
Antibiotic Resistance and Pathogenomics of Staphylococci Circulating in Novosibirsk, Russia

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain Isolation and Identification
2.2. Susceptibility Testing
2.3. Antibiotic Resistance Genes Detection
2.4. Study of Biofilm Formation
2.5. Complete Genome Sequencing and Analysis
3. Results
3.1. Staphylococcus Strain Isolation and Identification
3.2. Antibiotic Resistance and Resistance-Encoding Genes
3.3. MDR Isolate Sequence Type Identification
3.4. Analysis of the Investigated Genomes for the Presence of Antibiotic Resistance Genes
3.5. Virulence Factor Identification
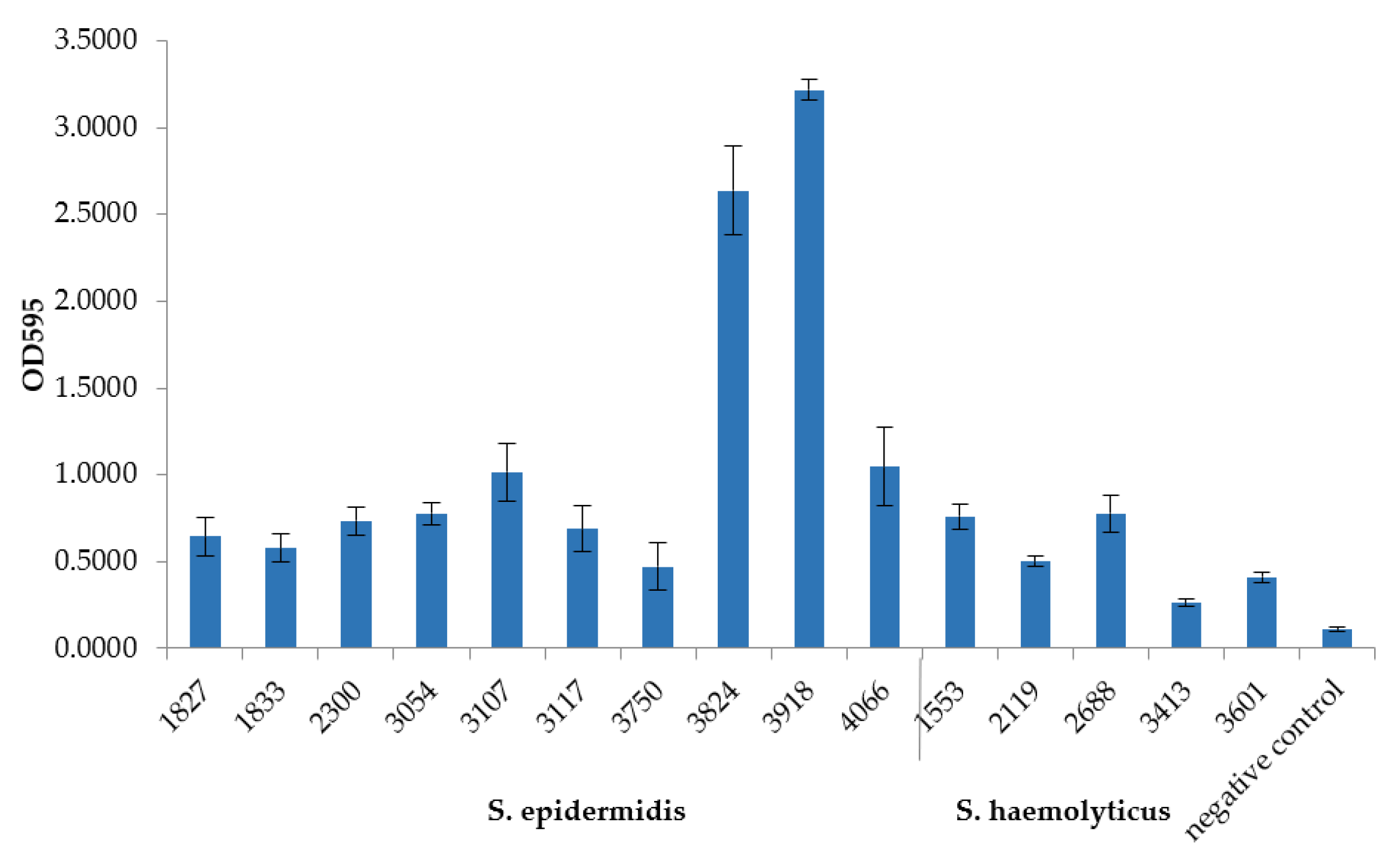
3.6. In Vitro Biofilm Formation by MDR S. epidermidis and S. haemolyticus Isolates and Factors Responsible for Adhesion and Biofilm Formation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Götz, F.; Bannerman, T.; Schleifer, K.H. The Prokaryotes; Springer: New York, NY, USA, 2006; pp. 5–75. [Google Scholar]
- Somayaji, R.; Priyantha, M.A.; Rubin, J.E.; Church, D. Human infections due to Staphylococcus pseudintermedius, an emerging zoonosis of canine origin: Report of 24 cases. Diagn. Microbiol. Infect. Dis. 2016, 85, 471–476. [Google Scholar] [CrossRef]
- Darlow, C.A.; Paidakakos, N.; Sikander, M.; Atkins, B. A spinal infection with Staphylococcus pseudintermedius. BMJ Case Rep. 2017, 2017, bcr2017221260. [Google Scholar] [CrossRef] [PubMed]
- Magleby, R.; Bemis, D.A.; Kim, D.; Carroll, K.C.; Castanheira, M.; Kania, S.A.; Jenkins, S.G.; Westblade, L.F. First reported human isolation of Staphylococcus delphini. Diagn. Microbiol. Infect. Dis. 2019, 94, 274–276. [Google Scholar] [CrossRef] [PubMed]
- Argemi, X.; Hansmann, Y.; Prola, K.; Prévost, G. Coagulase-Negative Staphylococci Pathogenomics. Int. J. Mol. Sci. 2019, 20, 1215. [Google Scholar] [CrossRef] [Green Version]
- Post, V.; Harris, L.G.; Morgenstern, M.; Mageiros, L.; Hitchings, M.D.; Méric, G.; Pascoe, B.; Sheppard, S.K.; Richards, R.G.; Moriarty, T.F. A comparative genomics study of Staphylococcus epidermidis from orthopedic device-related infections correlated with patient outcome. J. Clin. Microbiol. 2017, 55, 3089–3103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, M. Staphylococcus epidermidis—The ‘accidental’ pathogen. Nat. Rev. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef] [Green Version]
- Namvar, A.E.; Bastarahang, S.; Abbasi, N.; Ghehi, G.S.; Farhadbakhtiarian, S.; Arezi, P.; Hosseini, M.; Baravati, S.Z.; Jokar, Z.; Chermahin, S.G. Clinical characteristics of Staphylococcus epidermidis: A systematic review. GMS Hyg. Infect. Control 2014, 9, Doc23. [Google Scholar] [CrossRef]
- Gómez, P.; Lozano, C.; Benito, D.; Estepa, V.; Tenorio, C.; Zarazaga, M.; Torres, C. Characterization of staphylococci in urban wastewater treatment plants in Spain, with detection of methicillin resistant Staphylococcus aureus ST398. Environ. Pollut. 2016, 212, 71–76. [Google Scholar] [CrossRef]
- Xu, Z.; Misra, R.; Jamrozy, D.; Paterson, G.K.; Cutler, R.R.; Holmes, M.A.; Gharbia, S.; Mkrtchyan, H.V. Whole genome sequence and comparative genomics analysis of multi-drug resistant environmental Staphylococcus epidermidis ST59. G3 Genes Genomes Genet. 2018, 8, 2225–2230. [Google Scholar] [CrossRef] [Green Version]
- Stratchounski, L.S.; Dekhnich, A.V.; Kretchikov, V.A.; Edelstain, I.A.; Narezkina, A.D.; Afinogenov, G.E.; Akhmetova, L.I.; Boronina, L.G.; Gugutcidze, E.N.; Gudkova, L.V.; et al. Antimicrobial resistance of nosocomial strains of Staphylococcus aureus in Russia: Results of a prospective study. J. Chemother. 2005, 17, 54–60. [Google Scholar] [CrossRef]
- Dekhnich, A.V.; Nikulin, A.A.; Ryabkova, E.L.; Krechikova, O.I.; Sukhorukova, M.V.; Kozlov, R.S.; ROSNET Study Group. Epidemiology of resistance of S. aureus strains isolated from patients in the ICU of Russian hospitals: Results of a multicenter study. Clin. Microbiol. Antimicrob. Chemother. 2008, 10, 333–344. (In Russian) [Google Scholar]
- Sukhorukova, M.V.; Skleenova, E.Y.; Ivanchik, N.V.; Timokhova, A.V.; Edelstein, M.V.; Dekhnich, A.V.; Kozlov, R.S.; Shek, E.A.; «MARATHON» Study Group. Antimicrobial resistance of nosocomial Staphylococcus aureus isolates in Russia: Results of national multicenter surveillance study «MARATHON» 2011–2012. Clin. Microbiol. Antimicrob. Chemother. 2014, 16, 280–286. (In Russian) [Google Scholar]
- Romanov, A.V.; Dekhnich, A.V.; Sukhorukova, M.V.; Skleenova, E.Y.; Ivanchik, N.V.; Edelstein, M.V.; Kozlov, R.S.; the «MARATHON» Study Group. Antimicrobial resistance of nosocomial Staphylococcus aureus isolates in Russia: Results of multicenter epidemiological study «MARATHON» 2013–2014. Clin. Microbiol. Antimicrob. Chemother. 2017, 19, 57–62. (In Russian) [Google Scholar]
- Barantsevich, N.E.; Barantsevich, E.P. Species diversity and methicillin resistance in Staphylococcus spp. in nosocomial infections. Clin. Microbiol. Antimicrob. Chemother. 2019, 21, 207–211. (In Russian) [Google Scholar] [CrossRef] [Green Version]
- Kozlova, Y.N.; Fomenko, N.V.; Morozova, V.V.; Saranina, I.; Tikunov, A.; Ganichev, D.A.; Samokhin, A.; Pavlov, V.; Rozhnova, O.; Bondar, I.A.; et al. Genetic and biochemical characterization of staphylococci occurring in Novosibirsk, Russia. Vavilovskii Zhurnal Genet. Sel. 2017, 21, 952–958. [Google Scholar] [CrossRef] [Green Version]
- Fursova, K.; Sorokin, A.; Sokolov, S.; Dzhelyadin, T.; Shulcheva, I.; Shchannikova, M.; Nikanova, D.; Artem’eva, O.; Zinovieva, N.; Brovko, F. Virulence factors and phylogeny of Staphylococcus aureus associated with bovine mastitis in Russia based on genome sequences. Front. Vet. Sci. 2020, 7, 135. [Google Scholar] [CrossRef]
- Khokhlova, O.E.; Hung, W.C.; Wan, T.W.; Iwao, Y.; Takano, T.; Higuchi, W.; Yachenko, S.V.; Teplyakova, O.V.; Kamshilova, V.V.; Kotlovsky, Y.V.; et al. Healthcare- and community-associated methicillin-resistant Staphylococcus aureus (MRSA) and fatal pneumonia with pediatric deaths in Krasnoyarsk, Siberian Russia: Unique MRSA’s multiple virulence factors, genome, and stepwise evolution. PLoS ONE 2015, 10, e0128017. [Google Scholar] [CrossRef]
- Wan, T.W.; Khokhlova, O.E.; Iwao, Y.; Higuchi, W.; Hung, W.C.; Reva, I.V.; Singur, O.A.; Gostev, V.V.; Sidorenko, S.V.; Peryanova, O.V.; et al. Complete circular genome sequence of successful ST8/SCCmecIV community-associated methicillin-resistant Staphylococcus aureus (OC8) in Russia: One-megabase genomic inversion, IS256’s spread, and evolution of Russia ST8-IV. PLoS ONE 2016, 11, e0164168. [Google Scholar] [CrossRef]
- Morozova, V.; Kozlova, Y.; Shedko, E.; Kurilshikov, A.; Babkin, I.; Tupikin, A.; Yunusova, A.; Chernonosov, A.; Baykov, I.; Kondratov, I. Lytic Bacteriophage PM16 Specific for Proteus mirabilis: A Novel Member of the genus phiKMVvirus. Arch. Virol. 2016, 161, 2457–2472. [Google Scholar] [CrossRef]
- Madhaiyan, M.; Wirth, J.S.; Saravanan, V.S. Phylogenomic analyses of the Staphylococcaceae family suggest the reclassification of five species within the genus Staphylococcus as heterotypic synonyms, the promotion of five subspecies to novel species, the taxonomic reassignment of five Staphylococcus species to Mammaliicoccus gen. nov., and the formal assignment of Nosocomiicoccus to the family Staphylococcaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 5926–5936. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Hola, V.; Di Bonaventura, G.; Djukić, S.; Cirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef] [PubMed]
- Kocianova, S.; Vuong, C.; Yao, Y.; Voyich, J.M.; Fischer, E.R.; DeLeo, F.R.; Otto, M. Key role of poly-gamma-DL-glutamic acid in immune evasion and virulence of Staphylococcus epidermidis. J. Clin. Investig. 2005, 115, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, F.; Watanabe, S.; Baba, T.; Yuzawa, H.; Ito, T.; Morimoto, Y.; Kuroda, M.; Cui, L.; Takahashi, M.; Ankai, A.; et al. Whole-genome sequencing of Staphylococcus haemolyticus uncovers the extreme plasticity of its genome and the evolution of human-colonizing staphylococcal species. J. Bacteriol. 2005, 187, 7292–7308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flahaut, S.; Vinogradov, E.; Kelley, K.A.; Brennan, S.; Hiramatsu, K.; Lee, J.C. Structural and biological characterization of a capsular polysaccharide produced by Staphylococcus haemolyticus. J. Bacteriol. 2008, 190, 1649–1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keinhörster, D.; Salzer, A.; Duque-Jaramillo, A.; George, S.E.; Marincola, G.; Lee, J.C.; Weidenmaier, C.; Wolz, C. Revisiting the regulation of the capsular polysaccharide biosynthesis gene cluster in Staphylococcus aureus. Mol. Microbiol. 2019, 112, 1083–1099. [Google Scholar] [CrossRef] [Green Version]
- Arciola, C.R.; Campoccia, D.; Ravaioli, S.; Montanaro, L. Polysaccharide intercellular adhesin in biofilm: Structural and regulatory aspects. Front. Cell. Infect. Microbiol. 2015, 5, 7. [Google Scholar] [CrossRef] [Green Version]
- Foster, T.J. Surface Proteins of Staphylococcus epidermidis. Front. Microbiol. 2020, 11, 1829. [Google Scholar] [CrossRef]
- Clarke, S.R.; Harris, L.G.; Richards, R.G.; Foster, S.J. Analysis of Ebh, a 1.1-megadalton cell wall-associated fibronectin-binding protein of Staphylococcus aureus. Infect. Immun. 2002, 70, 6680–6687. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Khan, B.A.; Cheung, G.Y.; Bach, T.H.; Jameson-Lee, M.; Kong, K.F.; Queck, S.Y.; Otto, M. Staphylococcus epidermidis surfactant peptides promote biofilm maturation and dissemination of biofilm-associated infection in mice. J. Clin. Investig. 2011, 121, 238–248. [Google Scholar] [CrossRef]
- Dastgheyb, S.S.; Villaruz, A.E.; Le, K.Y.; Tan, V.Y.; Duong, A.C.; Chatterjee, S.S.; Cheung, G.Y.C.; Joo, H.-S.; Hickok, N.J.; Otto, M. Role of phenol-soluble modulins in formation of Staphylococcus aureus biofilms in synovial fluid. Infect. Immun. 2015, 83, 2966–2975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, K.Y.; Park, M.D.; Otto, M. Immune evasion mechanisms of Staphylococcus epidermidis biofilm infection. Front. Microbiol. 2018, 9, 359. [Google Scholar] [CrossRef] [PubMed]
- Pöhlmann-Dietze, P.; Ulrich, M.; Kiser, K.B.; Döring, G.; Lee, J.C.; Fournier, J.M.; Botzenhart, K.; Wolz, C. Adherence of Staphylococcus aureus to endothelial cells: Influence of capsular polysaccharide, global regulator agr, and bacterial growth phase. Infect. Immun. 2000, 68, 4865–4871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santajit, S.; Indrawattana, N. Mechanisms of antimicrobial resistance in ESKAPE pathogens. BioMed Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suwantarat, N.; Carroll, K.C. Epidemiology and molecular characterization of multidrug-resistant Gram-negative bacteria in Southeast Asia. Antimicrob. Resist. Infect. Control 2016, 5, 15. [Google Scholar] [CrossRef] [Green Version]
- Pedroso, S.; Sandes, S.; Filho, R.; Nunes, A.C.; Serufo, J.C.; Farias, L.M.; Carvalho, M.; Bomfim, M.; Santos, S.G. Coagulase-negative staphylococci isolated from human bloodstream infections showed multidrug resistance profile. Microb. Drug Resist. 2018, 24, 635–647. [Google Scholar] [CrossRef]
- Asante, J.; Amoako, D.G.; Abia, A.; Somboro, A.M.; Govinden, U.; Bester, L.A.; Essack, S.Y. Review of clinically and epidemiologically relevant coagulase-negative staphylococci in Africa. Microbial. Drug Resist. 2020, 26, 951–970. [Google Scholar] [CrossRef] [Green Version]
- Ben Zakour, N.L.; Beatson, S.A.; van den Broek, A.H.; Thoday, K.L.; Fitzgerald, J.R. Comparative genomics of the Staphylococcus intermedius group of animal pathogens. Front. Cell. Infect. Microbiol. 2012, 2, 44. [Google Scholar] [CrossRef] [Green Version]
- Jantorn, P.; Heemmamad, H.; Soimala, T.; Indoung, S.; Saising, J.; Chokpaisarn, J.; Wanna, W.; Tipmanee, V.; Saeloh, D. Antibiotic Resistance Profile and Biofilm Production of Staphylococcus pseudintermedius Isolated from Dogs in Thailand. Pharmaceuticals 2021, 14, 592. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Zhu, Y.; Song, Y.; Li, T.; Luo, T.; Sun, G.; Yang, C.; Cao, C.; Lu, Y.; Li, M. Molecular analysis of Staphylococcus epidermidis strains isolated from community and hospital environments in China. PLoS ONE 2013, 8, e62742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Cave, R.; Chen, L.; Yangkyi, T.; Liu, Y.; Li, K.; Meng, G.; Niu, K.; Zhang, W.; Tang, N.; et al. Antibiotic resistance and molecular characteristics of methicillin-resistant Staphylococcus epidermidis recovered from hospital personnel in China. J. Glob. Antimicrob. Resist. 2020, 22, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Seng, R.; Leungtongkam, U.; Thummeepak, R.; Chatdumrong, W.; Sitthisak, S. High prevalence of methicillin-resistant coagulase-negative staphylococci isolated from a university environment in Thailand. Int. Microbiol. 2017, 20, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Miragaia, M.; Thomas, J.C.; Couto, I.; Enright, M.C.; de Lencastre, H. Inferring a population structure for Staphylococcus epidermidis from multilocus sequence typing data. J. Bacteriol. 2007, 189, 2540–2552. [Google Scholar] [CrossRef] [Green Version]
- Mendes, R.E.; Deshpande, L.M.; Costello, A.J.; Farrell, D.J. Molecular epidemiology of Staphylococcus epidermidis clinical isolates from U.S. hospitals. Antimicrob. Agents Chemother. 2012, 56, 4656–4661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deplano, A.; Vandendriessche, S.; Nonhoff, C.; Dodemont, M.; Roisin, S.; Denis, O. National surveillance of Staphylococcus epidermidis recovered from bloodstream infections in Belgian hospitals. J. Antimicrob. Chemother. 2016, 71, 1815–1819. [Google Scholar] [CrossRef] [Green Version]
- Cremniter, J.; Sivadon-Tardy, V.; Caulliez, C.; Bauer, T.; Porcher, R.; Lortat-Jacob, A.; Piriou, P.; Judet, T.; Aegerter, P.; Herrmann, J.-L.; et al. Genetic analysis of glycopeptide-resistant Staphylococcus epidermidis strains from bone and joint infections. J. Clin. Microbiol. 2013, 51, 1014–1019. [Google Scholar] [CrossRef] [Green Version]
- Both, A.; Huang, J.; Qi, M.; Lausmann, C.; Weißelberg, S.; Büttner, H.; Lezius, S.; AFailla, V.; Christner, M.; Stegger, M.; et al. Distinct clonal lineages and within-host diversification shape invasive Staphylococcus epidermidis populations. PLoS Pathog. 2021, 17, e1009304. [Google Scholar] [CrossRef]
- Skachkova, T.S.; Zamyatin, M.N.; Orlova, O.A.; Yumtsunova, N.A.; Lashenkova, N.N.; Fomina, V.S.; Gusarov, V.G.; Mikhaylova, Y.V.; Shelenkov, A.A.; Goloveshkina, E.N.; et al. Monitoring methicillin-resistant staphylococcus strains in the Moscow medical and surgical center using molecular-biological methods. Epidemiol. Vaccinal Prev. 2021, 20, 44–50. [Google Scholar] [CrossRef]
- Lee, J.Y.H.; Monk, I.R.; Gonçalves da Silva, A.; Seemann, T.; Chua, K.Y.L.; Kearns, A.; Hill, R.; Woodford, N.; Bartels, M.D.; Strommenger, B.; et al. Global spread of three multidrug-resistant lineages of Staphylococcus epidermidis. Nat. Microbiol. 2018, 3, 1175–1185. [Google Scholar] [CrossRef]
- Arora, S.; Li, X.; Hillhouse, A.; Konganti, K.; Little, S.V.; Lawhon, S.D.; Threadgill, D.; Shelburne, S.; Hook, M. Staphylococcus epidermidis MSCRAMM SesJ is encoded in composite islands. mBio 2020, 11, e02911-19. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Panels | Origin of Strains | Number of Isolates |
|---|---|---|
| Strains isolated from hospitalized patients (n = 105) | Prosthetic joint and post-operative wound infections | 56 |
| Purulent diabetic ulcers | 43 | |
| Decubitus ulcers | 3 | |
| Sputum from a patient with pneumonia | 2 | |
| Cerebrospinal fluid | 1 | |
| Strains from outpatients (n = 228) | Respiratory tract infections | 83 |
| Skin and mucosal infections | 59 | |
| Urinary tract infections | 21 | |
| Bacterial vaginosis | 25 | |
| Fecal samples from patients with diarrhea | 36 | |
| Purulent diabetic ulcers | 4 | |
| Strains from pets (39 from dogs and 22 from cats) (n = 61) | Skin and mucosal infections | 57 |
| Fecal samples from animals with diarrhea | 4 |
| No | Species (Number of Isolates) | Isolation Source (Number of Isolates) | GenBank Identifier for 16S rRNA Gene |
|---|---|---|---|
| 1 | Staphylococcus aureus (167) | Hospital (74) | No ID |
| Outpatient (81) | No ID | ||
| Veterinary (12) | No ID | ||
| 2 | Staphylococcus auricularis (4) | Outpatient (4) | MZ014399–MZ014402 |
| 3 | Staphylococcus borealis (2) | Outpatient (2) | MZ014403, MZ014404 |
| 4 | Staphylococcus capitis (4) | Outpatient (4) | MZ014405–MZ014407, MZ014411 |
| 5 | Staphylococcus caprae (5) | Hospital (1) | MZ014409 |
| Outpatient (4) | MZ014408, MZ014410, MZ014412, MZ014413 | ||
| 6 | Staphylococcus carnosus (2) | Outpatient (2) | MZ014414, MZ014415 |
| 7 | Staphylococcus casei (2) | Outpatient (1) | MZ014416 |
| 8 | Staphylococcus coagulans/S. schleiferi subsp. coagulans (8) | Veterinary (8) | MW979964–MW979970 |
| 9 | Staphylococcus cohnii (2) | Outpatient (2) | MZ014417, MZ014418 |
| 10 | Staphylococcus devriesei (4) | Outpatient (4) | MZ014419–MZ014422 |
| 11 | Staphylococcus equorum (3) | Outpatient (1) | MZ014423 |
| Veterinary (2) | MW979955, MW979971 | ||
| 12 | Staphylococcus epidermidis (104) | Hospital (20) | MZ027385–MZ027397, MZ040881, MZ040882, MZ041685 |
| Outpatient (84) | MZ027349–MZ027358, MZ027360–MZ027362, MZ027364–MZ027366, MZ040893–MZ040916, MZ047203–MZ047210, MZ049531–MZ049537,MZ723062 | ||
| 13 | Staphylococcus felis (5) | Veterinary (5) | MW979972–MW979976 |
| 14 | Staphylococcus haemolyticus (22) | Hospital (2) | MZ027398, MZ027399 |
| Outpatient (19) | MZ027367–MZ027375, MZ040917–MZ040921, MZ047213–MZ047216, MZ723059, MZ723059 | ||
| Veterinary (1) | MW979956 | ||
| 15 | Staphylococcus hominis (14) | Hospital (2) | MZ014434, MZ014435 |
| Outpatient (11) | MZ014424–MZ014433 | ||
| Veterinary (1) | MW979977 | ||
| 16 | Staphylococcus pasteuri (1) | Outpatient (1) | MZ047217 |
| 17 | Staphylococcus pseudintermedius (24) | Veterinary (24) | MW965646, MW979953, MW979978–MW979995, MW979997, MW979998 |
| 18 | Staphylococcus simulans (8) | Hospital (1) | MZ014437 |
| Outpatient (5) | MZ014436, MZ014438–MZ014441 | ||
| Veterinary (2) | MW979999, MW980000 | ||
| 19 | Staphylococcus warneri (8) | Hospital (2) | MZ014445, MZ014447 |
| Outpatient (5) | MZ014442–MZ014444, MZ014446, MZ014448 | ||
| Veterinary (1) | MW979957 | ||
| 20 | Mammaliicoccus sciuri/Staphylococcus sciuri (3) 1 | Hospital (1) | MW965541 |
| Veterinary (2) | MW965545, MW965551 | ||
| 21 | Mammaliicoccus vitulinus/Staphylococcus vitulinus (1) 1 | Veterinary (1) | MW965552 |
| 22 | Mammaliicoccus lentus/Staphylococcus lentus (1) 1 | Veterinary (1) | MW965543 |
| Antimicrobial | Resistance-Encoding Gene(s) 1 | Number of Isolates with Resistance Profiles and ARGs That Were Identified in These Isolates | ||
|---|---|---|---|---|
| S. aureus | ||||
| Hospital (n = 74) | Outpatient (n = 81) | Veterinary (n = 12) | ||
| Cefoxitin | 13 | 2 | 0 | |
| mecA | 13 | 2 | 0 | |
| Penicillin | N.d 2 | N.d. | N.d. | |
| blaZ | 32 | 38 | 3 | |
| Gentamicin and/or Amikacin | 12 | 2 | 0 | |
| aac(6′)-Ie-aph(2″)-Ia | 8 | 1 | 0 | |
| aac(6′)-Ie-aph(2″)-Ia + ant(4′)-Ia | 1 | 1 | 0 | |
| aac(6′)-Ie-aph(2″)-Ia + aph(3′)-IIIa | 3 | 0 | 0 | |
| Erythromycin | 0 | 5 | 0 | |
| ermC | 0 | 5 | 0 | |
| Erythromycin and Clindamycin | 7 | 5 | 0 | |
| ermA | 5 | 0 | 0 | |
| ermC | 2 | 5 | 0 | |
| S. epidermidis | ||||
| Hospital (n = 20) | Outpatient (n = 84) | Veterinary (n = 0) | ||
| Cefoxitin | 13 | 18 | 0 | |
| mecA | 13 | 18 | 0 | |
| Penicillin | N.d. | N.d. | N.d. | |
| blaZ | 6 | 39 | 0 | |
| Gentamicin and/or Amikacin | 9 | 19 | 0 | |
| aac(6′)-Ie-aph(2″)-Ia | 4 | 11 | 0 | |
| ant(4′)-Ia | 0 | 3 | 0 | |
| aac(6′)-Ie-aph(2″)-Ia + ant(4′)-Ia | 5 | 2 | 0 | |
| aac(6′)-Ie-aph(2″)-Ia + aph(3′)-IIIa | 0 | 3 | 0 | |
| Erythromycin | 6 | 34 | 0 | |
| ermC | 2 | 5 | 0 | |
| msrA | 4 | 29 | 0 | |
| Clindamycin | 1 3 | 1 3 | 0 | |
| Erythromycin and Clindamycin | 4 | 2 | 0 | |
| ermA | 3 | 0 | 0 | |
| ermC | 1 | 0 | 0 | |
| msrA | 0 | 2 | 0 | |
| S. haemolyticus | ||||
| Hospital (n = 2) | Outpatient (n = 19) | Veterinary (n = 1) | ||
| Cefoxitin | 2 | 10 | 1 | |
| mecA | 2 | 10 | 1 | |
| Penicillin | N.d. | N.d. | N.d. | |
| blaZ | 1 | 3 | 0 | |
| Gentamicin and/or Amikacin | 2 | 10 | 1 | |
| aac(6′)-Ie-aph(2″)-Ia | 1 | 7 | 1 | |
| ant(4′)-Ia | 0 | 4 4 | 0 | |
| aac(6′)-Ie-aph(2″)-Ia + aph(3′)-IIIa | 1 | 3 | 0 | |
| Erythromycin | 2 | 13 | 0 | |
| ermC | 0 | 3 | 0 | |
| msrA | 2 | 10 | 1 4 | |
| S. hominis | ||||
| Hospital (n = 2) | Outpatient (n = 19) | Veterinary (n = 1) | ||
| Cefoxitin | 0 | 2 | 0 | |
| mecA | 0 | 2 | 0 | |
| Penicillin | N.d. | N.d. | N.d. | |
| blaZ | 1 | 3 | 0 | |
| Gentamicin and/or Amikacin | 0 | 2 | 0 | |
| ant(4′)-Ia | 0 | 1 4 | 0 | |
| aph(3′)-IIIa | 0 | 0 | 1 4 | |
| aac(6′)-Ie-aph(2″)-Ia + ant(4′)-Ia | 0 | 2 | 0 | |
| Erythromycin | 0 | 5 | 0 | |
| msrA | 0 | 4 | 0 | |
| ermA + msrA | 0 | 1 | 0 | |
| S. pseudintermedius | ||||
| Hospital (n = 0) | Outpatient (n = 0) | Veterinary (n = 24) | ||
| Penicillin | N.d. | N.d. | N.d. | |
| blaZ | 0 | 0 | 4 | |
| Gentamicin and/or Amikacin | 0 | 0 | 6 | |
| aac(6′)-Ie-aph(2″)-Ia | 0 | 0 | 1 | |
| aph(3′)-IIIa | 0 | 0 | 1 | |
| aac(6′)-Ie-aph(2″)-Ia + aph(3′)-IIIa | 0 | 0 | 4 | |
| Erythromycin and Clindamycin | 0 | 0 | 7 3 | |
| No | Species | CEMTC No of Isolate/16S rRNA GenBank ID | Source of Isolation | Resistance | Sequence Type | Clonal Complex |
|---|---|---|---|---|---|---|
| 1 | S. epidermidis | 1827/MZ027386 | Hospital (swab from purulent diabetic ulcer) | FOX, AK, CN, E, DA | ST 23 | CC23 |
| 2 | S. epidermidis | 1833/MZ027390 | Hospital (pure culture, post-operative wound infection) | FOX, AK, CN, E, DA | ST 23 | CC23 |
| 3 | S. epidermidis | 2300/MZ027350 | Outpatient (nasal swab, rhinitis) | FOX, AK, CN | ST 20 | S 1 |
| 4 | S. epidermidis | 3054/MZ027389 | Hospital (pure culture, post-operative wound infection) | FOX, CN, E, DA | ST 5 | CC5 |
| 5 | S. epidermidis | 3107/MZ027352 | Outpatient (nasal swab, rhinitis) | FOX, CN, E | ST 152 | CC5 |
| 6 | S. epidermidis | 3117/MZ027364 | Outpatient (faeces, diarrhea) | FOX, CN, E | ST 152 | CC5 |
| 7 | S. epidermidis | 3750/MZ027394 | Hospital (biopsy material, prosthetic joint infection) | FOX, AK, CN | ST 210 | CC5 |
| 8 | S. epidermidis | 3824/MZ027395 | Hospital (biopsy material, prosthetic joint infection) | FOX, AK, CN, E | ST 786 | CC5 |
| 9 | S. epidermidis | 3918/MZ027396 | Hospital (pure culture, post-operative wound infection) | FOX, AK, CN, E | ST 20 | S 1 |
| 10 | S. epidermidis | 4066/MZ027397 | Hospital (pure culture, post-operative wound infection) | FOX, AK, CN, E, DA | ST 17 | CC5 |
| 11 | S. haemolyticus | 1553/MZ027371 | Outpatient (urine sample, pyelonephritis) | FOX, AK, CN, E | ST 1 | CC3 |
| 12 | S. haemolyticus | 2119/MZ027368 | Outpatient (purulent diabetic ulcer) | FOX, CN, E | ST 3 | CC3 |
| 13 | S. haemolyticus | 2688/MZ723059 | Outpatient (faeces, diarrhea) | FOX, CN, E | ST 3 | CC3 |
| 14 | S. haemolyticus | 3413/MZ027399 | Hospital (sputum, ventilator-associated pneumonia) | FOX, AK, CN, E | ST 42 | CC3 |
| 15 | S. haemolyticus | 3601/MZ027370 | Outpatient (skin infection, skin scraping) | FOX, AK, CN, E | ST 8 | CC3 |
| Antibiotic Classes | ARG | S. epidermidis Isolates 1 | S. haemolyticus Isolates 1 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1827 | 1833 | 2300 | 3054 | 3107 | 3117 | 3750 | 3824 | 3918 | 4066 | 1553 | 2119 | 2688 | 3413 | 3601 | ||
| Penicillins | blaZ | ✓ 2 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||||
| Cephalosporins | mecA | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ |
| mecI | ✓ | |||||||||||||||
| mecR1 | ✓ | ✓ | ✓ | ✓ | ✓ | |||||||||||
| Macrolides and Lincosamides | ermA | ✓ | ✓ | ✓ | ||||||||||||
| ermC | ✓ | ✓ | ✓ | ✓ | ✓ | |||||||||||
| msrA | ✓ | ✓ | ✓ | ✓ | ✓ | |||||||||||
| mphC | ✓ | ✓ | ✓ | ✓ | ✓ | |||||||||||
| Aminoglycosides | aph(3′)-IIIa | ✓ | ✓ | ✓ | ||||||||||||
| ant(4′)-Ia | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||||||
| aac(6’)-Ie-aph(2″)-Ia | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |
| aad(6) | ✓ | ✓ | ||||||||||||||
| ant(9) | ✓ | ✓ | ✓ | |||||||||||||
| Quinolones | norA | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |||||
| qacA | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |||||||
| qacB | ✓ | ✓ | ✓ | |||||||||||||
| Chlorampheni- col | catA8 | ✓ | ||||||||||||||
| catA7 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||||||
| Fusidic acid | fusC | ✓ | ||||||||||||||
| Diaminopyrimi- dines | dfrC | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |||||
| dfrG | ✓ | ✓ | ✓ | ✓ | ✓ | |||||||||||
| Aminocoumarins | Aminocoumarin resistant gyrB | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||
| Lincosamides | lnuA | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||||
| Tetracyclines | tet(K) | ✓ | ✓ | ✓ | ||||||||||||
| tet(45) | ✓ | |||||||||||||||
| Fosfomycin | fosB | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |||||
| Bacitracin | BceSR | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ |
| Species and No in CEMTC | Exoenzymes | Toxins | Immune Evasion | |||||
|---|---|---|---|---|---|---|---|---|
| Cysteine Protease | Serine V8 Protease | Lipases | Thermonuclease | Hemo- lysin III | Hemolysin, Containing CBS Domain | β-Hemolysin | PGA, Capsule | |
| S. epidermidis 1827 | sspB | sspA | geh, lip | nuc | + 1 | + 1 | hlb | capA, pgsABCDE 2 |
| S. epidermidis 1833 | sspB | sspA | geh, lip | nuc | + | + | hlb | capA, pgsABCDE |
| S. epidermidis 2300 | sspB | sspA | geh, lip | nuc | + | + | hlb | capA, pgsABCDE |
| S. epidermidis 3054 | sspB | sspA | geh, lip | nuc | + | + | hlb | capA, pgsABCDE |
| S. epidermidis 3107 | sspB | sspA | geh, lip | nuc | + | + | hlb | capA, pgsABCDE |
| S. epidermidis 3117 | sspB | sspA | geh, lip | nuc | + | + | hlb | capA, pgsABCDE |
| S. epidermidis 3750 | sspB | sspA | geh, lip | nuc | + | + | hlb | capA, pgsABCDE |
| S. epidermidis 3824 | sspB | sspA | geh, lip | nuc | + | + | hlb | capA, pgsABCDE |
| S. epidermidis 3918 | sspB | sspA | geh, lip | nuc | + | + | hlb | capA, pgsABCDE |
| S. epidermidis 4066 | sspB | sspA | geh, lip | nuc | + | + | hlb | capA, pgsABCDE |
| S. haemolyticus 1553 | lip | nuc | + | + | capA, pgsABCDE | |||
| S. haemolyticus 2119 | sspB | sspA | geh, lip | nuc | + | + | capA, pgsABCDE, cap5L, cap5F, cap8C, cap8E, cap8I, cap8H, cap 8M, cap8N2 | |
| S. haemolyticus 2688 | lip | nuc | + | + | capA, pgsABCDE, cap5A | |||
| S. haemolyticus 3413 | lip | nuc | + | + | capA, pgsABCDE, cap5L, cap5F, cap8C, cap8E, cap8I, cap8H, cap 8M, cap8N2 | |||
| S. haemolyticus 3601 | lip | nuc | + | + | capA, pgsABCDE, cap5A, cap8C3 | |||
| Species and No in CEMTC | Ability to Form Biofilm | Adherence and Biofilm Formation Genes | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| PIA | PSM | Aap | Pls | Sas-Family Proteins | Autoly- sin | Fibronectin Binding Protein 1 | Elastin Binding Protein | Sdr-Family Proteins | ||
| S. epidermidis 1827 | strong | icaADBC, icaR | psmβ1, psmβ2 | pls | SasA, SasC, SasF | atlE | ebh | ebp | sdrG, sdrH | |
| S. epidermidis 1833 | moderate | icaADBC, icaR | pls | SasA, SasC, SasF | atlE | ebh | ebp | sdrG | ||
| S. epidermidis 2300 | strong | icaADBC, icaR | psmβ1, psmβ2 | aap | pls | SasA, SasC, SasF | atlE | ebh | ebp | sdrC, sdrF, sdrH |
| S. epidermidis 3054 | strong | psmβ1, psmβ2 | aap | SasA, SasC, SasF | atlE | ebh | ebp | sdrF, sdrG, sdrH | ||
| S. epidermidis 3107 | strong | psmβ1, psmβ2 | aap | SasA, SasC, SasF | atlE | ebh | ebp | sdrF, sdrG, sdrH | ||
| S. epidermidis 3117 | strong | psmβ1, psmβ2 | aap | SasA, SasC, SasF | atlE | ebh | ebp | sdrF, sdrG, sdrH | ||
| S. epidermidis 3750 | moderate | psmβ1, psmβ2 | pls | SasA, SasC, SasF | atlE | ebh | ebp | sdrE, sdrF, sdrG, sdrH | ||
| S. epidermidis 3824 | strong | icaADBC, icaR | aap | pls | SasA, SasC, SasF | atlE | ebp | sdrE, sdrF, sdrG, sdrH | ||
| S. epidermidis 3918 | strong | icaADBC, icaR | aap | pls | SasA, SasC, SasF, SasG | atlE | ebh | ebp | sdrE, sdrF, sdrG, sdrH | |
| S. epidermidis 4066 | strong | icaADBC, icaR | psmβ1, psmβ2 | aap | SasA, SasC, SasF | atlE | ebp | sdrF, sdrG, sdrH | ||
| S. haemolyticus 1553 | strong | aap | SasA, SasC, SasF, SasG | atlE | ebp | |||||
| S. haemolyticus 2119 | moderate | SasA, SasC, SasF, SasG | atlE | ebp | ||||||
| S. haemolyticus 2688 | strong | SasA, SasC, SasF, SasG | atlE | ebp | ||||||
| S. haemolyticus 3413 | weak | aap | SasA, SasC, SasF, SasG | atlE | ebp | sdrC | ||||
| S. haemolyticus 3601 | moderate | SasA, SasC, SasF, SasG | atlE | ebp | sdrC | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bardasheva, A.; Tikunov, A.; Kozlova, Y.; Zhirakovskaia, E.; Fedorets, V.; Fomenko, N.; Kalymbetova, T.; Chretien, S.; Pavlov, V.; Tikunova, N.; et al. Antibiotic Resistance and Pathogenomics of Staphylococci Circulating in Novosibirsk, Russia. Microorganisms 2021, 9, 2487. https://doi.org/10.3390/microorganisms9122487
Bardasheva A, Tikunov A, Kozlova Y, Zhirakovskaia E, Fedorets V, Fomenko N, Kalymbetova T, Chretien S, Pavlov V, Tikunova N, et al. Antibiotic Resistance and Pathogenomics of Staphylococci Circulating in Novosibirsk, Russia. Microorganisms. 2021; 9(12):2487. https://doi.org/10.3390/microorganisms9122487
Chicago/Turabian StyleBardasheva, Alevtina, Artem Tikunov, Yuliya Kozlova, Elena Zhirakovskaia, Valeriya Fedorets, Natalya Fomenko, Tatyana Kalymbetova, Svetlana Chretien, Vitaliy Pavlov, Nina Tikunova, and et al. 2021. "Antibiotic Resistance and Pathogenomics of Staphylococci Circulating in Novosibirsk, Russia" Microorganisms 9, no. 12: 2487. https://doi.org/10.3390/microorganisms9122487
APA StyleBardasheva, A., Tikunov, A., Kozlova, Y., Zhirakovskaia, E., Fedorets, V., Fomenko, N., Kalymbetova, T., Chretien, S., Pavlov, V., Tikunova, N., & Morozova, V. (2021). Antibiotic Resistance and Pathogenomics of Staphylococci Circulating in Novosibirsk, Russia. Microorganisms, 9(12), 2487. https://doi.org/10.3390/microorganisms9122487