
Antimicrobial Activity of Chemically and Biologically Treated Chitosan Prepared from Black Soldier Fly (Hermetia illucens) Pupal Shell Waste


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Chemical Extraction of Chitin from Hermetia illucens Pupal Exuviae
2.3. Biological Extraction of Chitin from Hermetia illucens Pupal Exuviae
2.3.1. Bacteria Isolates
2.3.2. Biological Extraction
2.3.3. Deproteinization and Demineralization
2.4. Deacetylation of Chitin to Chitosan
2.5. Determination of Percentage Yield of Chitin and Chitosan
2.6. Characterization of Chitin and Chitosan by Fourier-Transform Infrared (FTIR) Spectroscopy
2.7. Characterisation of Chitin and Chitosan by Scanning Electron Microscopy (SEM)
2.8. Antimicrobial Activities
2.8.1. Test Organisms for Antimicrobial Assay
2.8.2. Inoculum and Sample Preparation
2.8.3. Antimicrobial Susceptibility Assay
2.9. Data Collection and Analysis
3. Results and Discussions
3.1. Chitin Yield from the Pupal Exuviae of the Black Soldier Fly
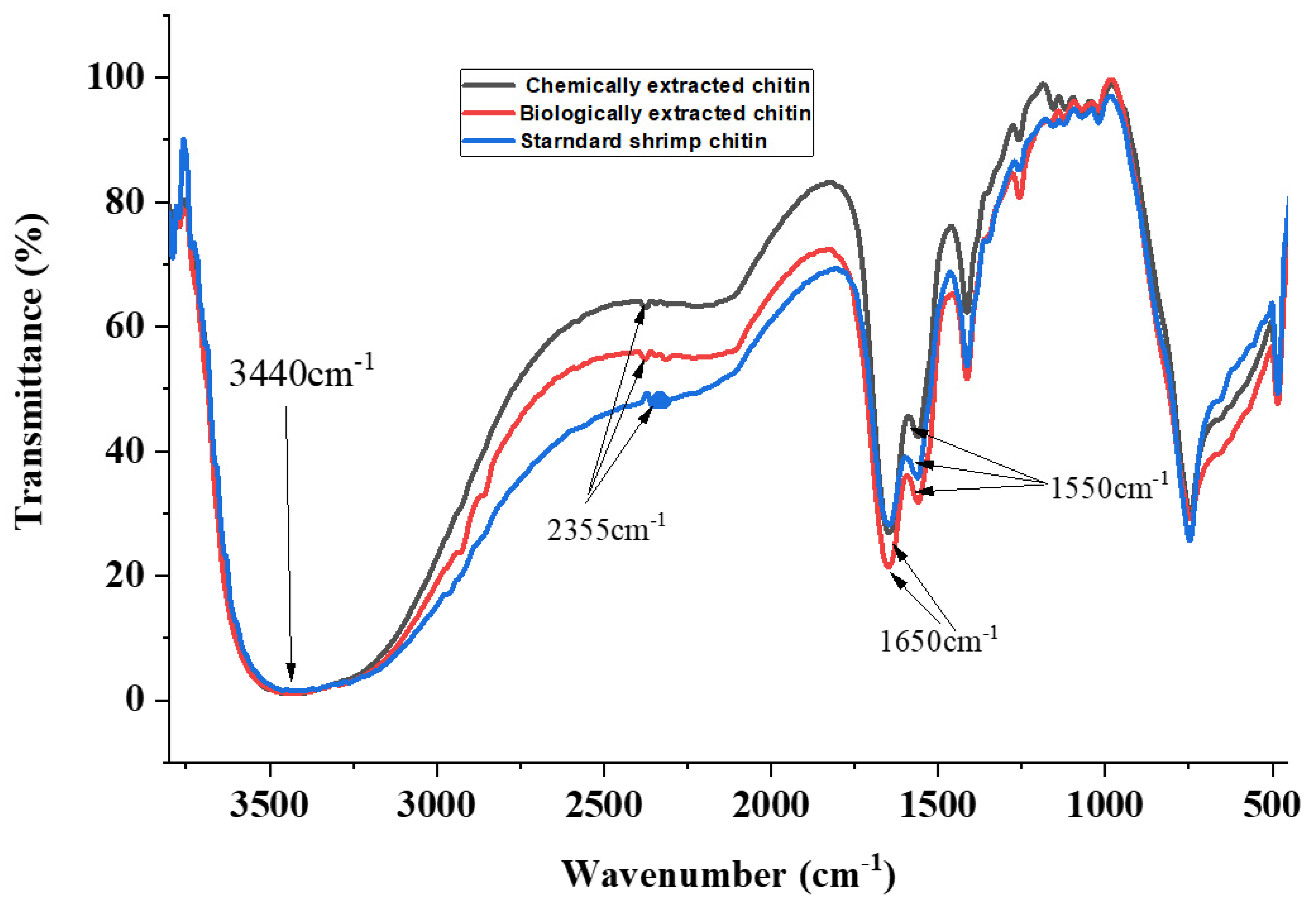
3.2. Analysis of Functional Groups in Chitin Extracted from the Black Soldier Fly
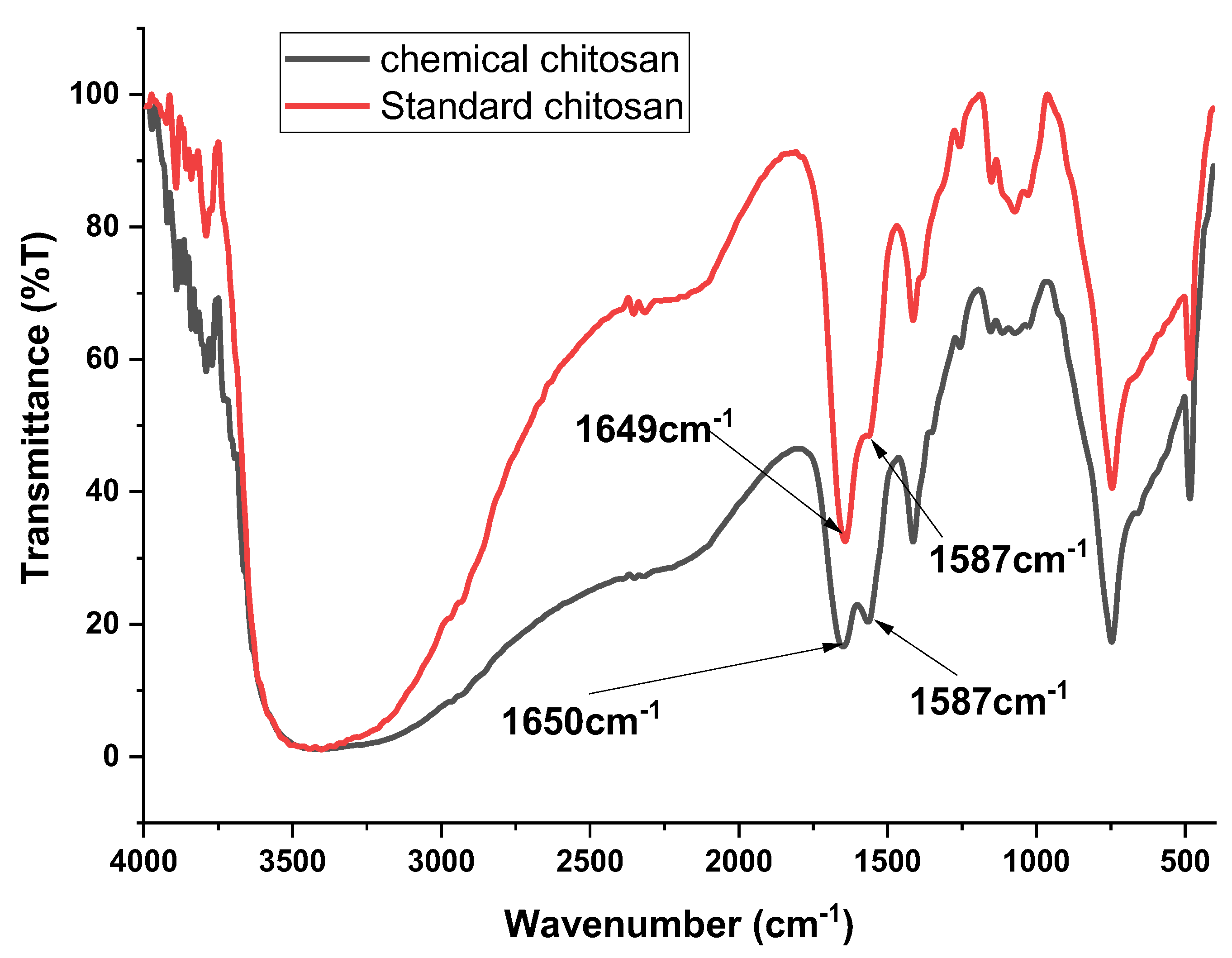
3.3. Analysis of Functional Groups in Chitosan Extracted from Black Soldier Fly
3.4. Characterization of Chitin and Chitosan by Scanning Electron Microscopy (SEM)
3.5. Antimicrobial Properties of Chitosan against Pathogenic Microbes
3.6. Comparative Effects of the Antimicrobial Activity of Chitosan from BSF, Shrimp, and Conventional Antibiotics against Selected Pathogenic Microbes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, M.; Rajput, M.; Soni, T.; Vivekanand, V.; Pareek, N. Chemoenzymatic production and engineering of chitooligosaccharides and N-acetyl glucosamine for refining biological activities. Front. Chem. 2020, 8, 469. [Google Scholar] [CrossRef]
- Joseph, S.M.; Krishnamoorthy, S.; Paranthaman, R.; Moses, J.; Anandharamakrishnan, C. A review on source-specific chemistry, functionality, and applications of chitin and chitosan. Carbohydr. Polym. Technol. Appl. 2021, 2, 100036. [Google Scholar] [CrossRef]
- Nechita, P. Applications of chitosan in wastewater treatment. Biol. Act. Appl. Mar. Polysacch. 2017, 1, 209–228. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Sader, K.; Meramo-Hurtado, S.; González-Delgado, A. Environmental sustainability analysis of chitosan microbeads production for pharmaceutical applications via computer-aided simulation, WAR and TRACI assessments. Sustain. Chem. Pharm. 2020, 15, 100212. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, Y.; Zhou, F.; Guo, J.; Tang, J.; Han, Y.; Li, Z.; Fu, C. Preparation, bioactivities and applications in food industry of chitosan-based maillard products: A review. Molecules 2020, 26, 166. [Google Scholar] [CrossRef]
- Shahrajabian, M.; Chaski, C.; Polyzos, N.; Tzortzakis, N.; Petropoulos, S. Sustainable agriculture systems in vegetable production using chitin and chitosan as plant biostimulants. Biomolecules 2021, 11, 819. [Google Scholar] [CrossRef] [PubMed]
- El Knidri, H.; Belaabed, R.; Addaou, A.; Laajeb, A.; Lahsini, A. Extraction, chemical modification and characterization of chitin and chitosan. Int. J. Biol. Macromol. 2018, 120, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Pachapur, V.L.; Guemiza, K.; Rouissi, T.; Sarma, S.J.; Brar, S.K. Novel biological and chemical methods of chitin extraction from crustacean waste using saline water. J. Chem. Technol. Biotechnol. 2016, 91, 2331–2339. [Google Scholar] [CrossRef] [Green Version]
- Zainol Abidin, N.A.; Kormin, F.; Zainol Abidin, N.A.; Mohamed Anuar, N.A.F.; Abu Bakar, M.F. The potential of insects as alternative sources of chitin: An overview on the chemical method of extraction from various sources. Int. J. Mol. Sci. 2020, 21, 4978. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.N.; Lee, P.P.; Chen, W.N. Microbial extraction of chitin from seafood waste using sugars derived from fruit waste-stream. AMB Express 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chai, J.; Hang, J.; Zhang, C.; Yang, J.; Wang, S.; Liu, S.; Fang, Y. Purification and characterization of chitin deacetylase active on insoluble chitin from Nitratireductor aquimarinus MCDA3-3. Int. J. Biol. Macromol. 2020, 152, 922–929. [Google Scholar] [CrossRef]
- Achur, R.N. Isolation and characterization of chitin from Millipede (Spirobolida). J. Basic Appl. Zool. 2018, 79, 30. [Google Scholar] [CrossRef] [Green Version]
- Marmier, T.; Szczepanski, C.R.; Candet, C.; Zenerino, A.; Godeau, R.-P.; Godeau, G. Investigation on Mecynorhina torquata Drury, 1782 (Coleoptera, Cetoniidae, Goliathini) cuticle: Surface properties, chitin and chitosan extraction. Int. J. Biol. Macromol. 2020, 164, 1164–1173. [Google Scholar] [CrossRef]
- Mohan, K.; Ganesan, A.R.; Muralisankar, T.; Jayakumar, R.; Sathishkumar, P.; Uthayakumar, V.; Chandirasekar, R.; Revathi, N. Recent insights into the extraction, characterization, and bioactivities of chitin and chitosan from insects. Trends Food Sci. Technol. 2020, 105, 17–42. [Google Scholar] [CrossRef] [PubMed]
- Vetter, J. Chitin content of cultivated mushrooms Agaricus bisporus, Pleurotus ostreatus and Lentinula edodes. Food Chem. 2007, 102, 6–9. [Google Scholar] [CrossRef]
- Ospina Álvarez, S.P.; Ramírez Cadavid, D.A.; Escobar Sierra, D.M.; Ossa Orozco, C.P.; Rojas Vahos, D.F.; Zapata Ocampo, P.; Atehortúa, L. Comparison of extraction methods of chitin from Ganoderma lucidum mushroom obtained in submerged culture. BioMed Res. Int. 2014, 169071. [Google Scholar] [CrossRef] [Green Version]
- Hassainia, A.; Satha, H.; Boufi, S. Chitin from Agaricus bisporus: Extraction and characterization. Int. J. Biol. Macromol. 2018, 117, 1334–1342. [Google Scholar] [CrossRef] [PubMed]
- Shumo, M.; Osuga, I.M.; Khamis, F.M.; Tanga, C.M.; Fiaboe, K.K.M.; Subramanian, S.; Ekesi, S.; Van Huis, A.; Borgemeister, C. The nutritive value of black soldier fly larvae reared on common organic waste streams in Kenya. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Zimri, M.N. Preparation and Electrospinning of Chitosan from Waste Black Soldier Fly Biomass; University of the Western Cape: Cape Town, South Africa, 2018. [Google Scholar]
- Aytekin, O.; Elibol, M. Cocultivation of Lactococcus lactis and Teredinobacter turnirae for biological chitin extraction from prawn waste. Bioprocess. Biosyst. Eng. 2009, 33, 393–399. [Google Scholar] [CrossRef]
- Kaur, S.; Dhillon, G.S. Recent trends in biological extraction of chitin from marine shell wastes: A review. Crit. Rev. Biotechnol. 2015, 35, 44–61. [Google Scholar] [CrossRef]
- Akram, K.; Ahn, J.-J.; Kwon, J.-H. Identification and characterization of gamma-irradiated dried Lentinus edodes using ESR, SEM, and FTIR analyses. J. Food Sci. 2012, 77, C690–C696. [Google Scholar] [CrossRef] [PubMed]
- Kourmouli, A.; Valenti, M.; Van Rijn, E.; Beaumont, H.J.E.; Kalantzi, O.-I.; Schmidt-Ott, A.; Biskos, G. Can disc diffusion susceptibility tests assess the antimicrobial activity of engineered nanoparticles? J. Nano. Res. 2018, 20, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spranghers, T.; Ottoboni, M.; Klootwijk, C.; Ovyn, A.; Deboosere, S.; De Meulenaer, B.; Michiels, J.; Eeckhout, M.; De Clercq, P.; De Smet, S. Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. J. Sci. Food Agric. 2017, 97, 2594–2600. [Google Scholar] [CrossRef] [PubMed]
- Caligiani, A.; Marseglia, A.; Leni, G.; Baldassarre, S.; Maistrello, L.; Dossena, A.; Sforza, S. Composition of black soldier fly prepupae and systematic approaches for extraction and fractionation of proteins, lipids and chitin. Food Res. Int. 2018, 105, 812–820. [Google Scholar] [CrossRef]
- Wong, C.-Y.; Rosli, S.S.; Uemura, Y.; Ho, Y.C.; Leejeerajumnean, A.; Kiatkittipong, W.; Cheng, C.-K.; Lam, M.-K.; Lim, J.-W. Potential protein and biodiesel sources from black soldier fly larvae: Insights of larval harvesting instar and fermented feeding medium. Energies 2019, 12, 1570. [Google Scholar] [CrossRef] [Green Version]
- Złotko, K.; Waśko, A.; Kamiński, D.; Budziak-Wieczorek, I.; Bulak, P.; Bieganowski, A. Isolation of chitin from black soldier fly (Hermetia illucens) and its usage to metal sorption. Polymers 2021, 13, 818. [Google Scholar] [CrossRef]
- Hajji, S.; Younes, I.; Ghorbel-Bellaaj, O.; Hajji, R.; Rinaudo, M.; Nasri, M.; Jellouli, K. Structural differences between chitin and chitosan extracted from three different marine sources. Int. J. Biol. Macromol. 2014, 65, 298–306. [Google Scholar] [CrossRef]
- Hahn, T.; Tafi, E.; Paul, A.; Salvia, R.; Falabella, P.; Zibek, S. Current state of chitin purification and chitosan production from insects. J. Chem. Technol. Biotechnol. 2020, 95, 2775–2795. [Google Scholar] [CrossRef]
- Erdogan, S.; Kaya, M. High similarity in physicochemical properties of chitin and chitosan from nymphs and adults of a grasshopper. Int. J. Biol. Macromol. 2016, 89, 118–126. [Google Scholar] [CrossRef]
- Marei, N.H.; El-Samie, E.A.; Salah, T.; Saad, G.R.; Elwahy, A.H. Isolation and characterization of chitosan from different local insects in Egypt. Int. J. Biol. Macromol. 2016, 82, 871–877. [Google Scholar] [CrossRef]
- Sargin, I.; Arslan, G.; Kaya, M. Production of magnetic chitinous microcages from ephippia of zooplankton Daphnia longispina and heavy metal removal studies. Carbohydr. Polym. 2019, 207, 200–210. [Google Scholar] [CrossRef]
- Hossain, M.; Iqbal, A. Production and characterization of chitosan from shrimp waste. J. Bangladesh Agric. Univ. 2014, 12, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.-W.; Han, Y.S.; Jo, Y.H.; Choi, M.H.; Kang, S.H.; Kim, S.-A.; Jung, W.-J. Extraction of chitin and chitosan from housefly, Musca domestica, pupa shells. Entomol. Res. 2016, 46, 324–328. [Google Scholar] [CrossRef]
- Yuan, Y.; Chen, L.; Shi, W.; Wang, Z.; Zhang, H. Insight into physicochemical, rheological, and antibacterial properties of chitosan extracted from Antarctic krill: A comparative study. Molecules 2020, 25, 4074. [Google Scholar] [CrossRef] [PubMed]
- Kaya, M.; Sofi, K.; Sargin, I.; Mujtaba, M. Changes in physicochemical properties of chitin at developmental stages (larvae, pupa and adult) of Vespa crabro (wasp). Carbohydr. Polym. 2016, 145, 64–70. [Google Scholar] [CrossRef]
- Tsurkan, M.V.; Voronkina, A.; Khrunyk, Y.; Wysokowski, M.; Petrenko, I.; Ehrlich, H. Progress in chitin analytics. Carbohydr. Polym. 2021, 252, 117204. [Google Scholar] [CrossRef]
- Eddya, M.; Tbib, B.; El-Hami, K. A comparison of chitosan properties after extraction from shrimp shells by diluted and concentrated acids. Heliyon 2020, 6, e03486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erdogan, S.; Kaya, M.; Akata, I. Chitin Extraction and Chitosan Production from Cell Wall of Two Mushroom Species (Lactarius vellereus and Phyllophora ribis); AIP Publishing LLC: Melville, NY, USA, 2017; Volume 1809, p. 020012. [Google Scholar]
- Ibitoye, B.E.; Lokman, I.H.; Hezmee, M.N.M.; Goh, Y.-M.; Zuki, A.B.Z.; Jimoh, A.A. Extraction and physicochemical characterization of chitin and chitosan isolated from house cricket. Biomed. Mater. 2017, 13, 025009. [Google Scholar] [CrossRef] [Green Version]
- Al Sagheer, F.; Al-Sughayer, M.; Muslim, S.; Elsabee, M. Extraction and characterization of chitin and chitosan from marine sources in Arabian Gulf. Carbohydr. Polym. 2009, 77, 410–419. [Google Scholar] [CrossRef]
- Kaya, M.; Baran, T.; Erdoğan, S.; Menteş, A.; Özüsağlam, M.A.; Çakmak, Y.S. Physicochemical comparison of chitin and chitosan obtained from larvae and adult Colorado potato beetle (Leptinotarsa decemlineata). Mater. Sci. Eng. C 2014, 45, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Kaya, M.; Erdogan, S.; Mol, A.; Baran, T. Comparison of chitin structures isolated from seven Orthoptera species. Int. J. Biol. Macromol. 2015, 72, 797–805. [Google Scholar] [CrossRef]
- Kaya, M.; Halıcı, M.G.; Duman, F.; Erdoğan, S.; Baran, T. Characterisation of α-chitin extracted from a lichenised fungus species Xanthoria parietina. Nat. Prod. Res. 2015, 29, 1280–1284. [Google Scholar] [CrossRef] [PubMed]
- Younes, I.; Rinaudo, M. Chitin and chitosan preparation from marine sources. Structure, properties and applications. Mar. Drugs 2015, 13, 1133–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Rehman, K.U.; Feng, W.; Yang, D.; Rehman, R.U.; Cai, M.; Zhang, J.; Yu, Z.; Zheng, L. Physicochemical structure of chitin in the developing stages of black soldier fly. Int. J. Biol. Macromol. 2020, 149, 901–907. [Google Scholar] [CrossRef]
- Kaya, M.; Baran, T.; Asan-Ozusaglam, M.; Cakmak, Y.S.; Tozak, K.O.; Mol, A.; Menteş, A.; Sezen, G. Extraction and characterization of chitin and chitosan with antimicrobial and antioxidant activities from cosmopolitan Orthoptera species (Insecta). Biotechnol. Bioprocess. Eng. 2015, 20, 168–179. [Google Scholar] [CrossRef]
- Leke-Aladekoba, A.A. Comparison of Extraction Methods and Characterisation of Chitin and Chitosan with Antimicrobial and Antioxidant Properties from Black Soldier Fly (Hermetia illucens) Meal. Master’s Thesis, Dalhousie University, Nova Scotia, Canada, 2018. [Google Scholar]
- Al-Nabulsi, A.; Osaili, T.; Sawalha, A.; Olaimat, A.N.; Albiss, B.A.; Mehyar, G.; Ayyash, M.; Holley, R. Antimicrobial activity of chitosan coating containing ZnO nanoparticles against E. coli O157:H7 on the surface of white brined cheese. Int. J. Food Microbiol. 2020, 334, 108838. [Google Scholar] [CrossRef]
- Alfaifi, M.Y.; Alkabli, J.; Elshaarawy, R.F. Suppressing of milk-borne pathogenic using new water-soluble chitosan-azidopropanoic acid conjugate: Targeting milk-preservation quality improvement. Int. J. Biol. Macromol. 2020, 164, 1519–1526. [Google Scholar] [CrossRef]
- Divya, K.; Vijayan, S.; George, T.K.; Jisha, M.S. Antimicrobial properties of chitosan nanoparticles: Mode of action and factors affecting activity. Fibers Polym. 2017, 18, 221–230. [Google Scholar] [CrossRef]
- Elshaarawy, R.F.; Ismail, L.A.; Alfaifi, M.Y.; Rizk, M.A.; Eltamany, E.; Janiak, C. Inhibitory activity of biofunctionalized silver-capped N-methylated water-soluble chitosan thiomer for microbial and biofilm infections. Int. J. Biol. Macromol. 2020, 152, 709–717. [Google Scholar] [CrossRef]
- Khalil, A.M.; Abdel-Monem, R.A.; Darwesh, O.M.; Hashim, A.I.; Nada, A.A.; Rabie, S.T. Synthesis, characterization, and evaluation of antimicrobial activities of chitosan and carboxymethyl chitosan schiff-base/silver nanoparticles. J. Chem. 2017, 2017, 1434320. [Google Scholar] [CrossRef]
- Umoren, S.A.; Solomon, M.M.; Nzila, A.; Obot, I.B. Preparation of silver/chitosan nanofluids using selected plant extracts: Characterization and antimicrobial studies against gram-positive and gram-negative bacteria. Materials 2020, 13, 1629. [Google Scholar] [CrossRef] [Green Version]
- Dash, S.; Kumar, M.; Pareek, N. Enhanced antibacterial potential of berberine via synergism with chitosan nanoparticles. Mater. Today Proc. 2020, 31, 640–645. [Google Scholar] [CrossRef]
- Namangkalakul, W.; Benjavongkulchai, S.; Pochana, T.; Promchai, A.; Satitviboon, W.; Howattanapanich, S.; Phuprasong, R.; Ungvijanpunya, N.; Supakanjanakanti, D.; Chaitrakoonthong, T.; et al. Activity of chitosan antifungal denture adhesive against common Candida species and Candida albicans adherence on denture base acrylic resin. J. Prosthet. Dent. 2020, 123, 181.e1–181.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.H.; Younus, H.; Allemailem, K.S.; Almatroudi, A.; Alrumaihi, F.; Alruwetei, A.M.; Alsahli, M.A.; Khan, A.; Alam Khan, M. Potential of methylglyoxal-conjugated chitosan nanoparticles in treatment of fluconazole-resistant Candida albicans infection in a murine model. Int. J. Nanomed. 2020, 15, 3681–3693. [Google Scholar] [CrossRef]
- Choi, W.H.; Yun, J.H.; Chu, J.P.; Chu, K.B. Antibacterial effect of extracts of Hermetia illucens (Diptera Stratiomyidae) larvae against G ram-negative bacteria. Entomol. Res. 2012, 42, 219–226. [Google Scholar] [CrossRef]
- Hipalaswins, W.M.; Balakumaran, M.; Jagadeeswari, S. Synthesis, characterization and antibacterial activity of chitosan nanoparticles and its impact on seed germination. J. Acad. Ind. Res. 2016, 5, 65. [Google Scholar]
- Chandrasekaran, M.; Kim, K.D.; Chun, S.C. Antibacterial activity of chitosan nanoparticles: A review. Processes 2020, 8, 1173. [Google Scholar] [CrossRef]
- Silva, C.; Matamá, T.; Kim, S.; Padrão, J.; Prasetyo, E.N.; Kudanga, T.; Nyanhongo, G.S.; Guebitz, G.M.; Casal, M.; Cavaco-Paulo, A. Antimicrobial and antioxidant linen via laccase-assisted grafting. React. Funct. Polym. 2011, 71, 713–720. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, O.M.; El-Baghdady, K.Z.; Khalil, M.; El Borhamy, M.I.; Meligi, G.A. Antibacterial, antibiofilm and cytotoxic activities of biogenic polyvinyl alcohol-silver and chitosan-silver nanocomposites. J. Polym. Res. 2020, 27, 1–9. [Google Scholar] [CrossRef]
- Kipkoech, C.; Kinyuru, J.N.; Imathiu, S.; Meyer-Rochow, V.B.; Roos, N. In vitro study of cricket chitosan’s potential as a prebiotic and a promoter of probiotic microorganisms to control pathogenic bacteria in the human gut. Foods 2021, 10, 2310. [Google Scholar] [CrossRef]
- Shapi’I, R.A.; Othman, S.H.; Nordin, N.; Basha, R.K.; Naim, M.N. Antimicrobial properties of starch films incorporated with chitosan nanoparticles: In vitro and in vivo evaluation. Carbohydr. Polym. 2020, 230, 115602. [Google Scholar] [CrossRef] [PubMed]
- Homaeigohar, S.; Boccaccini, A.R. Antibacterial biohybrid nanofibers for wound dressings. Acta Biomater. 2020, 107, 25–49. [Google Scholar] [CrossRef] [PubMed]
- Youssef, K.; Hashim, A.F. Inhibitory effect of clay/chitosan nanocomposite against Penicillium digitatum on citrus and its possible mode of action. Jordan J. Biol. Sci. 2020, 13, 349–355. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chitin Extraction Treatments | Chitin Yield(g) (Mean ± S.D) |
|---|---|
| B. subtilis + L. plantarum | 7.78 ± 0.68 a |
| P. aeruginosa + L. plantarum | 9.47 ± 0.52 b |
| L.plantarum + B. subtilis | 8.76 ± 0.88 b |
| L.plantarum+P.aeruginosa | 7.99 ± 1.16 a |
| P. aeruginosa + B. subtilis +L. plantarum. | 11.85 ± 1.16 c |
| Chemical extraction. | 10.18 ± 0.42 c |
| p-value | <0.001 |
| Bacteria Species | |||||
|---|---|---|---|---|---|
| Concentration (g/mL) | C. albicans | E. coli | S. aureus | B. subtilis | P. aeruginosa |
| 0.5 | 14.33 ± 3.21 b | 14.33 ± 3.21 b | 13.33 ± 1.53 b | 13.67 ± 2.52 b | 14.33 ± 0.58 b |
| 1.0 | 16.67 ± 3.21 b | 20.00 ± 4.36 c | 16.67 ± 2.31 c | 16.67 ± 1.53 c | 16.67 ± 1.15 c |
| 2.5 | 22.00 ± 2.65 c | 25.33 ± 2.89 c | 21.67 ± 4.93 d | 22.67 ± 2.52 d | 18.33 ± 1.53 d |
| 5.0 | 26.00 ± 3.61 c | 26.33 ± 2.89 c | 23.33 ± 3.79 d | 24.67 ± 1.53 d | 20.33 ± 1.53 d |
| 1%Acetic acid (+control) | 13.33 ± 1.46 b | 11.67 ± 1.46 b | 11.67 ± 1.46 a | 10.00 ± 1.46 b | 12.33 ± 1.46 b |
| Sterile distilled water (-control) | 0.00 ± 1.46 a | 0.00 ± 1.46 a | 0.00 ± 1.46 a | 0.00 ± 1.46 a | 0.00 ± 1.46 a |
| p-values | 0.0081 | 0.0087 | 0.0245 | 0.0006 | 0.0023 |
| Bacteria Species | |||||
|---|---|---|---|---|---|
| Antibiotics | C. albicans | E. coli | S. aureus | B. subtilis | P. aeruginosa |
| BSF chitosan | 19.75 ± 5.48 c | 21.5 ± 5.78 d | 18.75 ± 5.07 b | 19.42 ± 4.96 b | 17.42 ± 2.54 b |
| Shrimp chitosan | 16.58 ± 4.19 c | 19.42 ± 5.58 d | 15.92 ± 5.32 b | 18.17 ± 5.02 b | 14.83 ± 3.16 b |
| Ampicillin (25 mcg) | 0.00 ± 1.46 a | 0.00 ± 1.46 a | 0.00 ± 1.46 a | 0.00 ± 1.46 a | 0.00 ± 1.46 a |
| Tetracycline (100 mcg) | 25.00 ± 1.46 c | 20.00 ± 1.46 d | 19.00 ± 1.46 b | 24.00 ± 1.46 c | 20.00 ± 1.46 c |
| Nitrofurantoin (200 mcg) | 15.00 ± 1.46 c | 15.00 ± 1.46 d | 19.00 ± 1.46 b | 18.00 ± 1.46 b | 22.00 ± 1.46 d |
| Nalidixic acid (30 mcg) | 0.00 ± 1.46 a | 27.00 ± 1.46 e | 21.00 ± 1.46 b | 20.00 ± 1.46 b | 20.00 ± 1.46 d |
| Streptomycin (25 mcg) | 15.00 ± 1.46 c | 15.00 ± 1.46 d | 19.00 ± 1.46 b | 0.00 ± 1.46 a | 15.00 ± 1.46 b |
| Sulfathiazole (200 mcg) | 10.00 ± 1.46 b | 10.00 ± 1.46 b | 0.00 ± 1.46 a | 0.00 ± 1.46 a | 0.00 ± 1.46 a |
| Co-trimoxazole (25 mcg) | 17.00 ± 1.46 c | 12.00 ± 1.46 c | 0.00 ± 1.46 a | 0.00 ± 1.46 a | 0.00 ± 1.46 a |
| Gentamycin (10 mcg) | 17.00 ± 1.46 c | 28.00 ± 1.46 e | 29.00 ± 1.46 c | 31.00 ± 1.46 d | 30.00 ± 1.46 e |
| p-Values | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lagat, M.K.; Were, S.; Ndwigah, F.; Kemboi, V.J.; Kipkoech, C.; Tanga, C.M. Antimicrobial Activity of Chemically and Biologically Treated Chitosan Prepared from Black Soldier Fly (Hermetia illucens) Pupal Shell Waste. Microorganisms 2021, 9, 2417. https://doi.org/10.3390/microorganisms9122417
Lagat MK, Were S, Ndwigah F, Kemboi VJ, Kipkoech C, Tanga CM. Antimicrobial Activity of Chemically and Biologically Treated Chitosan Prepared from Black Soldier Fly (Hermetia illucens) Pupal Shell Waste. Microorganisms. 2021; 9(12):2417. https://doi.org/10.3390/microorganisms9122417
Chicago/Turabian StyleLagat, Mevin Kiprotich, Samuel Were, Francis Ndwigah, Violah Jepkogei Kemboi, Carolyne Kipkoech, and Chrysantus Mbi Tanga. 2021. "Antimicrobial Activity of Chemically and Biologically Treated Chitosan Prepared from Black Soldier Fly (Hermetia illucens) Pupal Shell Waste" Microorganisms 9, no. 12: 2417. https://doi.org/10.3390/microorganisms9122417
APA StyleLagat, M. K., Were, S., Ndwigah, F., Kemboi, V. J., Kipkoech, C., & Tanga, C. M. (2021). Antimicrobial Activity of Chemically and Biologically Treated Chitosan Prepared from Black Soldier Fly (Hermetia illucens) Pupal Shell Waste. Microorganisms, 9(12), 2417. https://doi.org/10.3390/microorganisms9122417