
Alteration of Salmonella enterica Virulence and Host Pathogenesis through Targeting sdiA by Using the CRISPR-Cas9 System

 , ,
, ,  , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids Enzymes, Media and Chemicals
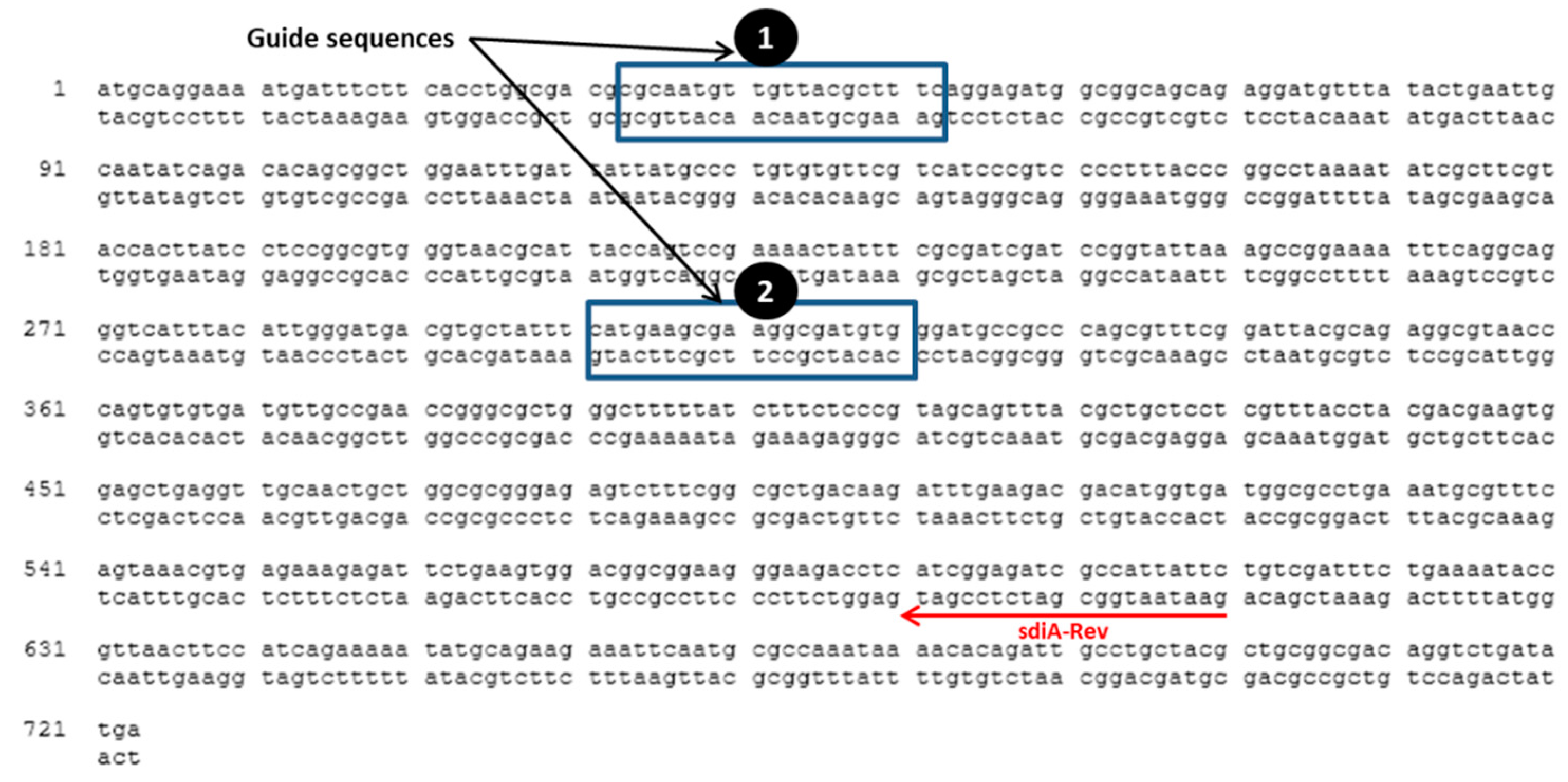
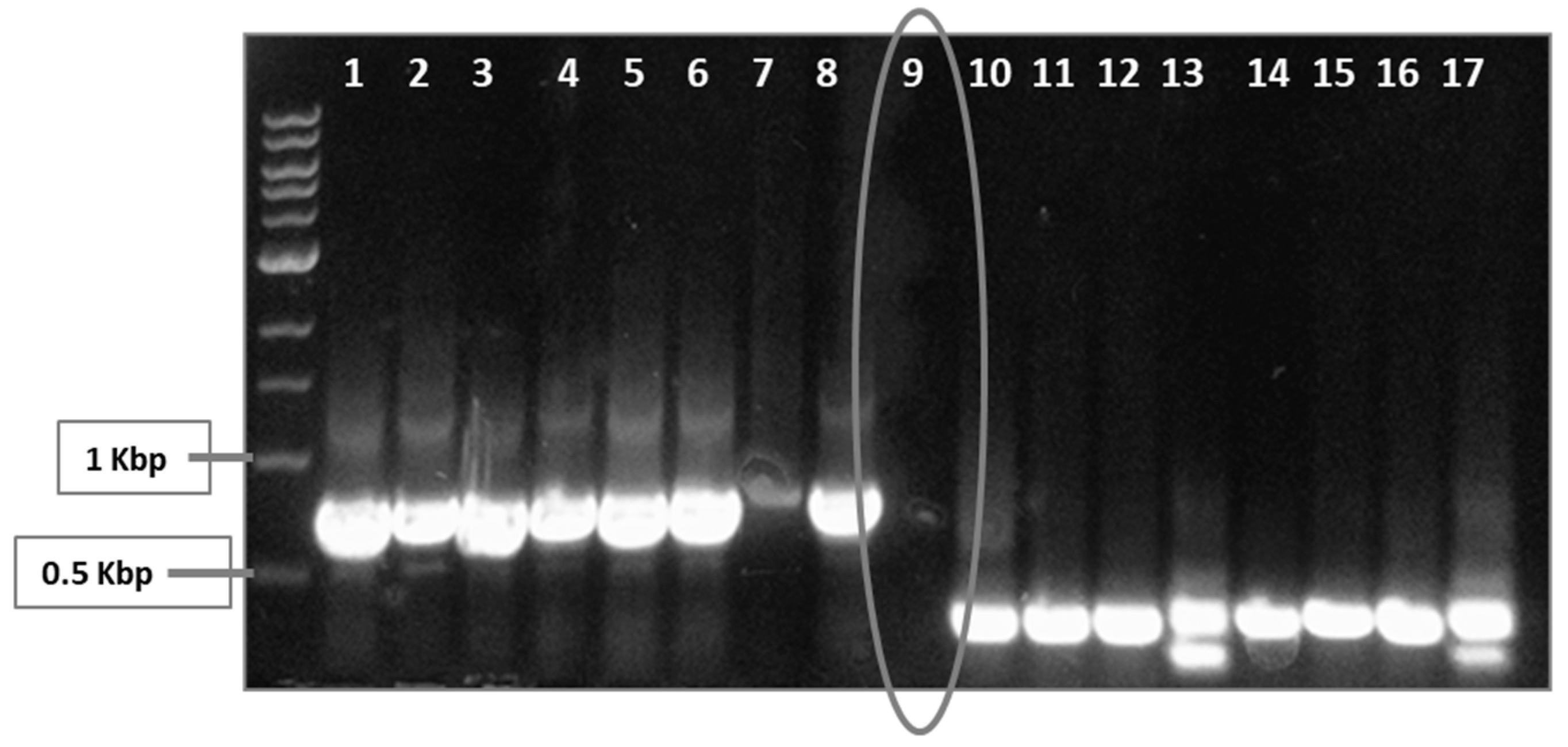
2.2. Targeting sdiA by CRISPR/Cas9
2.3. Adhesion Assay
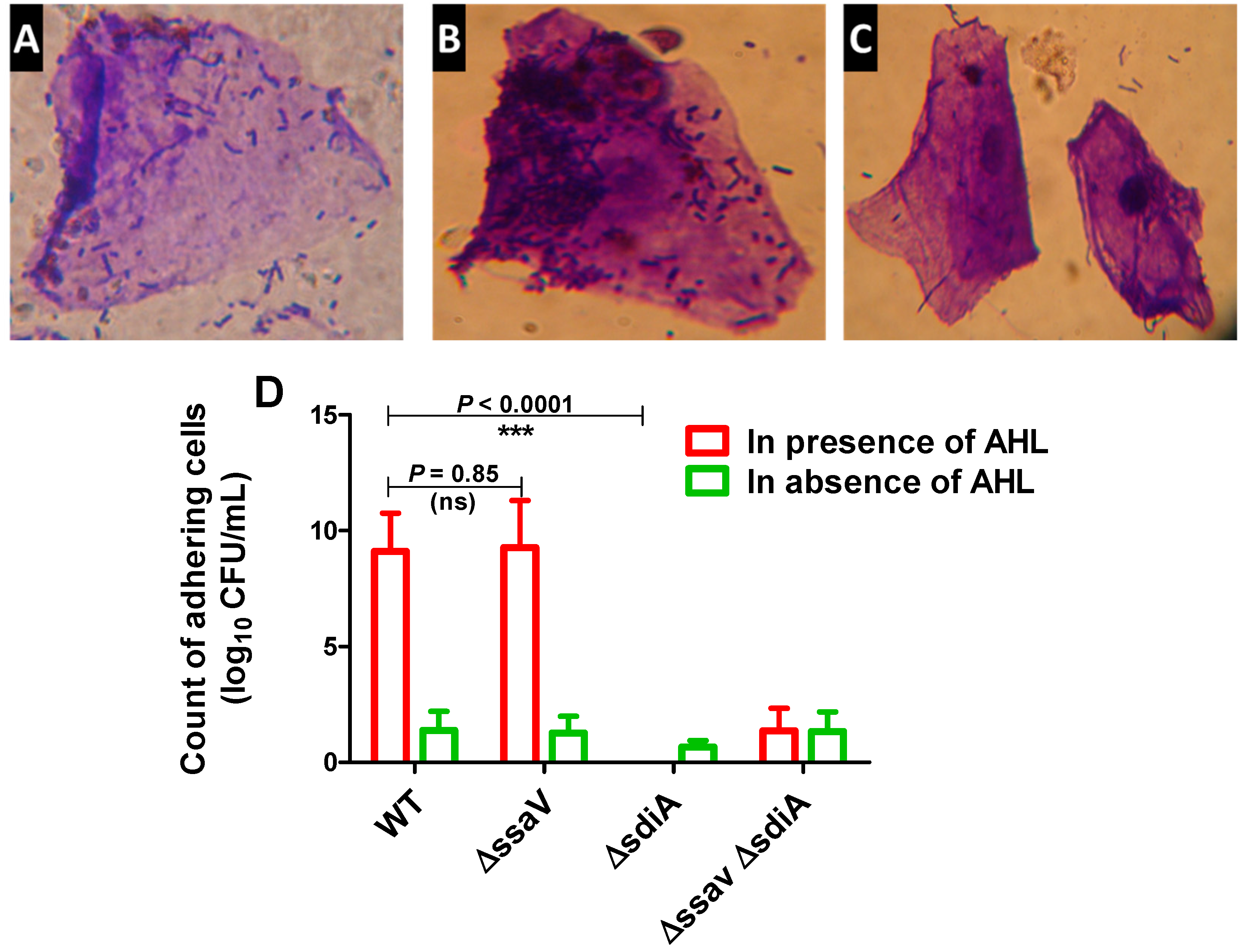
2.3.1. Adhesion to Epithelial Cells
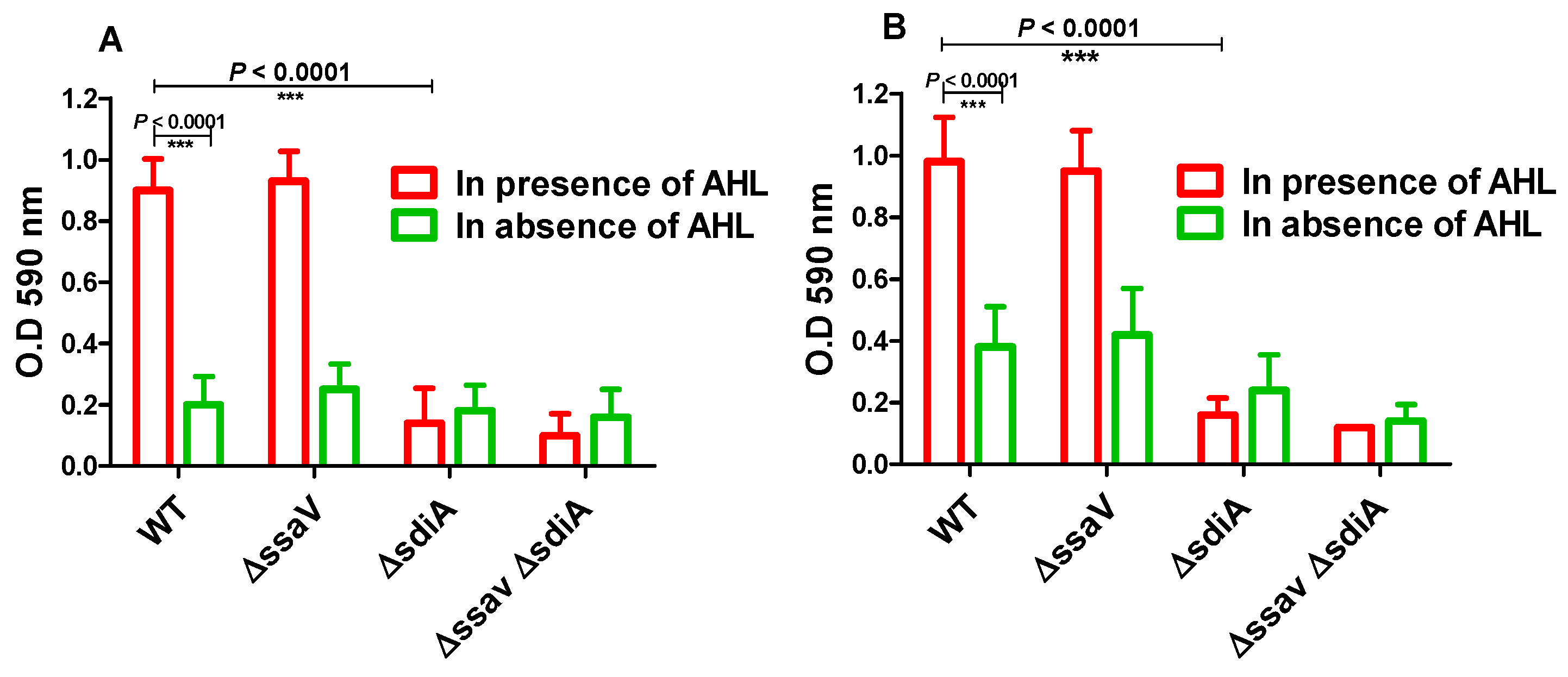
2.3.2. Adhesion to Abiotic Surface and Biofilm Formation
2.4. Invasion Assay and Intracellular Replication
2.5. The Intracellular Behavior of Salmonella Mutants
2.5.1. Construction of SPI2 Expressing Plasmid
2.5.2. Evaluation of SPI2 Effectors Expression
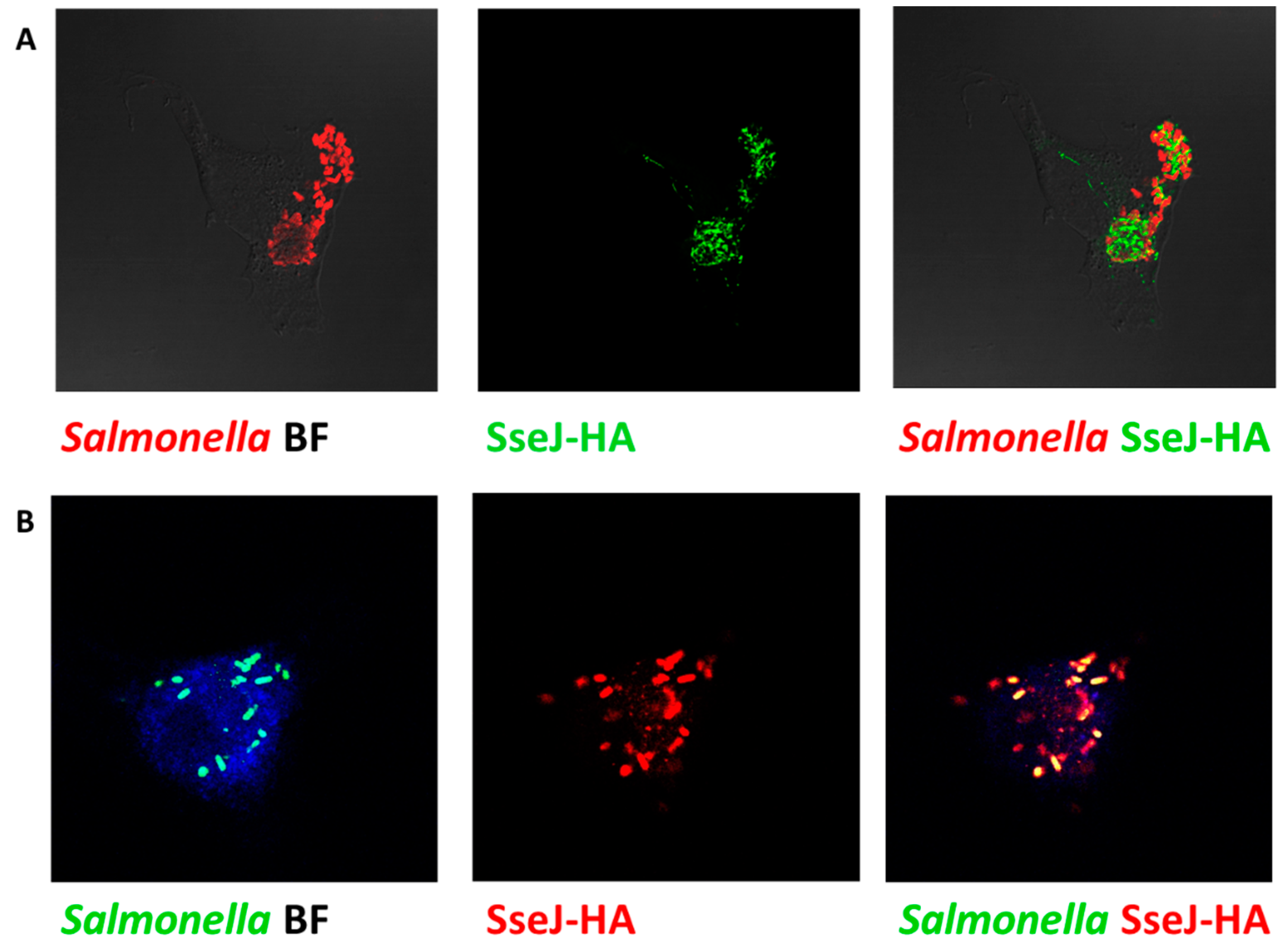
2.5.3. Evaluation of Translocation Efficiency
2.6. The Effect on Mutation on Bacterial Susceptibility to Antibiotics
2.7. In Vivo Assessment of the Pathogenesis of Tested Mutants
2.8. Statistical Analysis
3. Results
3.1. CRISPR/Cas9 System Targets sdiA
3.2. Functional Testing of S. enterica ∆sdiA
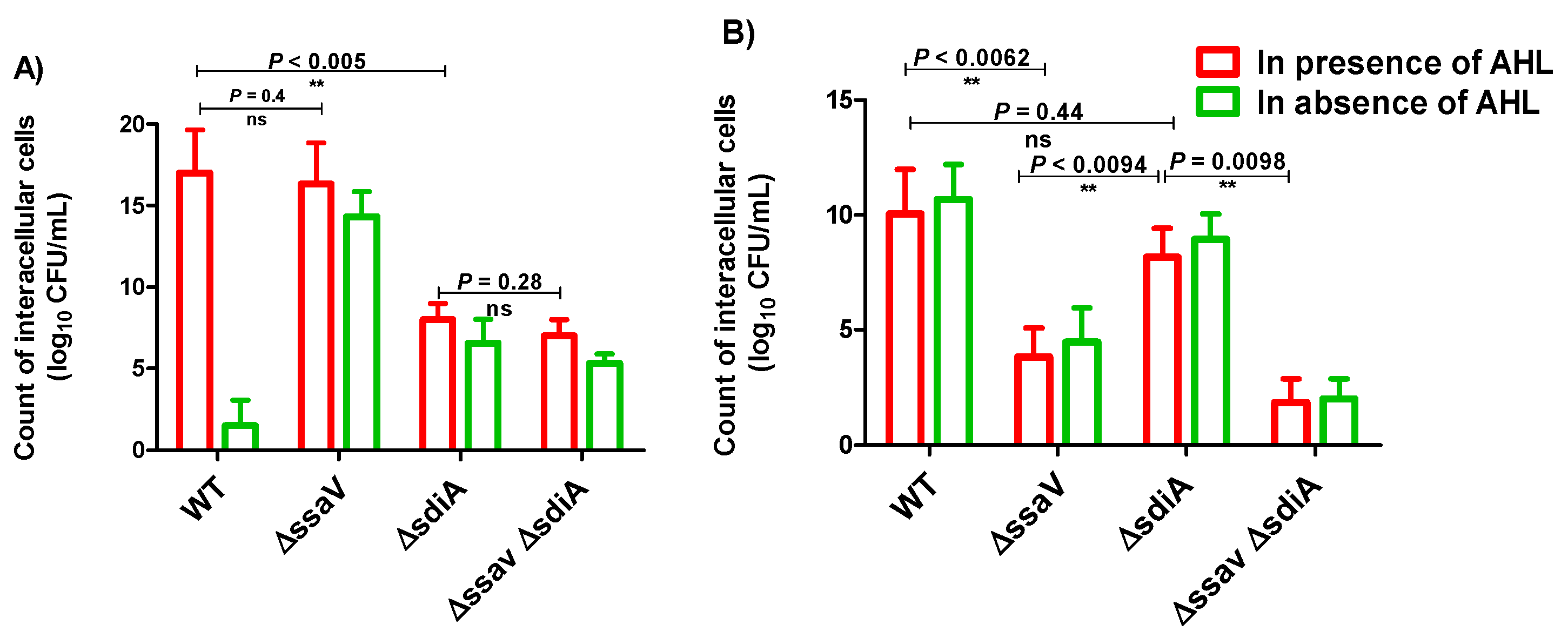
3.3. Intercellular Survival of S. enterica ΔsdiA
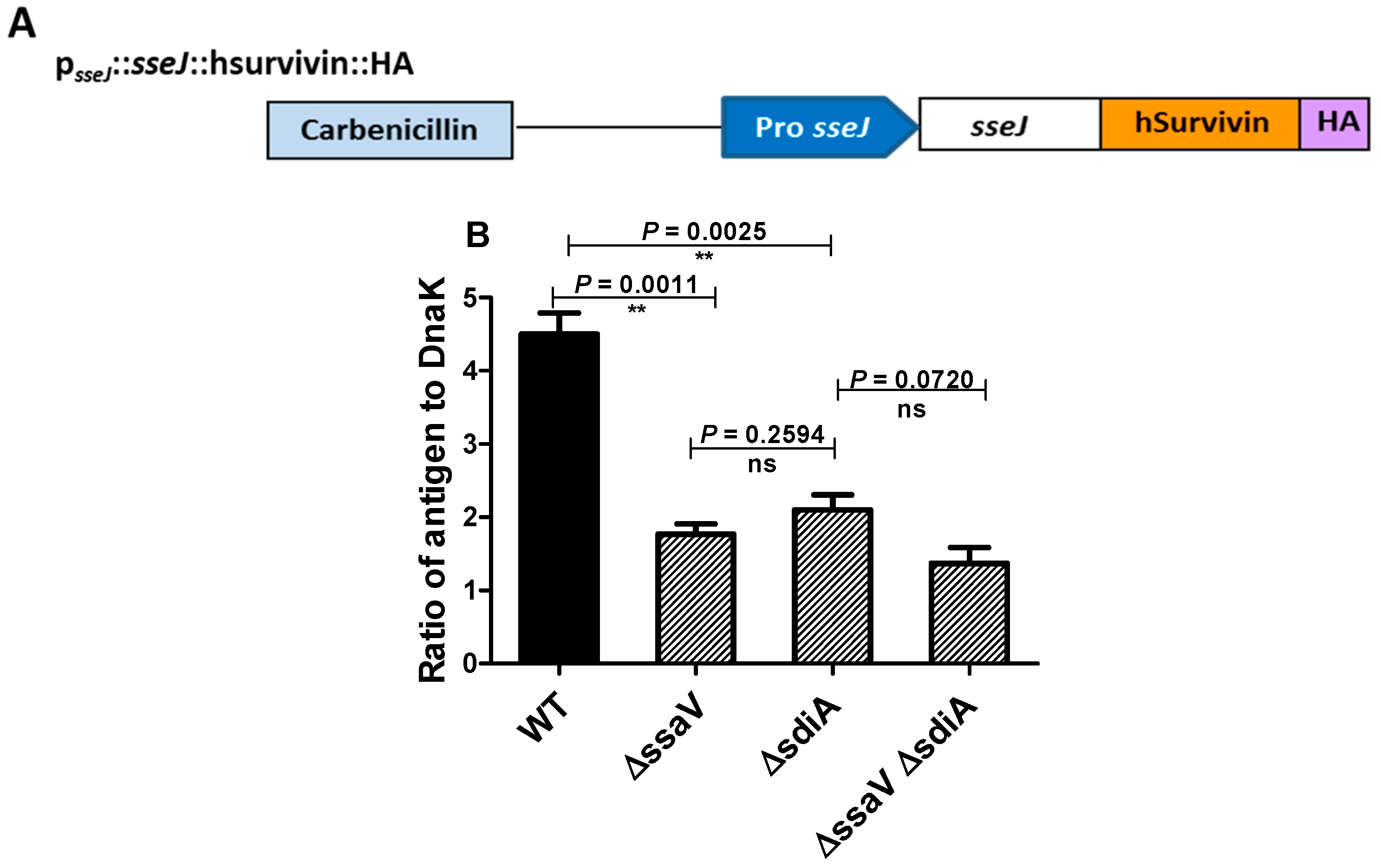
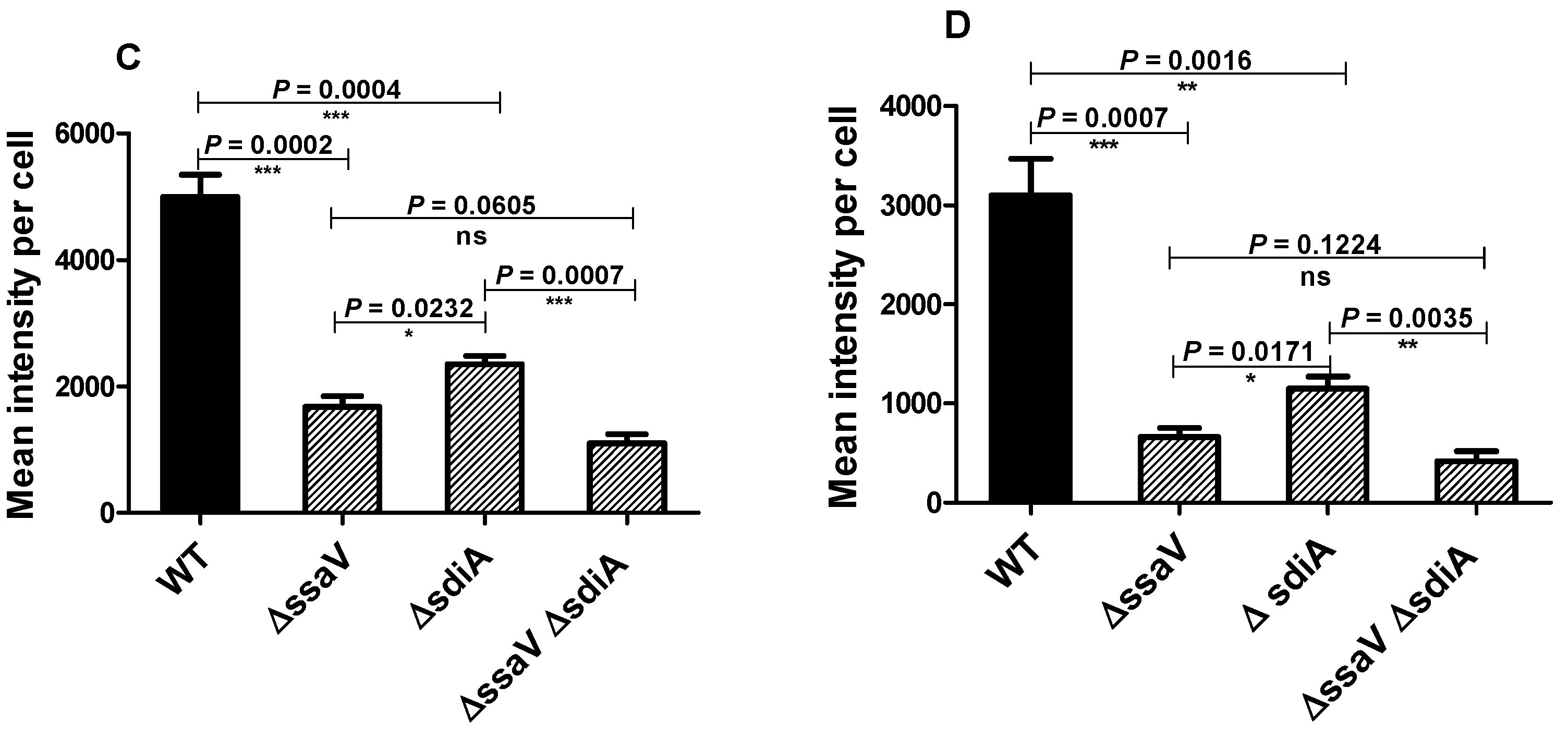
3.4. Assessment the Expression of SPI2 Effectors S. enterica Strains
3.5. MICs and MBICs of S. enterica Mutant Strains
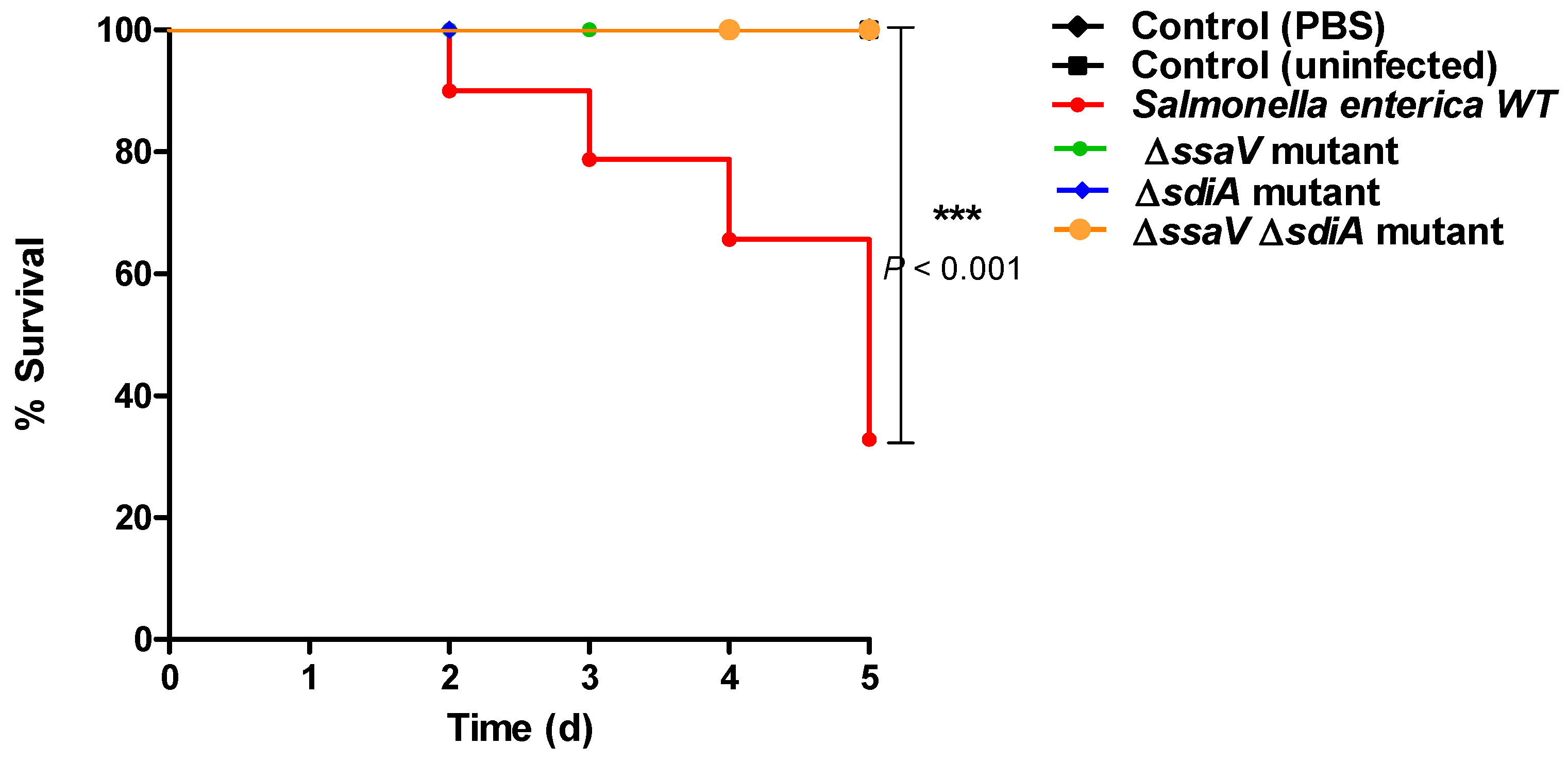
3.6. Mutation in sdiA and/or ssaV Genes Decreases S. enterica Virulence In Vivo
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hegazy, W.A.H.; Hensel, M. Salmonella enterica as a vaccine carrier. Futur. Microbiol. 2012, 7, 111–127. [Google Scholar] [CrossRef]
- Haraga, A.; Ohlson, M.B.; Miller, S.I. Salmonellae interplay with host cells. Nat. Rev. Genet. 2008, 6, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, P. Process of protein transport by the Type III secretion system. Microbiol. Mol. Biol. Rev. 2004, 68, 771–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerlach, R.G.; Hensel, M. Salmonella pathogenicity islands in host specificity, host pathogen-interactions and antibiotics re-sistance of Salmonella enterica. Berl. Munch. Tierarztl. Wochenschr. 2007, 120, 317–327. [Google Scholar]
- Askoura, M.; Hegazy, W.A.H. Ciprofloxacin interferes with Salmonella Typhimurium intracellular survival and host virulence through repression of Salmonella pathogenicity island-2 (SPI-2) genes expression. Pathog. Dis. 2020, 78, 78. [Google Scholar] [CrossRef] [PubMed]
- Deiwick, J.; Salcedo, S.; Boucrot, E.; Gilliland, S.M.; Henry, T.; Petermann, N.; Waterman, S.R.; Gorvel, J.-P.; Holden, D.W.; Méresse, S. The translocated Salmonella effector proteins SseF and SseG interact and are required to establish an intracellular replication niche. Infect. Immun. 2006, 74, 6965–6972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browne, S.H.; Hasegawa, P.; Okamoto, S.; Fierer, J.; Guiney, D.G. Identification of Salmonella SPI-2 secretion system compo-nents required for SpvB-mediated cytotoxicity in macrophages and virulence in mice. FEMS Immunol. Med. Microbiol. 2008, 52, 194–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wael, A.H.H.; Hisham, A.A.; Hegazy, W.A.H.; Abbas, H.A. Evaluation of the role of SsaV Salmonella pathogenicity island-2 dependent type III secretion system components on the virulence behavior of Salmonella enterica serovar Typhimurium. Afr. J. Biotechnol. 2017, 16, 718–726. [Google Scholar] [CrossRef] [Green Version]
- Abbas, H.A.; Hegazy, W.A.H. Repurposing anti-diabetic drug “Sitagliptin” as a novel virulence attenuating agent in Serratia marcescens. PLoS ONE 2020, 15, e0231625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegazy, W.A.H.; Khayat, M.T.; Ibrahim, T.S.; Nassar, M.S.; Bakhrebah, M.A.; Abdulaal, W.H.; Alhakamy, N.A.; Bendary, M.M. Repurposing anti-diabetic drugs to cripple Quorum sensing in Pseudomonas aeruginosa. Microorganisms 2020, 8, 1285. [Google Scholar] [CrossRef] [PubMed]
- Askoura, M.; Youns, M.; Hegazy, W.A.H. Investigating the influence of iron on Campylobacter jejuni transcriptome in re-sponse to acid stress. Microb. Pathog. 2020, 138, 103777. [Google Scholar] [CrossRef]
- Li, G.; Yan, C.; Xu, Y.; Feng, Y.; Wu, Q.; Lv, X.; Yang, B.; Wang, X.; Xia, X. Punicalagin inhibits Salmonella virulence factors and has anti-quorum-sensing potential. Appl. Environ. Microbiol. 2014, 80, 6204–6211. [Google Scholar] [CrossRef] [Green Version]
- Jiang, T.; Li, M. Quorum sensing inhibitors: A patent review. Expert Opin. Ther. Patents 2013, 23, 867–894. [Google Scholar] [CrossRef]
- Khayyat, A.; Hegazy, W.; Shaldam, M.; Mosbah, R.; Almalki, A.; Ibrahim, T.; Khayat, M.; Khafagy, E.-S.; Soliman, W.; Abbas, H. Xylitol inhibits growth and blocks virulence in Serratia marcescens. Microorganism 2021, 9, 1083. [Google Scholar] [CrossRef] [PubMed]
- Janssens, J.C.A.; Metzger, K.; Daniels, R.; Ptacek, D.; Verhoeven, T.; Habel, L.W.; Vanderleyden, J.; De Vos, D.E.; De Keersmaecker, S.C.J. Synthesis of N-Acyl Homoserine Lactone analogues reveals strong activators of SdiA, the Salmonella en-terica Serovar Typhimurium LuxR Homologue. Appl. Environ. Microbiol. 2007, 73, 535–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michael, B.; Smith, J.N.; Swift, S.; Heffron, F.; Ahmer, B.M.M. SdiA of Salmonella enterica is a LuxR Homolog that detects mixed microbial communities. J. Bacteriol. 2001, 183, 5733–5742. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.N.; Ahmer, B.M.M. Detection of other microbial species by Salmonella: Expression of the SdiA regulon. J. Bacteriol. 2003, 185, 1357–1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegazy, W.A.H.; Xu, X.; Metelitsa, L.; Hensel, M. Evaluation of Salmonella enterica Type III secretion system effector proteins as carriers for heterologous vaccine antigens. Infect. Immun. 2012, 80, 1193–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Hegazy, W.; Guo, L.; Gao, X.; Courtney, A.N.; Kurbanov, S.; Liu, D.; Tian, G.; Manuel, E.; Diamond, D.; et al. Effective cancer vaccine platform based on attenuated Salmonella and a Type III secretion system. Cancer Res. 2014, 74, 6260–6270. [Google Scholar] [CrossRef] [Green Version]
- Theys, J.; Barbe, S.; Landuyt, W.; Nuyts, S.; Mellaert, L.; Wouters, B.; Anné, J.; Lambin, P. Tumor-specific gene delivery using genetically engineered bacteria. Curr. Gene Ther. 2003, 3, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Wiedenheft, B.; Sternberg, S.H.; Doudna, J.A. RNA-guided genetic silencing systems in bacteria and archaea. Nat. Cell Biol. 2012, 482, 331–338. [Google Scholar] [CrossRef]
- Charpentier, E.; Doudna, J.A. Rewriting a genome. Nat. Cell Biol. 2013, 495, 50–51. [Google Scholar] [CrossRef]
- Deveau, H.; Garneau, J.E.; Moineau, S. CRISPR/Cas system and its role in phage-bacteria interactions. Annu. Rev. Microbiol. 2010, 64, 475–493. [Google Scholar] [CrossRef] [PubMed]
- Horvath, P.; Barrangou, R. CRISPR/Cas, the immune system of bacteria and Archaea. Science 2010, 327, 167–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Bikard, D.; Cox, D.; Zhang, F.; Marraffini, A.L. RNA-guided editing of bacterial genomes using CRISPR-Cas sys-tems. Nat. Biotechnol. 2013, 31, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Chen, B.; Duan, C.; Sun, B.; Yang, J.; Yang, S. Multigene editing in the Escherichia coli genome via the CRISPR-Cas9 system. Appl. Environ. Microbiol. 2015, 81, 2506–2514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, J.-H.; Van Pijkeren, J.-P. CRISPR–Cas9-assisted recombineering in Lactobacillus reuteri. Nucleic Acids Res. 2014, 42, e131. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, Z.; Seo, S.-O.; Choi, K.; Lu, T.; Jin, Y.-S.; Blaschek, H.P. Markerless chromosomal gene deletion in Clostrid-ium beijerinckii using CRISPR/Cas9 system. J. Biotechnol. 2015, 200, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Chung, J.-H.; Kim, H.M.; Kim, D.-W.; Kim, H.H. Designed nucleases for targeted genome editing. Plant. Biotechnol. J. 2015, 14, 448–462. [Google Scholar] [CrossRef]
- Xie, K.; Yang, Y. RNA-guided genome editing in plants using a CRISPR–Cas system. Mol. Plant. 2013, 6, 1975–1983. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and applications of CRISPR-Cas9 for genome engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef] [Green Version]
- Braukmann, M.; Methner, U.; Berndt, A. Immune reaction and survivability of Salmonella Typhimurium and Salmonella Infantis after infection of primary avian macrophages. PLoS ONE 2015, 10, e0122540. [Google Scholar] [CrossRef] [Green Version]
- Hegazy, W.; Youns, M. TALENs construction: Slowly but surely. Asian Pac. J. Cancer Prev. 2016, 17, 3329–3334. [Google Scholar]
- Bolotin, A.; Quinquis, B.; Sorokin, A.; Ehrlich, S.D. Clustered regularly interspaced short palindrome repeats (CRISPRs) have spacers of extrachromosomal origin. Microbiology 2005, 151, 2551–2561. [Google Scholar] [CrossRef] [Green Version]
- Mojica, F.J.M.; Díez-Villaseñor, C.; García-Martínez, J.; Soria, E. Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements. J. Mol. Evol. 2005, 60, 174–182. [Google Scholar] [CrossRef]
- Pourcel, C.; Salvignol, G.; Vergnaud, G. CRISPR elements in Yersinia pestis acquire new repeats by preferential uptake of bac-teriophage DNA, and provide additional tools for evolutionary studies. Microbiology 2005, 151, 653–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marraffini, L.A.; Sontheimer, E.J. CRISPR interference limits horizontal gene transfer in Staphylococci by targeting DNA. Sci-ence 2008, 322, 1843–1845. [Google Scholar] [CrossRef] [Green Version]
- Heler, R.; Marraffini, L.A.; Bikard, D. Adapting to new threats: The generation of memory by CRISPR-Cas immune systems. Mol. Microbiol. 2014, 93, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Makarova, K.S.; Haft, D.H.; Barrangou, R.; Brouns, S.J.; Charpentier, E.; Horvath, P.; Moineau, S.; Mojica, F.J.; Wolf, Y.; Ya-kunin, A.; et al. Evolution and classification of the CRISPR–Cas systems. Nat. Rev. Genet. 2011, 9, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Gratz, S.J.; Wildonger, J.; Harrison, M.M.; O’Connor-Giles, K.M. CRISPR/Cas9-mediated genome engineering and the promise of designer flies on demand. Fly 2013, 7, 249–255. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Marraffini, L.A. CRISPR-Cas: New tools for genetic manipulations from bacterial immunity systems. Annu. Rev. Microbiol. 2015, 69, 209–228. [Google Scholar] [CrossRef]
- Huang, H.; Zheng, G.; Jiang, W.; Hu, H.; Lu, Y. One-step high-efficiency CRISPR/Cas9-mediated genome editing in Strepto-myces. Acta Biochim. Biophys. Sin. 2015, 47, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Fierer, J.; Okamoto, S.; Banerjee, A.; Guiney, D.G. Diarrhea and Colitis in mice require the Salmonella Pathogenicity Island 2-Encoded secretion function but Not SifA or Spv effectors. Infect. Immun. 2012, 80, 3360–3370. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhang, Z.; Pace, L.; Lillehoj, H.; Zhang, S. Functions exerted by the virulence-associated type-three secretion systems during Salmonella enterica serovar Enteritidis invasion into and survival within chicken oviduct epithelial cells and macro-phages. Avian Pathol. 2009, 38, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Vesterlund, S.; Paltta, J.; Karp, M.; Ouwehand, A. Measurement of bacterial adhesion—In vitro evaluation of different meth-ods. J. Microbiol. Methods 2005, 60, 225–233. [Google Scholar] [CrossRef]
- Schmidt, M.; Olejnik-Schmidt, A.K.; Myszka, K.; Borkowska, M.; Grajek, W. Evaluation of quantitative PCR Measurement of bacterial colonization of Epithelial cells. Pol. J. Microbiol. 2010, 59, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Khayyat, A.; Abbas, H.; Mohamed, M.; Asfour, H.; Khayat, M.; Ibrahim, T.; Youns, M.; Khafagy, E.-S.; Abu Lila, A.; Safo, M.; et al. Not only antimicrobial: Metronidazole Mitigates the virulence of Proteus mirabilis isolated from macerated diabetic foot ulcer. Appl. Sci. 2021, 11, 6847. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Dakić, I.; Savić, B.; Švabić-Vlahović, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Al Saqr, A.; Aldawsari, M.F.; Khafagy, E.-S.; Shaldam, M.A.; Hegazy, W.A.H.; Abbas, H.A. A novel use of Allopurinol as a quorum-sensing inhibitor in Pseudomonas aeruginosa. Antibiotics 2021, 10, 1385. [Google Scholar] [CrossRef]
- Hölzer, S.U.; Hensel, M. Divergent roles of Salmonella Pathogenicity Island 2 and metabolic traits during interaction of S. en-terica Serovar Typhimurium with host cells. PLoS ONE 2012, 7, e33220. [Google Scholar] [CrossRef]
- Aldawsari, M.; Khafagy, E.-S.; Saqr, A.; Alalaiwe, A.; Abbas, H.; Shaldam, M.; Hegazy, W.; Goda, R. Tackling virulence of Pseudomonas aeruginosa by the Natural Furanone Sotolon. Antibiotics 2021, 10, 871. [Google Scholar] [CrossRef] [PubMed]
- Al Saqr, A.; Khafagy, E.-S.; Alalaiwe, A.; Aldawsari, M.; Alshahrani, S.; Anwer, K.; Khan, S.; Lila, A.; Arab, H.; Hegazy, W. Synthesis of gold nanoparticles by using green machinery: Characterization and in vitro toxicity. Nanomaterials 2021, 11, 808. [Google Scholar] [CrossRef] [PubMed]
- Khayyat, A.N.; Abbas, H.A.; Khayat, M.T.; Shaldam, M.A.; Askoura, M.; Asfour, H.Z.; Khafagy, E.-S.; Abu Lila, A.S.; Allam, A.N.; Hegazy, W.A.H. Secnidazole is a promising imidazole mitigator of Serratia marcescens virulence. Microorganism 2021, 9, 2333. [Google Scholar] [CrossRef] [PubMed]
- Hegazy, W.A.; Khayat, M.T.; Ibrahim, T.S.; Youns, M.; Mosbah, R.; Soliman, W.E. Repurposing of antidiabetics as Serratia marcescens virulence inhibitors. Braz. J. Microbiol. 2021, 52, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Bendary, M.M.; Ibrahim, D.; Mosbah, R.A.; Mosallam, F.; Hegazy, W.A.H.; Awad, N.F.S.; Alshareef, W.A.; Al Omar, S.Y.; Zaitone, S.A.; El-Hamid, M.I.A. Thymol Nanoemulsion: A new therapeutic option for extensively drug resistant foodborne pathogens. Antibiotics 2020, 10, 25. [Google Scholar] [CrossRef]
- Rychlik, I.; Barrow, P.A. Salmonella stress management and its relevance to behaviour during intestinal colonisation and infec-tion. FEMS Microbiol. Rev. 2005, 29, 1021–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, A.J.; Ahmer, B.M. Detection of acyl-homoserine lactones by Escherichia and Salmonella. Curr. Opin. Microbiol. 2011, 14, 188–193. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.N.; Dyszel, J.L.; Soares, J.A.; Ellermeier, C.D.; Altier, C.; Lawhon, S.D.; Adams, L.G.; Konjufca, V.; Curtiss, R.; Slauch, J.M.; et al. SdiA, an N-Acylhomoserine lactone receptor, becomes active during the transit of Salmonella enterica through the gastrointestinal tract of turtles. PLoS ONE 2008, 3, e2826. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Que, F.; Liao, L.; Zhou, M.; You, L.; Zhao, Q.; Li, Y.; Niu, H.; Wu, S.; Huang, R. Study on the promotion of bacterial biofilm formation by a Salmonella Conjugative Plasmid and the Underlying mechanism. PLoS ONE 2014, 9, e109808. [Google Scholar] [CrossRef] [Green Version]
- Marraffini, L.A.; Sontheimer, E.J. CRISPR interference: RNA-directed adaptive immunity in bacteria and archaea. Nat. Rev. Genet. 2010, 11, 181–190. [Google Scholar] [CrossRef]
- Nesse, L.L.; Berg, K.; Vestby, L.K.; Olsaker, I.; Djønne, B. Salmonella Typhimurium invasion of HEp-2 epithelial cells in vitro is increased by N-acylhomoserine lactone quorum sensing signals. Acta Vet. Scand. 2011, 53, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouwman, C.W.; Kohli, M.; Killoran, A.; Touchie, G.A.; Kadner, R.J.; Martin, N.L. Characterization of SrgA, a Salmonella en-terica Serovar Typhimurium virulence plasmid-encoded paralogue of the Disulfide Oxidoreductase DsbA, essential for bio-genesis of plasmid-encoded fimbriae. J. Bacteriol. 2003, 185, 991–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miki, T.; Okada, N.; Danbara, H. Two periplasmic disulfide oxidoreductases, DsbA and SrgA, target outer membrane protein SpiA, a component of the Salmonella Pathogenicity Island 2 Type III secretion system. J. Biol. Chem. 2004, 279, 34631–34642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pati, N.B.; Vishwakarma, V.; Selvaraj, S.K.; Dash, S.; Saha, B.; Singh, N.; Suar, M. Salmonella Typhimurium TTSS-2 deficient mig-14 mutant shows attenuation in immunocompromised mice and offers protection against wild-type Salmonella Typhi-murium infection. BMC Microbiol. 2013, 13, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Zhang, M.Z.; Yan, L.; Lillehoj, H.; Pace, L.W.; Zhang, S. Induction of CXC Chemokine messenger-RNA expression in chicken oviduct epithelial cells by Salmonella enterica serovar Enteritidis via the type three secretion system–1. Avian Dis. 2009, 53, 396–404. [Google Scholar] [CrossRef]
- Periaswamy, B.; Maier, L.; Vishwakarma, V.; Slack, E.; Kremer, M.; Andrews-Polymenis, H.L.; McClelland, M.; Grant, A.J.; Suar, M.; Hardt, W.-D. Live attenuated S. Typhimurium vaccine with improved safety in immuno-compromised mice. PLoS ONE 2012, 7, e45433. [Google Scholar] [CrossRef] [Green Version]
- Aldawsari, M.; Alalaiwe, A.; Khafagy, E.-S.; Al Saqr, A.; Alshahrani, S.; Alsulays, B.; Alshehri, S.; Abu Lila, A.; Rizvi, S.D.; Hegazy, W. Efficacy of SPG-ODN 1826 Nanovehicles in inducing M1 phenotype through TLR-9 activation in murine alveolar J774A.1 cells: Plausible nano-immunotherapy for lung carcinoma. Int. J. Mol. Sci. 2021, 22, 6833. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligonucleotide | Sequence (5′–3′) |
|---|---|
| Oligo I | AAAC CGCAATGTTGTTACGCTTTC G |
| Oligo II | AAAAC GAAAGCGTAACAACATTGCG |
| Oligo III | AAAC CATGAAGCGAAGGCGATGTG G |
| Oligo IV | AAAAC CACATCGCCTTCGCTTCATG |
| sdiA-Rev | GAA TAA TGG CGA TCT CCG AT |
| Seq-primer | CCATAAAATATGCAGGAAA |
| hSurv-For-EcoRV | TACGATATCGGTGCCCCGACGTTGCCCCC |
| hSurvivin-HA-Rev-XbaI | ATTTCTAGATTAAGCGTAGTCTGGGACGTCGTATGGGTAATCCATAGCAGCCAGCTGCTC |
| SseJ-Pro-For-KpnI | TACGGTACCTCACATAAAACACTAGCAC |
| SseJ-Rev-EcoRV | ACGGATATCTTCAGTGGAATAATGATGAGC |
| T7-Seq | TAATACGACTCACTATAGGG |
| T3-Seq | AATTAACCCTCACTAAAGG |
| Antibiotic | WT | ΔssaV | ΔsdiA | ΔssaVΔsdiA | ||||
|---|---|---|---|---|---|---|---|---|
| MIC | MBIC | MIC | MBIC | MIC | MBIC | MIC | MBIC | |
| Ampicillin | 256 | 2048 | 128 | 2048 | 128 | 1024 | 64 | 512 |
| Ampicillin/Sulbactam | 128 | 1024 | 32 | 512 | 32 | 512 | 16 | 128 |
| Amoxicillin/clavulanic acid | 128 | 1024 | 64 | 512 | 32 | 512 | 32 | 256 |
| Piperacillin | 32 | 256 | 8 | 32 | 8 | 16 | 8 | 16 |
| Azetronam | 32 | 512 | 32 | 256 | 16 | 128 | 8 | 128 |
| Imipenem | 4 | 8 | 2 | 4 | 4 | 4 | 2 | 4 |
| Cephardine | 64 | 512 | 16 | 256 | 32 | 512 | 16 | 256 |
| Ceftazidime | 32 | 1024 | 8 | 256 | 8 | 128 | 4 | 64 |
| Cefotaxime | 16 | 256 | 4 | 64 | 4 | 64 | 4 | 64 |
| Cefepime | 8 | 128 | 4 | 32 | 2 | 16 | 2 | 16 |
| Ciprofloxacin | 8 | 12 | 2 | 4 | 2 | 4 | 1 | 2 |
| Levofloxacin | 4 | 16 | 2 | 4 | 1 | 2 | 1 | 2 |
| Gatifloxacin | 4 | 16 | 2 | 8 | 2 | 8 | 1 | 4 |
| Tobramycin | 16 | 512 | 2 | 128 | 2 | 64 | 2 | 64 |
| Gentamycin | 16 | 512 | 2 | 64 | 2 | 32 | 2 | 32 |
| Tetracycline | 64 | 1024 | 8 | 512 | 16 | 512 | 8 | 512 |
| Chloramphenicol | 64 | 1024 | 16 | 512 | 8 | 256 | 8 | 256 |
| Trimehoprim/Sulfamethoxazole | 128 | 2048 | 64 | 1024 | 32 | 512 | 16 | 512 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Askoura, M.; Almalki, A.J.; Lila, A.S.A.; Almansour, K.; Alshammari, F.; Khafagy, E.-S.; Ibrahim, T.S.; Hegazy, W.A.H. Alteration of Salmonella enterica Virulence and Host Pathogenesis through Targeting sdiA by Using the CRISPR-Cas9 System. Microorganisms 2021, 9, 2564. https://doi.org/10.3390/microorganisms9122564
Askoura M, Almalki AJ, Lila ASA, Almansour K, Alshammari F, Khafagy E-S, Ibrahim TS, Hegazy WAH. Alteration of Salmonella enterica Virulence and Host Pathogenesis through Targeting sdiA by Using the CRISPR-Cas9 System. Microorganisms. 2021; 9(12):2564. https://doi.org/10.3390/microorganisms9122564
Chicago/Turabian StyleAskoura, Momen, Ahmad J. Almalki, Amr S. Abu Lila, Khaled Almansour, Farhan Alshammari, El-Sayed Khafagy, Tarek S. Ibrahim, and Wael A. H. Hegazy. 2021. "Alteration of Salmonella enterica Virulence and Host Pathogenesis through Targeting sdiA by Using the CRISPR-Cas9 System" Microorganisms 9, no. 12: 2564. https://doi.org/10.3390/microorganisms9122564
APA StyleAskoura, M., Almalki, A. J., Lila, A. S. A., Almansour, K., Alshammari, F., Khafagy, E. -S., Ibrahim, T. S., & Hegazy, W. A. H. (2021). Alteration of Salmonella enterica Virulence and Host Pathogenesis through Targeting sdiA by Using the CRISPR-Cas9 System. Microorganisms, 9(12), 2564. https://doi.org/10.3390/microorganisms9122564