape Brassica napus. Am. J. Plant Sci. 2012, 3, 773. [Google Scholar] [CrossRef] [Green Version]
- Christensen, R.H.B. Ordinal—Regression Models for Ordinal Data. R Package Version 2019.12-10. 2019. Available online: https://CRAN.R-project.org/package=ordinal (accessed on 6 October 2021).
- Gianni, C.; Caretta, G.; Romano, C. Skin infection due to Geomyces pannorum var. pannorum. Mycoses 2003, 46, 430–432. [Google Scholar] [CrossRef] [PubMed]
- Verant, M.L.; Boyles, J.G.; Waldrep, W., Jr.; Wibbelt, G.; Blehert, D.S. Temperature-dependent growth of Geomyces destructans, the fungus that causes bat white-nose syndrome. PLoS ONE 2012, 7, e46280. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, V.; Springer, D.J.; Behr, M.J.; Ramani, R.; Li, X.; Peck, M.K.; Ren, P.; Bopp, D.J.; Wood, B.; Samsonoff, W.A. Morphological and molecular characterizations of psychrophilic fungus Geomyces destructans from New York bats with white nose syndrome (WNS). PLoS ONE 2010, 5, e10783. [Google Scholar] [CrossRef]
- Marshall, W. Aerial transport of keratinaceous substrate and distribution of the fungus Geomyces pannorum in Antarctic soils. Microb. Ecol. 1998, 36, 212–219. [Google Scholar] [CrossRef]
- Sazanova, K.V.; Senik, S.V.; Kirtsideli, I.Y.; Shavarda, A.L. Metabolomic profiling and lipid composition of Arctic and Antarctic strains of micromycetes Geomyces pannorum and Thelebolus microsporus grown at different temperatures. Microbiology 2019, 88, 282–291. [Google Scholar] [CrossRef]
- Vohník, M.; Fendrych, M.; Albrechtová, J.; Vosátka, M. Intracellular colonization of Rhododendron and Vaccinium roots by Cenococcum geophilum, Geomyces pannorum and Meliniomyces variabilis. Folia Microbiol. 2007, 52, 407–414. [Google Scholar] [CrossRef]
- Domsch, K.H.; Gams, W.; Anderson, T.-H. Compendium of Soil Fungi; Academic Press Ltd.: London, UK, 1980; Volume 1. [Google Scholar]
- Lacourt, I.; Girlanda, M.; Perotto, S.; Del Pero, M.; Zuccon, D.; Luppi, A.M. Nuclear ribosomal sequence analysis of Oidiodendron: Towards a redefinition of ecologically relevant species. New Phytol. 2001, 149, 565–576. [Google Scholar] [CrossRef]
- Rosa, L.H.; Almeida Vieira, M.d.L.; Santiago, I.F.; Rosa, C.A. Endophytic fungi community associated with the dicotyledonous plant Colobanthus quitensis (Kunth) Bartl.(Caryophyllaceae) in Antarctica. FEMS Microbiol. Ecol. 2010, 73, 178–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohse, R.; Jakobs-Schönwandt, D.; Vidal, S.; Patel, A.V. Evaluation of new fermentation and formulation strategies for a high endophytic establishment of Beauveria bassiana in oilseed rape plants. Biol. Control 2015, 88, 26–36. [Google Scholar] [CrossRef]
- Akello, J.; Dubois, T.; Coyne, D.; Kyamanywa, S. Endophytic Beauveria bassiana in banana (Musa spp.) reduces banana weevil (Cosmopolites sordidus) fitness and damage. Crop Prot. 2008, 27, 1437–1441. [Google Scholar] [CrossRef]
- Jaber, L.R.; Enkerli, J. Effect of seed treatment duration on growth and colonization of Vicia faba by endophytic Beauveria bassiana and Metarhizium brunneum. Biol. Control 2016, 103, 187–195. [Google Scholar] [CrossRef]
- Gautam, S.; Mohankumar, S.; Kennedy, J. Induced host plant resistance in cauliflower by Beauveria bassiana. J. Entomol. Zool. Stud. 2016, 4, 476–482. [Google Scholar]
- Posada, F.; Vega, F.E. Establishment of the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales) as an endophyte in cocoa seedlings (Theobroma cacao). Mycologia 2005, 97, 1195–1200. [Google Scholar] [CrossRef]
- Ownley, B.H.; Griffin, M.R.; Klingeman, W.E.; Gwinn, K.D.; Moulton, J.K.; Pereira, R.M. Beauveria bassiana: Endophytic colonization and plant disease control. J. Invertebr. Pathol. 2008, 98, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Rondot, Y.; Reineke, A. Endophytic Beauveria bassiana in grapevine Vitis vinifera (L.) reduces infestation with piercing-sucking insects. Biol. Control 2018, 116, 82–89. [Google Scholar] [CrossRef]
- Bing, L.A.; Lewis, L.C. Suppression of Ostrinia nubilalis (Hübner)(Lepidoptera: Pyralidae) by endophytic Beauveria bassiana (Balsamo) Vuillemin. Environ. Entomol. 1991, 20, 1207–1211. [Google Scholar] [CrossRef]
- Russo, M.L.; Scorsetti, A.C.; Vianna, M.F.; Cabello, M.; Ferreri, N.; Pelizza, S. Endophytic effects of Beauveria bassiana on corn (Zea mays) and its herbivore, Rachiplusia nu (Lepidoptera: Noctuidae). Insects 2019, 10, 110. [Google Scholar] [CrossRef] [Green Version]
- Lefort, M.-C.; McKinnon, A.; Nelson, T.L.; Glare, T. Natural occurrence of the entomopathogenic fungi Beauveria bassiana as a vertically transmitted endophyte of Pinus radiata and its effect on above-and below-ground insect pests. N. Z. Plant Prot. 2016, 69, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Tefera, T.; Vidal, S. Effect of inoculation method and plant growth medium on endophytic colonization of sorghum by the entomopathogenic fungus Beauveria bassiana. BioControl 2009, 54, 663–669. [Google Scholar] [CrossRef] [Green Version]
- Kasambala Donga, T.; Vega, F.E.; Klingen, I. Establishment of the fungal entomopathogen Beauveria bassiana as an endophyte in sugarcane, Saccharum officinarum. Fungal Ecol. 2018, 35, 70–77. [Google Scholar] [CrossRef]
- Qayyum, M.A.; Wakil, W.; Arif, M.J.; Sahi, S.T.; Dunlap, C.A. Infection of Helicoverpa armigera by endophytic Beauveria bassiana colonizing tomato plants. Biol. Control 2015, 90, 200–207. [Google Scholar] [CrossRef]
- Sánchez-Rodríguez, A.R.; Raya-Díaz, S.; Zamarreño, Á.M.; García-Mina, J.M.; del Campillo, M.C.; Quesada-Moraga, E. An endophytic Beauveria bassiana strain increases spike production in bread and durum wheat plants and effectively controls cotton leafworm (Spodoptera littoralis) larvae. Biol. Control 2018, 116, 90–102. [Google Scholar] [CrossRef]
- Vega, F.E. Insect pathology and fungal endophytes. J. Invertebr. Pathol. 2008, 98, 277–279. [Google Scholar] [CrossRef]
- Koch, E.; Zink, P.; Ullrich, C.I.; Kleespies, R.G. Light microscopic studies on the development of Beauveria bassiana and other putative endophytes in leaf tissues. J. Kult. 2018, 70, 95–107. [Google Scholar]
- Mascarin, G.M.; Jaronski, S.T. The production and uses of Beauveria bassiana as a microbial insecticide. World J. Microbiol. Biotechnol. 2016, 32, 177. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, A.C.; Saari, S.; Moran-Diez, M.E.; Meyling, N.V.; Raad, M.; Glare, T.R. Beauveria bassiana as an endophyte: A critical review on associated methodology and biocontrol potential. BioControl 2017, 62, 1–17. [Google Scholar] [CrossRef]
- Zimmermann, G. Review on safety of the entomopathogenic fungi Beauveria bassiana and Beauveria brongniartii. Biocontrol Sci. Technol. 2007, 17, 553–596. [Google Scholar] [CrossRef]
- Steinhaus, E.A. Disease in a Minor Chord: Being a Semihistorical and Semibiographical Account of a Period in Science When One Could Be Happily yet Seriously Concerned with the Diseases of Lowly Animals without Backbones, Especially the Insects; The Ohio State University Press: Columbus, OH, USA, 1975. [Google Scholar]
- Barra-Bucarei, L.; González, M.G.; Iglesias, A.F.; Aguayo, G.S.; Peñalosa, M.G.; Vera, P.V. Beauveria bassiana multifunction as an endophyte: Growth promotion and biologic control of Trialeurodes vaporariorum, (Westwood) (Hemiptera: Aleyrodidae) in tomato. Insects 2020, 11, 591. [Google Scholar] [CrossRef]
- Li, Z.; Li, C.; Huang, B.; Fan, M. Discovery and demonstration of the teleomorph of Beauveria bassiana (Bals.) Vuill., an important entomogenous fungus. Chin. Sci. Bull. 2001, 46, 751–753. [Google Scholar] [CrossRef]
- Jaber, L.R.; Enkerli, J. Fungal entomopathogens as endophytes: Can they promote plant growth? Biocontrol Sci. Technol. 2017, 27, 28–41. [Google Scholar] [CrossRef]
- Khonsanit, A.; Luangsa-ard, J.J.; Thanakitpipattana, D.; Noisripoom, W.; Chaitika, T.; Kobmoo, N. Cryptic diversity of the genus Beauveria with a new species from Thailand. Mycol. Prog. 2020, 19, 291–315. [Google Scholar] [CrossRef]
- Gasmi, L.; Baek, S.; Kim, J.C.; Kim, S.; Lee, M.R.; Park, S.E.; Shin, T.Y.; Lee, S.J.; Parker, B.L.; Kim, J.S. Gene diversity explains variation in biological features of insect killing fungus, Beauveria bassiana. Sci. Rep. 2021, 11, 91. [Google Scholar] [CrossRef]
- Pus, W. Plant-Mediated Effects of Trichoderma spp. and Beauveria bassiana Isolates on Insect and Pathogen Resistance. Master’s Thesis, Lincoln University, Lincoln, New Zealand, 2017. [Google Scholar]
- Tomilova, O.G.; Shaldyaeva, E.M.; Kryukova, N.A.; Pilipova, Y.V.; Schmidt, N.S.; Danilov, V.P.; Kryukov, V.Y.; Glupov, V.V. Entomopathogenic fungi decrease rhizoctonia disease in potato in field conditions. PeerJ 2020, 8, e9895. [Google Scholar] [CrossRef] [PubMed]
- Culebro-Ricaldi, J.; Ruíz-Valdiviezo, V.; Rodríguez-Mendiola, M.A.; Avila Miranda, M.E.; Gutiérrez-Miceli, F.; Cruz-Rodríguez, R.I.; Dendooven, L.; Montes-Molina, J. Antifungal properties of Beauveria bassiana strains against Fusarium oxysporum f. Sp. Lycopersici race 3 in tomato crop. J. Environ. Biol. 2017, 38, 821–827. [Google Scholar] [CrossRef]
- Bark, Y.G.; Lee, D.G.; Kang, S.; Kim, Y. Antibiotic properties of an entomopathogenic fungus, Beauveria bassiana, on Fusarium oxysporum and Botrytis cinerea. Korean J. Plant Pathol. 1996, 12, 245–250. [Google Scholar]
- Griffin, M.; Ownley, B.; Klingeman, W.; Pereira, R. Evidence of induced systemic resistance with Beauveria bassiana against Xanthomonas in cotton. Phytopathology 2006, 96, S42. [Google Scholar]
- Xu, Y.; Orozco, R.; Wijeratne, E.K.; Gunatilaka, A.L.; Stock, S.P.; Molnár, I. Biosynthesis of the cyclooligomer depsipeptide beauvericin, a virulence factor of the entomopathogenic fungus Beauveria bassiana. Chem. Biol. 2008, 15, 898–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fetzer, A.; Müller, K.; Schmid, M.; Eisner, P. Rapeseed proteins for technical applications: Processing, isolation, modification and functional properties—A review. Ind. Crop. Prod. 2020, 158, 112986. [Google Scholar] [CrossRef]
- Van de Wouw, A.P.; Elliott, V.L.; Chang, S.; López-Ruiz, F.J.; Marcroft, S.J.; Idnurm, A. Identification of isolates of the plant pathogen Leptosphaeria maculans with resistance to the triazole fungicide fluquinconazole using a novel in planta assay. PLoS ONE 2017, 12, e0188106. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Akhavan, A.; Hwang, S.-F.; Strelkov, S.E. Decreased sensitivity of Leptosphaeria maculans to pyraclostrobin in Alberta, Canada. Plant Dis. 2020, 104, 2462–2468. [Google Scholar] [CrossRef]
- Parsa, S.; Ortiz, V.; Vega, F.E. Establishing fungal entomopathogens as endophytes: Towards endophytic biological control. J. Vis. Exp. 2013, 74, 50360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaber, L.R.; Ownley, B.H. Can we use entomopathogenic fungi as endophytes for dual biological control of insect pests and plant pathogens? Biol. Control 2018, 116, 36–45. [Google Scholar] [CrossRef]
- Butt, T.M.; Coates, C.J.; Dubovskiy, I.M.; Ratcliffe, N.A. Chapter Nine—Entomopathogenic Fungi: New Insights into Host–Pathogen Interactions. In Advances in Genetics; Lovett, B., St. Leger, R.J., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 94, pp. 307–364. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roodi, D.; Millner, J.P.; McGill, C.R.; Johnson, R.D.; Hea, S.-Y.; Brookes, J.J.; Glare, T.R.; Card, S.D. Development of Plant–Fungal Endophyte Associations to Suppress Phoma Stem Canker in Brassica. Microorganisms 2021, 9, 2387. https://doi.org/10.3390/microorganisms9112387
Roodi D, Millner JP, McGill CR, Johnson RD, Hea S-Y, Brookes JJ, Glare TR, Card SD. Development of Plant–Fungal Endophyte Associations to Suppress Phoma Stem Canker in Brassica. Microorganisms. 2021; 9(11):2387. https://doi.org/10.3390/microorganisms9112387
Chicago/Turabian StyleRoodi, Davood, James P. Millner, Craig R. McGill, Richard D. Johnson, Shen-Yan Hea, Jenny J. Brookes, Travis R. Glare, and Stuart D. Card. 2021. "Development of Plant–Fungal Endophyte Associations to Suppress Phoma Stem Canker in Brassica" Microorganisms 9, no. 11: 2387. https://doi.org/10.3390/microorganisms9112387
APA StyleRoodi, D., Millner, J. P., McGill, C. R., Johnson, R. D., Hea, S. -Y., Brookes, J. J., Glare, T. R., & Card, S. D. (2021). Development of Plant–Fungal Endophyte Associations to Suppress Phoma Stem Canker in Brassica. Microorganisms, 9(11), 2387. https://doi.org/10.3390/microorganisms9112387