
Lactobacillus delbrueckii CRL 581 Differentially Modulates TLR3-Triggered Antiviral Innate Immune Response in Intestinal Epithelial Cells and Macrophages

 , ,
, ,  ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lactobacilli Growth Conditions and Case in Hydrolysates Preparation
2.2. Porcine Intestinal Epithelial Cells and TLR3 Activation
2.3. RT-qPCR in PIE Cells
2.4. Adhesion to PIE Cells
2.5. Adhesion to Porcine Mucins
2.6. Murine Macrophages Cultures
2.7. RT-qPCR in Macrophages
2.8. Statistical Analysis
3. Results
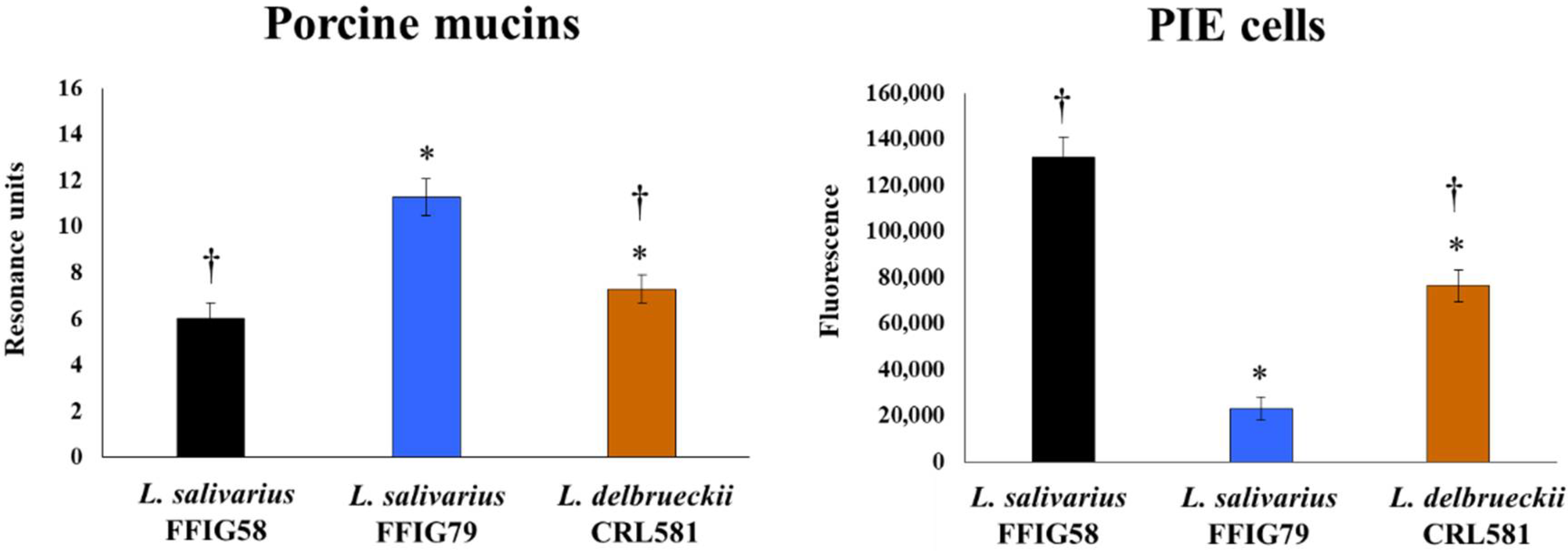
3.1. Adhesion Properties of L. delbrueckii subsp. lactis CRL 581
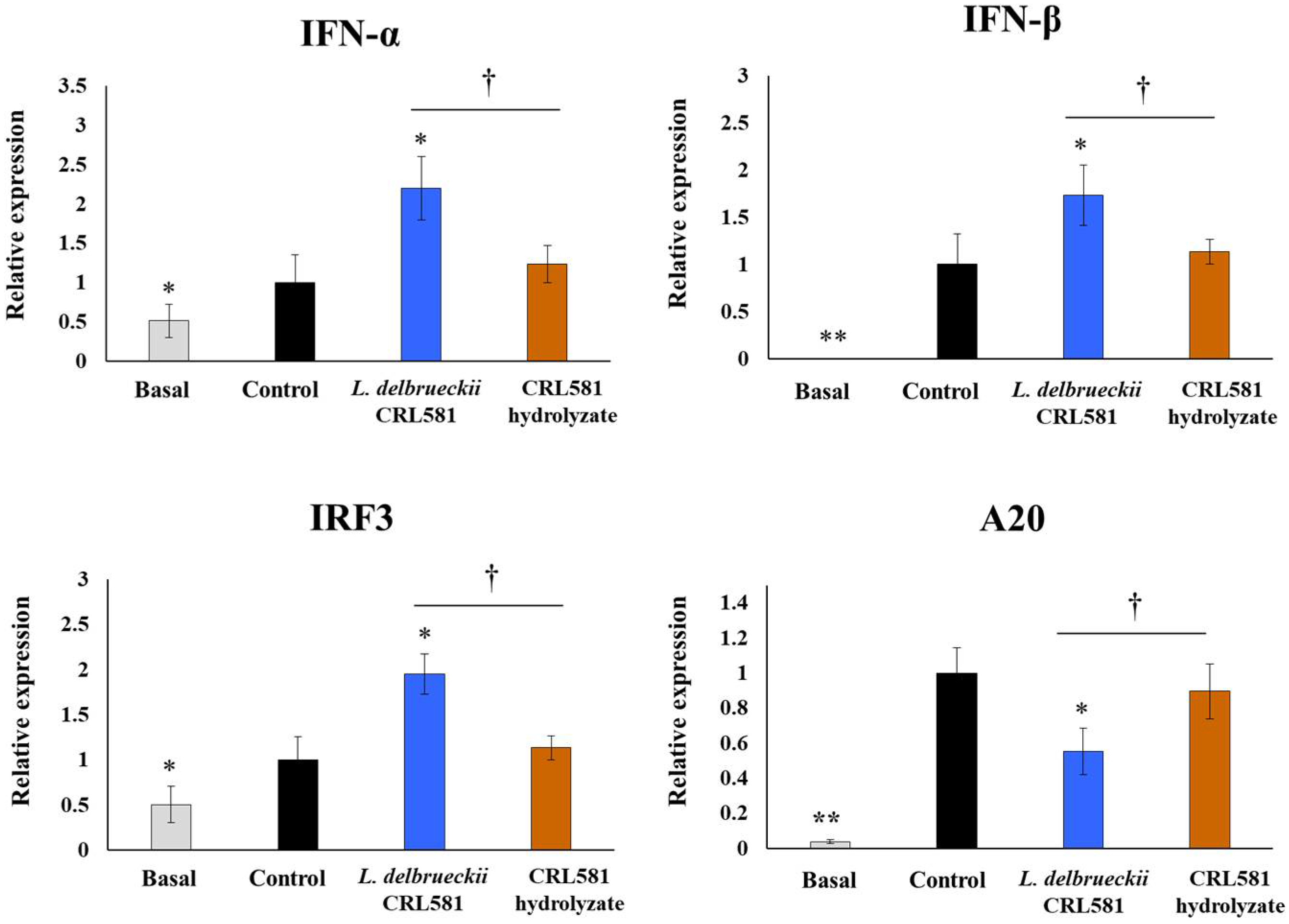
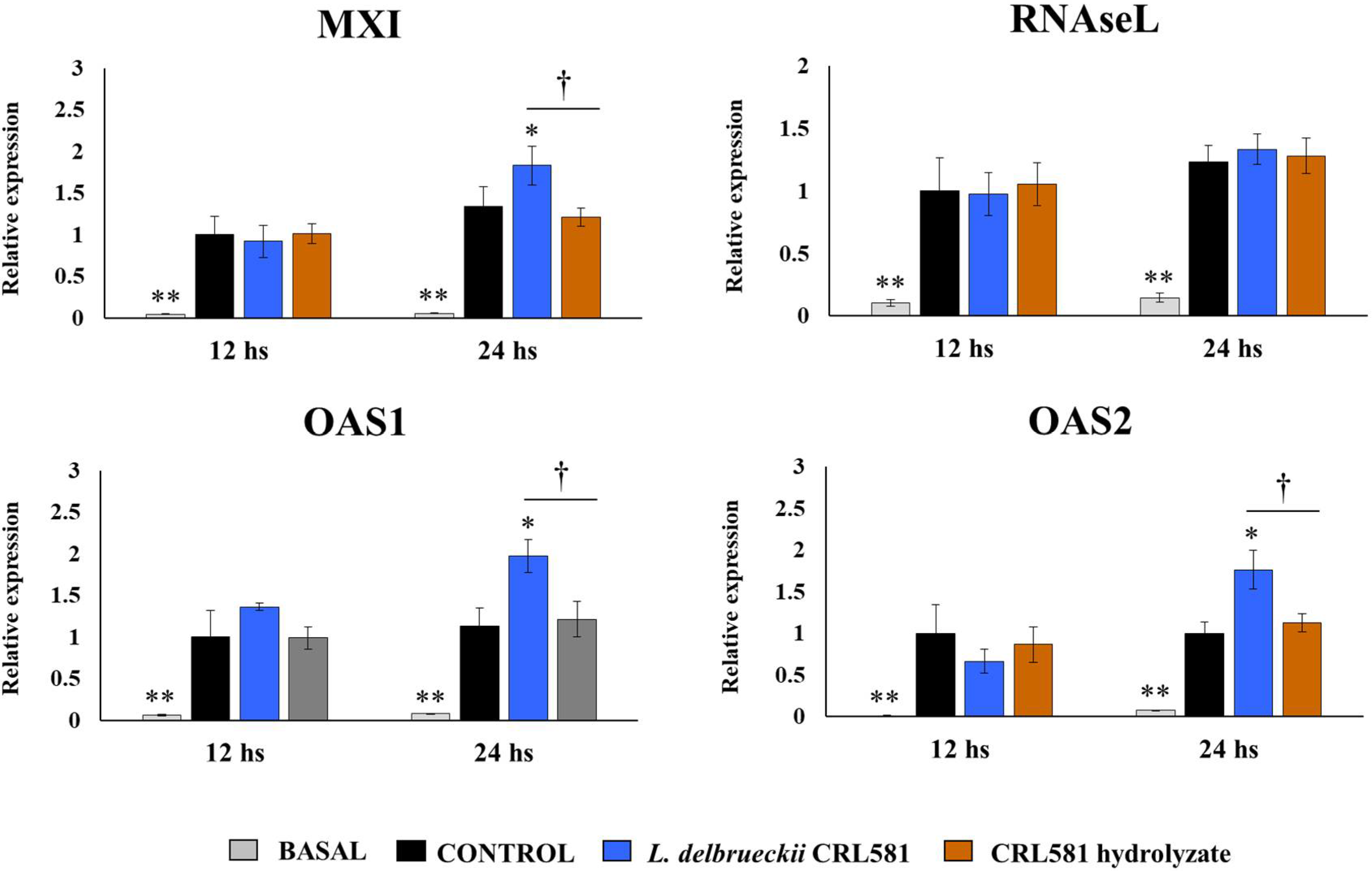
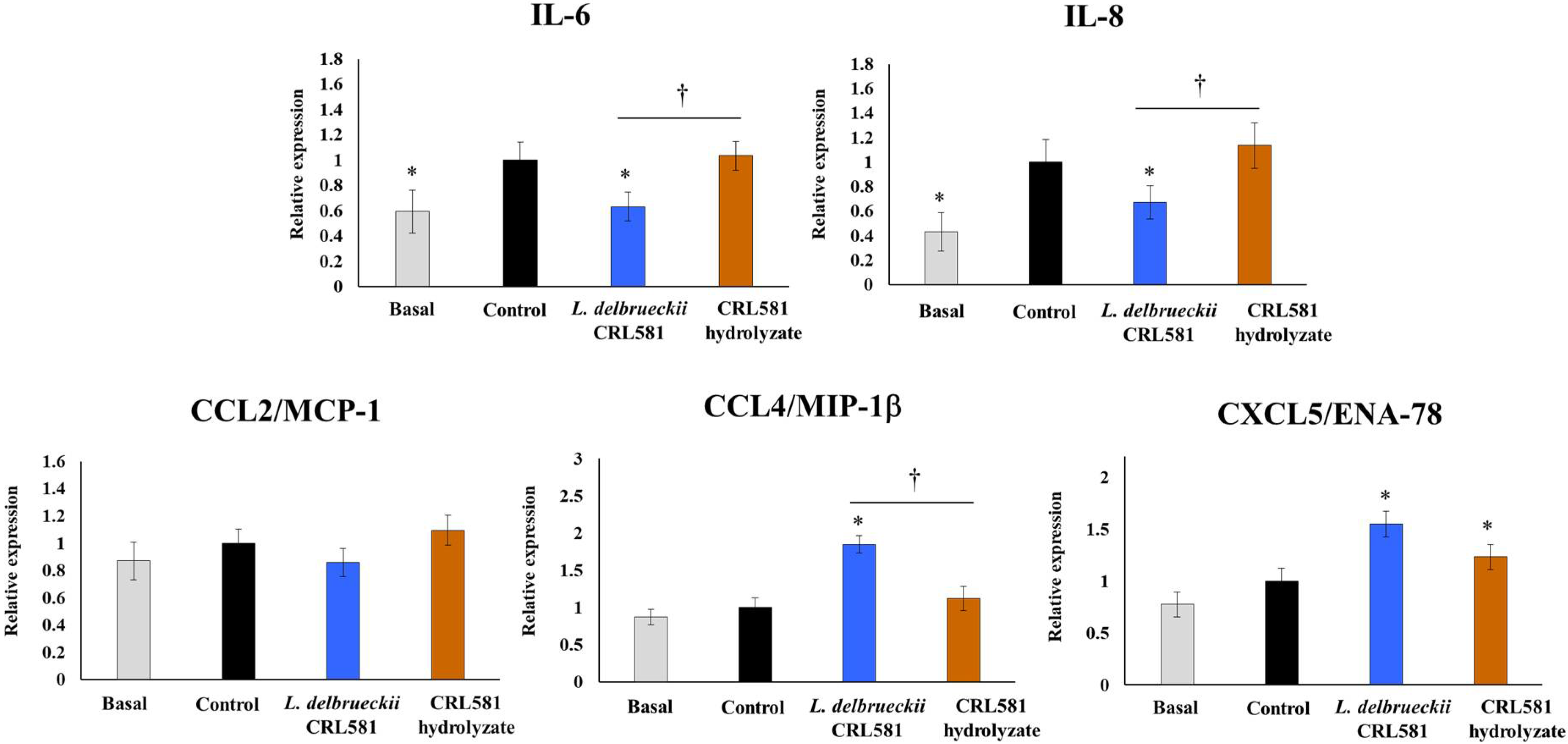
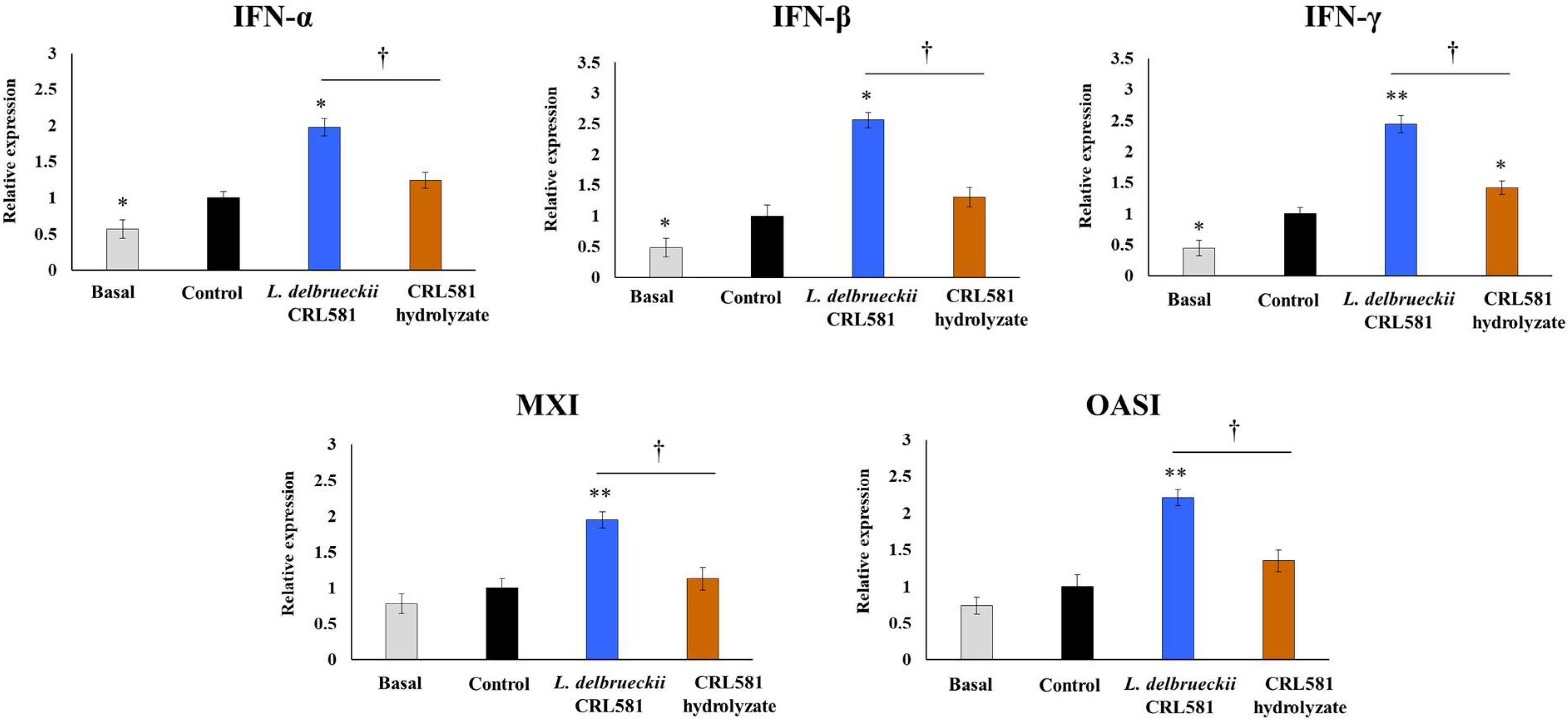
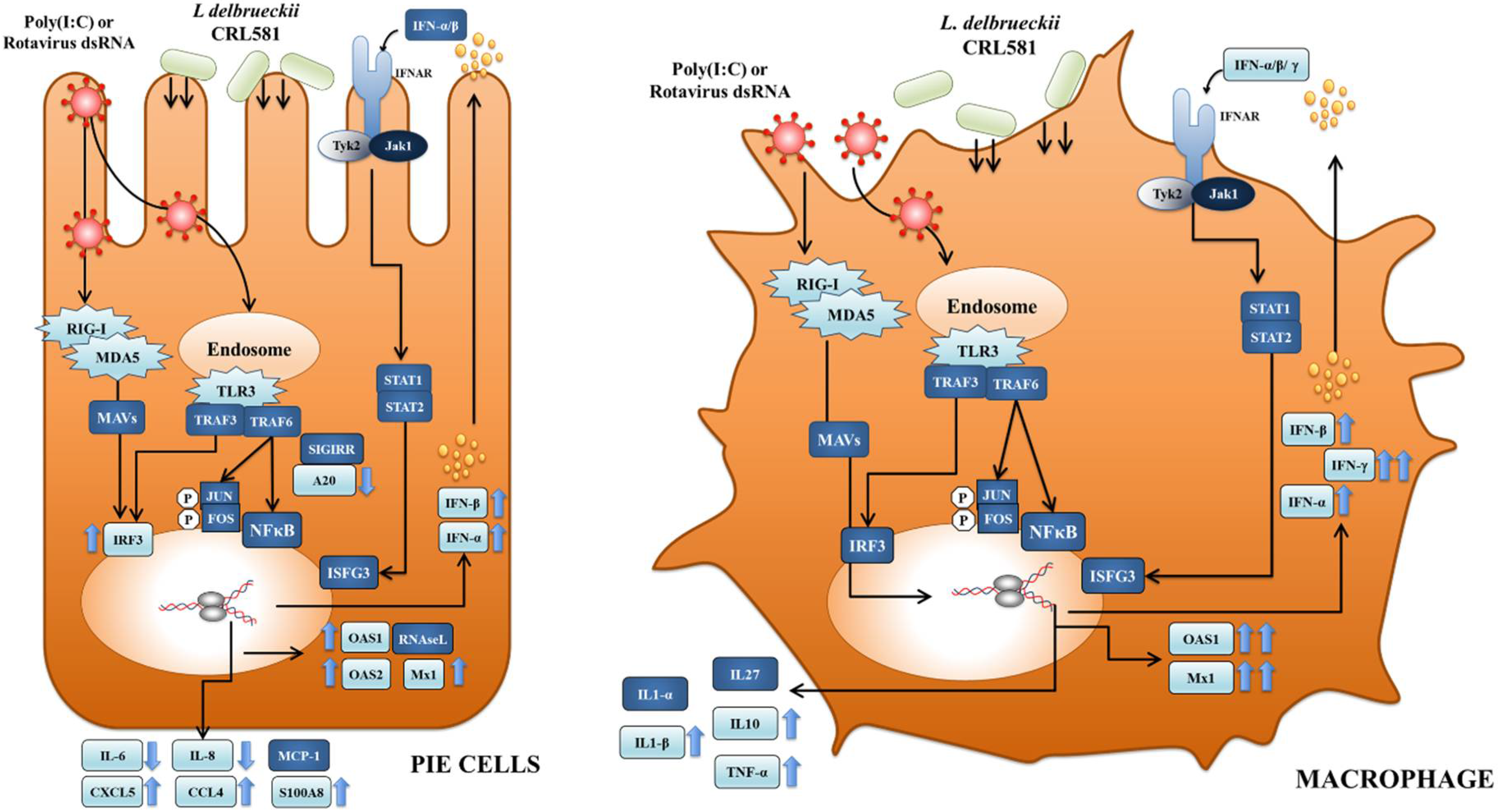
3.2. Immunomodulatory Properties of L. delbrueckii subsp. lactis CRL 581 in PIE Cells
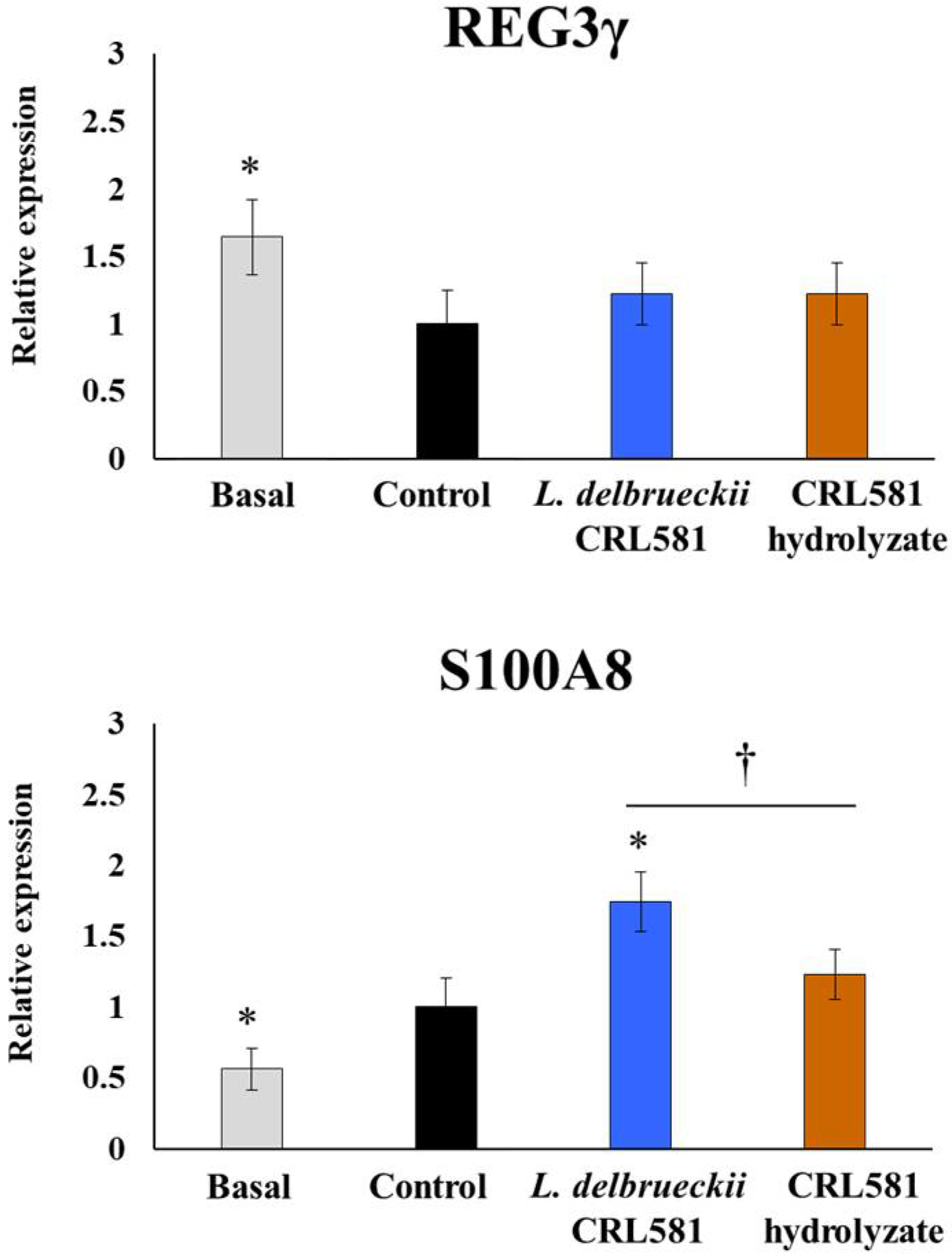
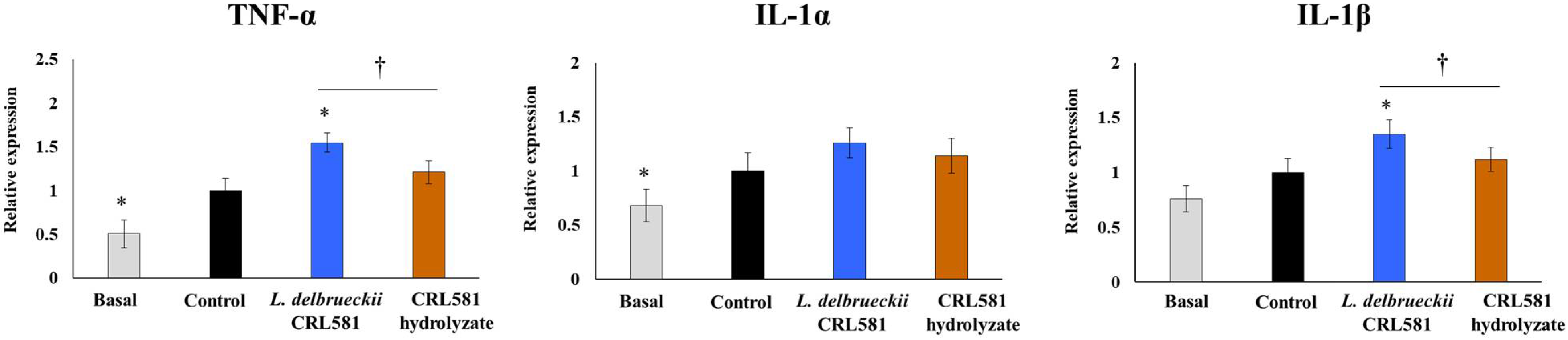
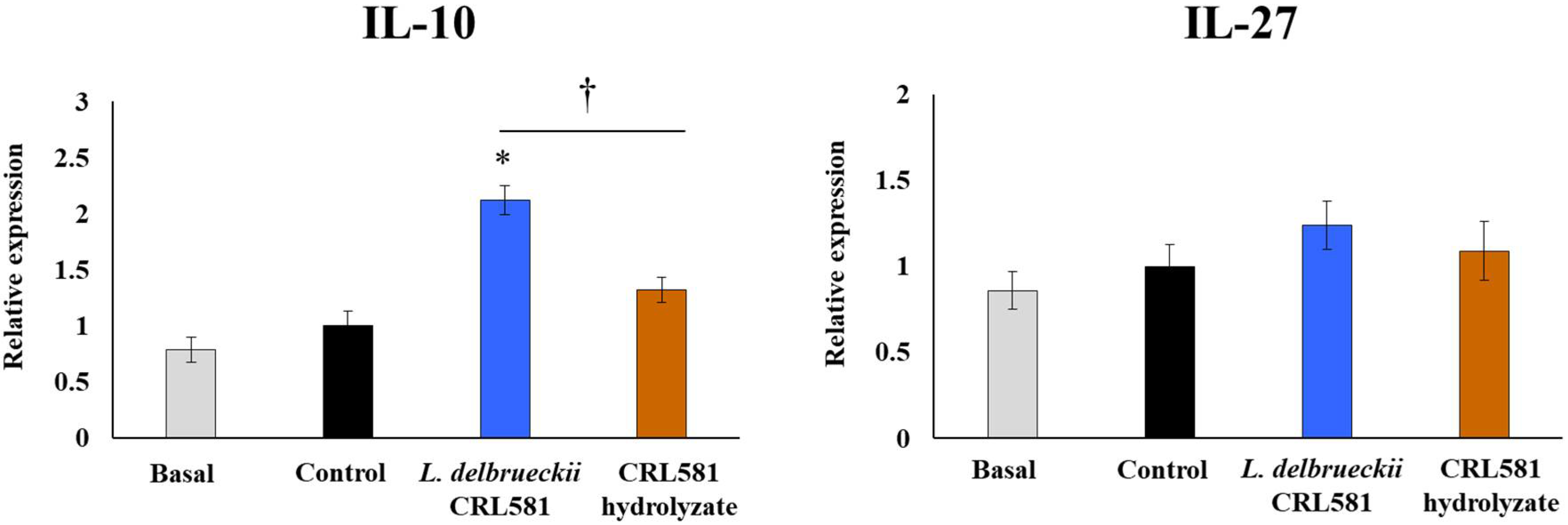
3.3. Immunomodulatory Properties of L. delbrueckii subsp. lactis CRL 581 in Macrophages
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lin, C.-L.; Chen, S.-C.; Liu, S.-Y.; Chen, K.-T. Disease Caused by Rotavirus Infection. Open Virol. J. 2014, 8, 14–19. [Google Scholar] [CrossRef] [Green Version]
- McQuade, E.T.R.; Platts-Mills, J.A. Monitoring the impact of rotavirus vaccines on a global scale. Lancet Glob. Heal. 2019, 7, e817–e818. [Google Scholar] [CrossRef] [Green Version]
- Tate, J.E.; Burton, A.B.; Boschi-Pinto, C.; Parashar, U.D. World Health Organization—Coordinated Global Rotavirus Surveillance Network. Global, regional, and national estimates of rotavirus mortality in children, <5 years of age, 2000–2013. Clin. Infect. Dis. 2016, 62 (Suppl. 2), S96–S105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, K.O.; Kim, Y.; Saif, L.J. Rotavirus and Reovirus. In Diseases of Swine; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; Wiley: New York, NY, USA, 2012; pp. 621–634. [Google Scholar]
- Shepherd, F.K.; Freeman, M.J.; Culhane, M.R.; Marthaler, D.G. Reoviruses (Rotaviruses and Reoviruses). In Diseases of Swine; Wiley: New York, NY, USA, 2019; pp. 715–727. [Google Scholar]
- Gandhi, G.R.; Santos, V.S.; Denadai, M.; Calisto, V.K.D.S.; Quintans, J.; Silva, A.; Araújo, A.A.D.S.; Narain, N.; Cuevas, L.E.; Júnior, L.J.Q.; et al. Cytokines in the management of rotavirus infection: A systematic review of in vivo studies. Cytokine 2017, 96, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, C.A.; Acosta, O. Inflammatory and oxidative stress in rotavirus infection. World J. Virol. 2016, 5, 38–62. [Google Scholar] [CrossRef]
- Di Fiore, I.J.; Holloway, G.; Coulson, B.S. Innate immune responses to rotavirus infection in macrophages depend on MAVS but involve neither the NLRP3 inflammasome nor JNK and p38 signaling pathways. Virus Res. 2015, 208, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Pott, J.; Stockinger, S.; Torow, N.; Smoczek, A.; Lindner, C.; McInerney, G.; Bäckhed, F.; Baumann, U.; Pabst, O.; Bleich, A.; et al. Age-Dependent TLR3 Expression of the Intestinal Epithelium Contributes to Rotavirus Susceptibility. PLoS Pathog. 2012, 8, e1002670. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Wei, H.; Sun, R.; Tian, Z. Recognition of Double-Stranded RNA by TLR3 Induces Severe Small Intestinal Injury in Mice. J. Immunol. 2007, 178, 4548–4556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clancy, R. Immunobiotics and the probiotic evolution. FEMS Immunol. Med. Microbiol. 2003, 38, 9–12. [Google Scholar] [CrossRef] [Green Version]
- Villena, J.; Kitazawa, H. Modulation of Intestinal TLR4-Inflammatory Signaling Pathways by Probiotic Microorganisms: Lessons Learned from Lactobacillus jensenii TL2937. Special issue: Toll-Like receptor activation in immunity vs. tolerance. Front. Immunol. 2014, 4, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Villena, J.; Kitazawa, H. The Modulation of Mucosal Antiviral Immunity by Immunobiotics: Could They Offer Any Benefit in the SARS-CoV-2 Pandemic? Front. Physiol. 2020, 11, 699. [Google Scholar] [CrossRef]
- Villena, J.; Li, C.; Vizoso-Pinto, M.; Sacur, J.; Ren, L.; Kitazawa, H. Lactiplantibacillus plantarum as a Potential Adjuvant and Delivery System for the Development of SARS-CoV-2 Oral Vaccines. Microorganisms 2021, 9, 683. [Google Scholar] [CrossRef]
- Villena, J.; Vizoso-Pinto, M.G.; Kitazawa, H. Intestinal Innate Antiviral Immunity and Immunobiotics: Beneficial Effects against Rotavirus Infection. Special issue: Immunobiotics: Interactions of Beneficial Microbes with the Immune System. Front. Immunol. 2016, 7, 563. [Google Scholar] [CrossRef] [Green Version]
- Brown, L.; Pingitore, E.; Mozzi, F.; Saavedra, L.; Villegas, J.M.; Hebert, E.M. Lactic Acid Bacteria as Cell Factories for the Generation of Bioactive Peptides. Protein Pept. Lett. 2017, 24, 146–155. [Google Scholar] [CrossRef]
- Brown, L.; Villegas, J.M.; Elean, M.; Fadda, S.; Mozzi, F.; Saavedra, L.; Hebert, E.M. YebC, a putative transcriptional factor involved in the regulation of the proteolytic system of Lactobacillus. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Elean, M.; Albarracín, L.; Cataldo, P.; Londero, A.; Kitazawa, H.; Saavedra, L.; Villena, J.; Hebert, E.M. New immunobiotics from highly proteolytic Lactobacillus delbrueckii strains: Their impact on intestinal antiviral innate immune response. Benef. Microbes 2020, 11, 375–390. [Google Scholar] [CrossRef] [PubMed]
- Salazar, M.; Malpeli, A.; Hebert, E.; Cravero, R.; Espeche, W.; Sisnieguez, C.L.; Stavile, R.N.; March, C.; Sisnieguez, C.E.L.; Tavella, M.; et al. Effect of cheese containing Lactobacillus delbrueckii subsp. lactis CRL 581 on blood pressure in prehypertensive and stage 1 hypertensive subjects. Rev. Chil. Nutr. 2019, 46, 701–707. [Google Scholar] [CrossRef] [Green Version]
- Villegas, J.M.; Picariello, G.; Mamone, G.; Turbay, M.B.E.; de Giori, G.S.; Hebert, E.M. Milk-derived angiotensin-I-converting enzymeinhibitory peptides generated by Lactobacillus delbrueckii subsp. lactis CRL 581. Peptidomics 2014, 1, 22–29. [Google Scholar] [CrossRef]
- Zelaya, H.; Alvarez, S.; Kitazawa, H.; Villena, J. Respiratory Antiviral Immunity and Immunobiotics: Beneficial Effects on Inflammation-Coagulation Interaction during Influenza Virus Infection. Special issue: Immunobiotics: Interactions of Beneficial Microbes with the Immune System. Front. Immunol. 2016, 7, 633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albarracin, L.; Kobayashi, H.; Iida, H.; Sato, N.; Nochi, T.; Aso, H.; Salva, S.; Alvarez, S.; Kitazawa, H.; Villena, J. Transcriptomic Analysis of the Innate Antiviral Immune Response in Porcine Intestinal Epithelial Cells: Influence of Immunobiotic Lactobacilli. Front. Immunol. 2017, 8, 57. [Google Scholar] [CrossRef] [Green Version]
- Albarracin, L.; Garcia-Castillo, V.; Masumizu, Y.; Indo, Y.; Islam, A.; Suda, Y.; Garcia-Cancino, A.; Aso, H.; Takahashi, H.; Kitazawa, H.; et al. Efficient Selection of New Immunobiotic Strains with Antiviral Effects in Local and Distal Mucosal Sites by Using Porcine Intestinal Epitheliocytes. Front. Immunol. 2020, 11, 543. [Google Scholar] [CrossRef] [PubMed]
- Nygard, A.-B.; Jørgensen, C.B.; Cirera, S.; Fredholm, M. Selection of reference genes for gene expression studies in pig tissues using SYBR green qPCR. BMC Mol. Biol. 2007, 8, 67. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.; Albarracin, L.; Indo, Y.; Arce, L.; Masumizu, Y.; Tomokiyo, M.; Islam, A.; Garcia-Castillo, V.; Ikeda-Ohtsubo, W.; Nochi, T.; et al. Selection of Immunobiotic Ligilactobacillus salivarius Strains from the Intestinal Tract of Wakame-Fed Pigs: Functional and Genomic Studies. Microorganisms 2020, 8, 1659. [Google Scholar] [CrossRef]
- Clua, P.; Tomokiyo, M.; Tonetti, F.R.; Islam, A.; Castillo, V.G.; Marcial, G.; Salva, S.; Alvarez, S.; Takahashi, H.; Kurata, S.; et al. The Role of Alveolar Macrophages in the Improved Protection against Respiratory Syncytial Virus and Pneumococcal Superinfection Induced by the Peptidoglycan of Lactobacillus rhamnosus CRL1505. Cells 2020, 9, 1653. [Google Scholar] [CrossRef]
- Makino, S.; Ikegami, S.; Kano, H.; Sashihara, T.; Sugano, H.; Horiuchi, H.; Saito, T.; Oda, M. Immunomodulatory Effects of Polysaccharides Produced by Lactobacillus delbrueckii ssp. bulgaricus OLL1073R-1. J. Dairy Sci. 2006, 89, 2873–2881. [Google Scholar] [CrossRef] [Green Version]
- Makino, S.; Sato, A.; Goto, A.; Nakamura, M.; Ogawa, M.; Chiba, Y.; Hemmi, J.; Kano, H.; Takeda, K.; Okumura, K.; et al. Enhanced natural killer cell activation by exopolysaccharides derived from yogurt fermented with Lactobacillus delbrueckii ssp. bulgaricus OLL1073R-1. J. Dairy Sci. 2016, 99, 915–923. [Google Scholar] [CrossRef] [Green Version]
- Nagai, T.; Makino, S.; Ikegami, S.; Itoh, H.; Yamada, H. Effects of oral administration of yogurt fermented with Lactobacillus delbrueckii ssp. bulgaricus OLL1073R-1 and its exopolysaccharides against influenza virus infection in mice. Int. Immunopharmacol. 2011, 11, 2246–2250. [Google Scholar] [CrossRef]
- Kanmani, P.; Albarracin, L.; Kobayashi, H.; Iida, H.; Komatsu, R.; Kober, A.H.; Ikeda-Ohtsubo, W.; Suda, Y.; Aso, H.; Makino, S.; et al. Exopolysaccharides from Lactobacillus delbrueckii OLL1073R-1 modulate innate antiviral immune response in porcine intestinal epithelial cells. Mol. Immunol. 2018, 93, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Dhanani, A.; Bagchi, T. The expression of adhesin EF-Tu in response to mucin and its role in Lactobacillus adhesion and competitive inhibition of enteropathogens to mucin. J. Appl. Microbiol. 2013, 115, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Harvey, K.L.; Jarocki, V.M.; Charles, I.G.; Djordjevic, S.P. The Diverse Functional Roles of Elongation Factor Tu (EF-Tu) in Microbial Pathogenesis. Front. Microbiol. 2019, 10, 2351. [Google Scholar] [CrossRef]
- Lin, J.-D.; Feng, N.; Sen, A.; Balan, M.; Tseng, H.-C.; McElrath, C.; Smirnov, S.V.; Peng, J.; Yasukawa, L.L.; Durbin, R.K.; et al. Distinct Roles of Type I and Type III Interferons in Intestinal Immunity to Homologous and Heterologous Rotavirus Infections. PLoS Pathog. 2016, 12, e1005600. [Google Scholar] [CrossRef]
- Meylan, E.; Tschopp, J. Toll-Like Receptors and RNA Helicases: Two Parallel Ways to Trigger Antiviral Responses. Mol. Cell 2006, 22, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Vijay-Kumar, M.; Gentsch, J.R.; Kaiser, W.J.; Borregaard, N.; Offermann, M.K.; Neish, A.S.; Gewirtz, A.T. Protein Kinase R Mediates Intestinal Epithelial Gene Remodeling in Response to Double-Stranded RNA and Live Rotavirus. J. Immunol. 2005, 174, 6322–6331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, H.-H.; Schneider, W.M.; Rice, C.M. Interferons and viruses: An evolutionary arms race of molecular interactions. Trends Immunol. 2015, 36, 124–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.E.; Oh, J.E.; Lee, H.K. Cell-Penetrating Mx1 Enhances Anti-Viral Resistance against Mucosal Influenza Viral Infection. Viruses 2019, 11, 109. [Google Scholar] [CrossRef]
- Sánchez-Tacuba, L.; Rojas, M.; Arias, C.F.; López, S. Rotavirus Controls Activation of the 2′-5′-Oligoadenylate Synthetase/RNase L Pathway Using at Least Two Distinct Mechanisms. J. Virol. 2015, 89, 12145–12153. [Google Scholar] [CrossRef] [Green Version]
- López, S.; Arias, C.F. Rotavirus–host cell interactions: An arms race. Curr. Opin. Virol. 2012, 2, 389–398. [Google Scholar] [CrossRef]
- Villena, J.; Salva, S.; Núñez, M.; Corzo, J.; Tolaba, R.; Faedda, J.; Font, G.; Alvarez, S. Probiotics for Everyone! The novel immunobiotic Lactobacillus rhamnosus CRL1505 and the beginning of social probiotic programs in Argentina. Int. J. Biotechnol. Wellness Ind. 2012, 1, 189–198. [Google Scholar]
- Kanmani, P.; Albarracin, L.; Kobayashi, H.; Hebert, E.M.; Saavedra, L.; Komatsu, R.; Gatica, B.; Miyazaki, A.; Ikeda-Ohtsubo, W.; Suda, Y.; et al. Genomic Characterization of Lactobacillus delbrueckii TUA4408L and Evaluation of the Antiviral Activities of its Extracellular Polysaccharides in Porcine Intestinal Epithelial Cells. Front. Immunol. 2018, 9, 2178. [Google Scholar] [CrossRef]
- Saitoh, T.; Yamamoto, M.; Miyagishi, M.; Taira, K.; Nakanishi, M.; Fujita, T.; Akira, S.; Yamamoto, N.; Yamaoka, S. A20 Is a Negative Regulator of IFN Regulatory Factor 3 Signaling. J. Immunol. 2005, 174, 1507–1512. [Google Scholar] [CrossRef] [Green Version]
- Hosoya, S.; Villena, J.; Shimazu, T.; Tohno, M.; Fujie, H.; Chiba, E.; Shimosato, T.; Aso, H.; Suda, Y.; Kawai, Y.; et al. Immunobiotic lactic acid bacteria beneficially regulate immune response triggered by poly(I:C) in porcine intestinal epithelial cells. Veter- Res. 2011, 42, 111. [Google Scholar] [CrossRef] [Green Version]
- Marranzino, G.; Villena, J.; Salva, S.; Alvarez, S. Stimulation of macrophages by immunobioticLactobacillusstrains: Influence beyond the intestinal tract. Microbiol. Immunol. 2012, 56, 771–781. [Google Scholar] [CrossRef]
- Miettinen, M.; Pietilä, T.E.; Kekkonen, R.A.; Kankainen, M.; Latvala, S.; Pirhonen, J.; Österlund, P.; Korpela, R.; Julkunen, I. Nonpathogenic Lactobacillus rhamnosus activates the inflammasome and antiviral responses in human macrophages. Gut Microbes 2012, 3, 510–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salva, S.; Villena, J.; Alvarez, S. Immunomodulatory activity of Lactobacillus rhamnosus strains isolated from goat milk: Impact on intestinal and respiratory infections. Int. J. Food Microbiol. 2010, 141, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Bain, C.; Mowat, A.M. Macrophages in intestinal homeostasis and inflammation. Immunol. Rev. 2014, 260, 102–117. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, X.; Li, J.; Zhou, Y.; Ho, W. RIG-I activation inhibits HIV replication in macrophages. J. Leukoc. Biol. 2013, 94, 337–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Guo, M.; Wang, X.; Li, J.; Wang, Y.; Ye, L.; Dai, M.; Zhou, L.; Persidsky, Y.; Ho, W. TLR3 activation efficiency by high or low molecular mass poly I:C. Innate Immun. 2013, 19, 184–192. [Google Scholar] [CrossRef] [Green Version]
- Mercer, J.; Greber, U.F. Virus interactions with endocytic pathways in macrophages and dendritic cells. Trends Microbiol. 2013, 21, 380–388. [Google Scholar] [CrossRef] [Green Version]
- Chen, I.-Y.; Ichinohe, T. Response of host inflammasomes to viral infection. Trends Microbiol. 2015, 23, 55–63. [Google Scholar] [CrossRef]
- Schroder, K.; Tschopp, J. The Inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.A.; Offit, P.A. Rotavirus-specific proteins are detected in murine macrophages in both intestinal and extraintestinal lymphoid tissues. Microb. Pathog. 1998, 24, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Graham, K.L.; O’Donnell, J.A.; Tan, Y.; Sanders, N.; Carrington, E.M.; Allison, J.; Coulson, B.S. Rotavirus Infection of Infant and Young Adult Nonobese Diabetic Mice Involves Extraintestinal Spread and Delays Diabetes Onset. J. Virol. 2007, 81, 6446–6458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Azevedo, M.S.; Wen, K.; Gonzalez, A.; Saif, L.J.; Li, G.; Yousef, A.E.; Yuan, L. Probiotic Lactobacillus acidophilus enhances the immunogenicity of an oral rotavirus vaccine in gnotobiotic pigs. Vaccine 2008, 26, 3655–3661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohanty, S.K.; Ivantes, C.A.P.; Mourya, R.; Pacheco, C.; A Bezerra, J. Macrophages Are Targeted by Rotavirus in Experimental Biliary Atresia and Induce Neutrophil Chemotaxis by Mip2/Cxcl2. Pediatr. Res. 2010, 67, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Ge, Y.; Mansell, A.; Ussher, J.; Brooks, A.; Manning, K.; Wang, C.J.H.; Taylor, J.A. Rotavirus NSP4 Triggers Secretion of Proinflammatory Cytokines from Macrophages via Toll-Like Receptor 2. J. Virol. 2013, 87, 11160–11167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guinn, Z.P.; Petro, T.M. Interferon regulatory factor 3 plays a role in macrophage responses to interferon-γ. Immunobiology 2019, 224, 565–574. [Google Scholar] [CrossRef]
- Österlund, P.I.; Pietilä, T.E.; Veckman, V.; Kotenko, S.V.; Julkunen, I. IFN Regulatory Factor Family Members Differentially Regulate the Expression of Type III IFN (IFN-lambda) Genes. J. Immunol. 2007, 179, 3434–3442. [Google Scholar] [CrossRef] [Green Version]
- Meyer, T.; A Horisberger, M. Combined action of mouse alpha and beta interferons in influenza virus-infected macrophages carrying the resistance gene Mx. J. Virol. 1984, 49, 709–716. [Google Scholar] [CrossRef] [Green Version]
- Behera, A.K.; Kumar, M.; Lockey, R.F.; Mohapatra, S.S. 2′-5′ Oligoadenylate Synthetase Plays a Critical Role in Interferon-gamma Inhibition of Respiratory Syncytial Virus Infection of Human Epithelial Cells. J. Biol. Chem. 2002, 277, 25601–25608. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, T. Protective effects of lactic acid bacteria on influenza A virus infection. AIMS Allergy Immunol. 2017, 1, 138–142. [Google Scholar] [CrossRef]
- Guo, L.; Xu, X.-Q.; Zhou, L.; Zhou, R.-H.; Wang, X.; Li, J.-L.; Liu, J.-B.; Liu, H.; Zhang, B.; Ho, W.-Z. Human Intestinal Epithelial Cells Release Antiviral Factors That Inhibit HIV Infection of Macrophages. Front. Immunol. 2018, 9, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tada, A.; Zelaya, H.; Clua, P.; Salva, S.; Alvarez, S.; Kitazawa, H.; Villena, J. Immunobiotic Lactobacillus strains reduce small intestinal injury induced by intraepithelial lymphocytes after Toll-like receptor 3 activation. Inflamm. Res. 2016, 65, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Villena, J.; Chiba, E.; Vizoso-Pinto, M.G.; Tomosada, Y.; Takahashi, T.; Ishizuka, T.; Aso, H.; Salva, S.; Álvarez, S.; Kitazawa, H. Immunobiotic Lactobacillus rhamnosus strains differentially modulate antiviral immune response in porcine intestinal epithelial and antigen presenting cells. BMC Microbiol. 2014, 14, 126. [Google Scholar] [CrossRef] [Green Version]
- Hakim, M.S.; Ding, S.; Chen, S.; Yin, Y.; Su, J.; van der Woude, C.J.; Fuhler, G.M.; Peppelenbosch, M.P.; Pan, Q.; Wang, W. TNF-α exerts potent anti-rotavirus effects via the activation of classical NF-κB pathway. Virus Res. 2018, 253, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Broggi, A.; Tan, Y.; Granucci, F.; Zanoni, I. IFN-λ suppresses intestinal inflammation by non-translational regulation of neutrophil function. Nat. Immunol. 2017, 18, 1084–1093. [Google Scholar] [CrossRef]
- Ware, C.F. Network Communications: Lymphotoxins, LIGHT, and TNF. Annu. Rev. Immunol. 2005, 23, 787–819. [Google Scholar] [CrossRef]
- Warfield, K.L.; Blutt, S.E.; Crawford, S.E.; Kang, G.; Conner, M.E. Rotavirus Infection Enhances Lipopolysaccharide-Induced Intussusception in a Mouse Model. J. Virol. 2006, 80, 12377–12386. [Google Scholar] [CrossRef] [Green Version]
- Turbay, M.E.; De Leblanc, A.d.M.; Perdigón, G.; De Giori, G.S.; Hebert, E.M. β-Casein hydrolysate generated by the cell envelope-associated proteinase of Lactobacillus delbrueckii ssp. lactis CRL 581 protects against trinitrobenzene sulfonic acid-induced colitis in mice. J. Dairy Sci. 2012, 95, 1108–1118. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elean, M.; Albarracin, L.; Fukuyama, K.; Zhou, B.; Tomokiyo, M.; Kitahara, S.; Araki, S.; Suda, Y.; Saavedra, L.; Villena, J.; et al. Lactobacillus delbrueckii CRL 581 Differentially Modulates TLR3-Triggered Antiviral Innate Immune Response in Intestinal Epithelial Cells and Macrophages. Microorganisms 2021, 9, 2449. https://doi.org/10.3390/microorganisms9122449
Elean M, Albarracin L, Fukuyama K, Zhou B, Tomokiyo M, Kitahara S, Araki S, Suda Y, Saavedra L, Villena J, et al. Lactobacillus delbrueckii CRL 581 Differentially Modulates TLR3-Triggered Antiviral Innate Immune Response in Intestinal Epithelial Cells and Macrophages. Microorganisms. 2021; 9(12):2449. https://doi.org/10.3390/microorganisms9122449
Chicago/Turabian StyleElean, Mariano, Leonardo Albarracin, Kohtaro Fukuyama, Binghui Zhou, Mikado Tomokiyo, Shugo Kitahara, Shota Araki, Yoshihito Suda, Lucila Saavedra, Julio Villena, and et al. 2021. "Lactobacillus delbrueckii CRL 581 Differentially Modulates TLR3-Triggered Antiviral Innate Immune Response in Intestinal Epithelial Cells and Macrophages" Microorganisms 9, no. 12: 2449. https://doi.org/10.3390/microorganisms9122449
APA StyleElean, M., Albarracin, L., Fukuyama, K., Zhou, B., Tomokiyo, M., Kitahara, S., Araki, S., Suda, Y., Saavedra, L., Villena, J., Hebert, E. M., & Kitazawa, H. (2021). Lactobacillus delbrueckii CRL 581 Differentially Modulates TLR3-Triggered Antiviral Innate Immune Response in Intestinal Epithelial Cells and Macrophages. Microorganisms, 9(12), 2449. https://doi.org/10.3390/microorganisms9122449