
Energy Conservation in the Acetogenic Bacterium Clostridium aceticum

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Conditions for Growth of C. aceticum
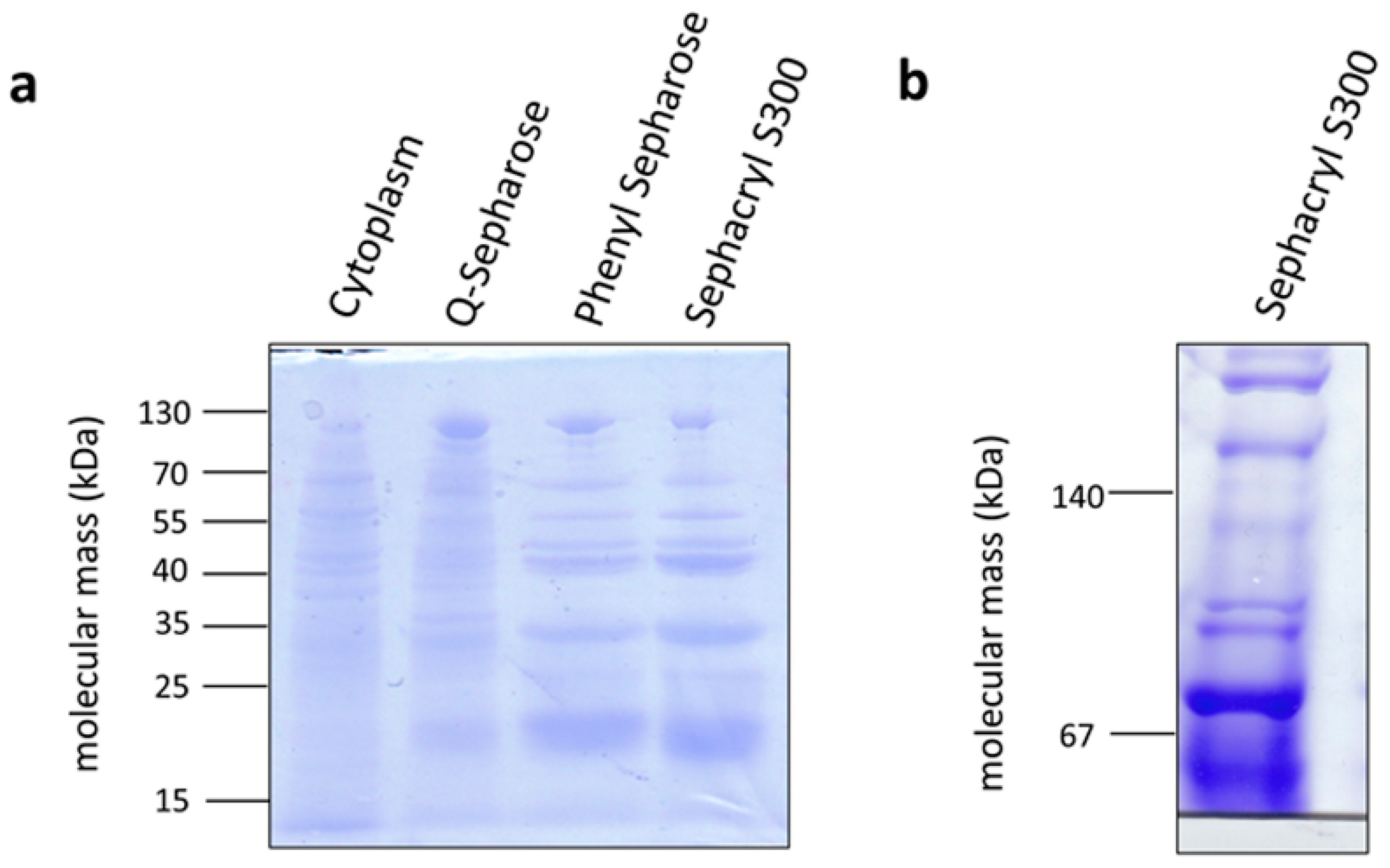
2.2. Purification of Cytosol and Membranes from C. aceticum
2.3. Purification of the Methylene-Tetrahydrofolate Reductase
2.4. Measurement of Rnf Activity
2.5. Measurement of ATPase Activity
2.6. Measurements of Methylene-Tetrahydrofolate Reductase Activity
2.7. Measurements of Methylene-Tetrahydrofolate Dehydrogenase Activity
2.8. Analytical Methods
3. Results
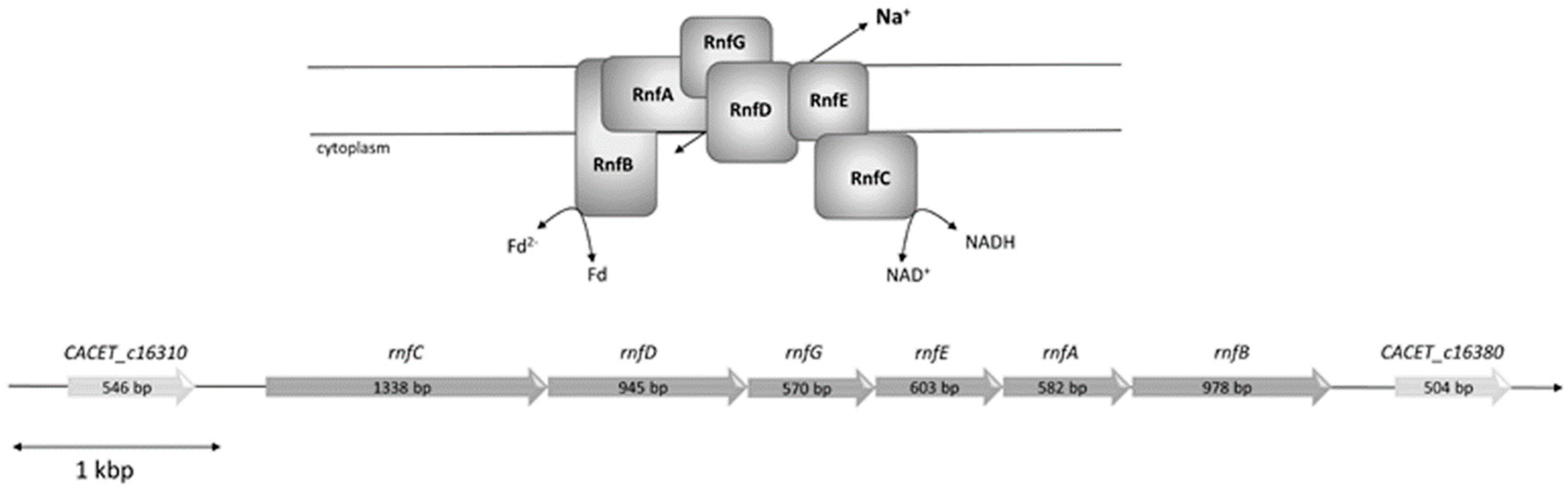
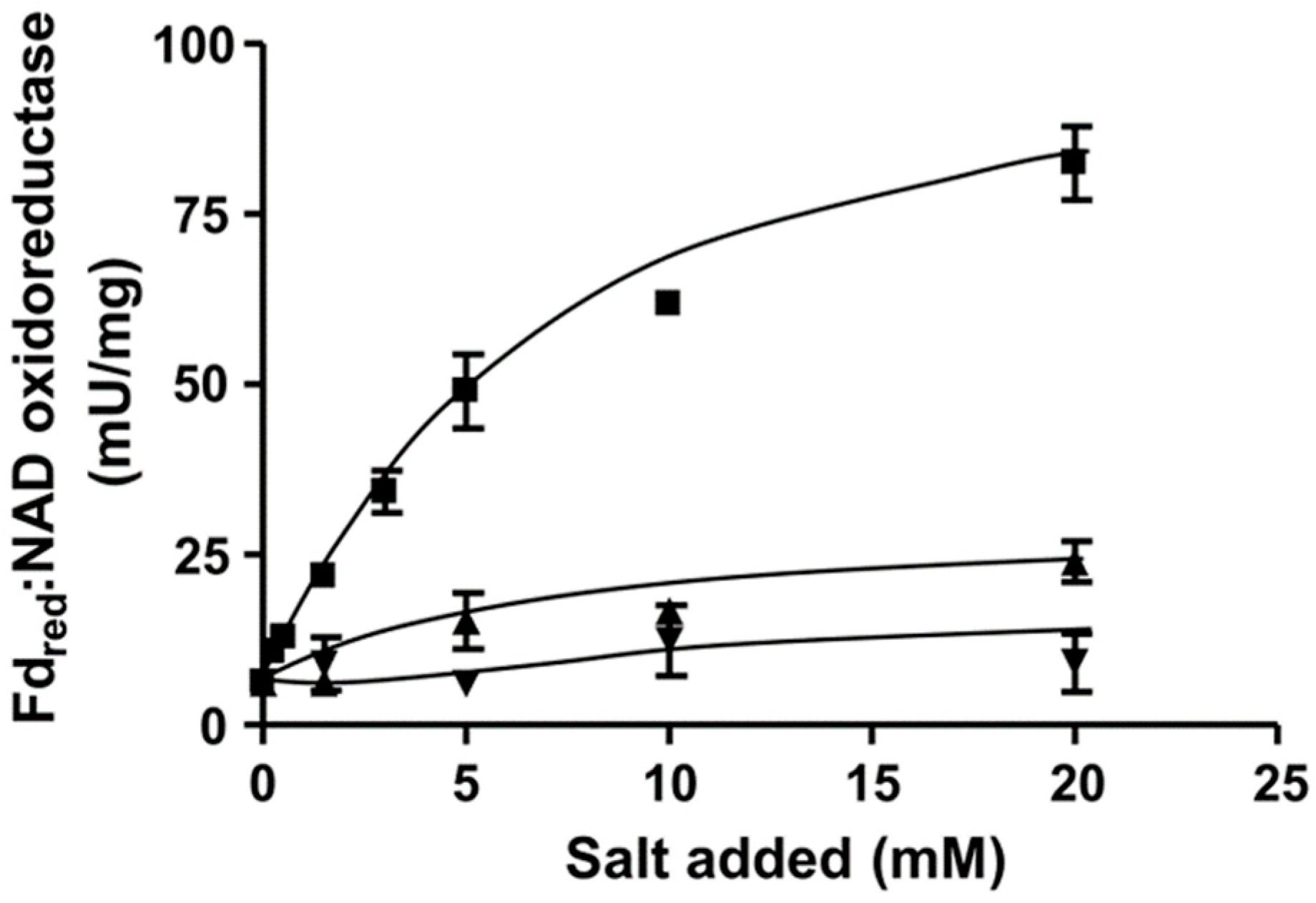
3.1. The Rnf Complex from C. aceticum Requires Na+ for Activity
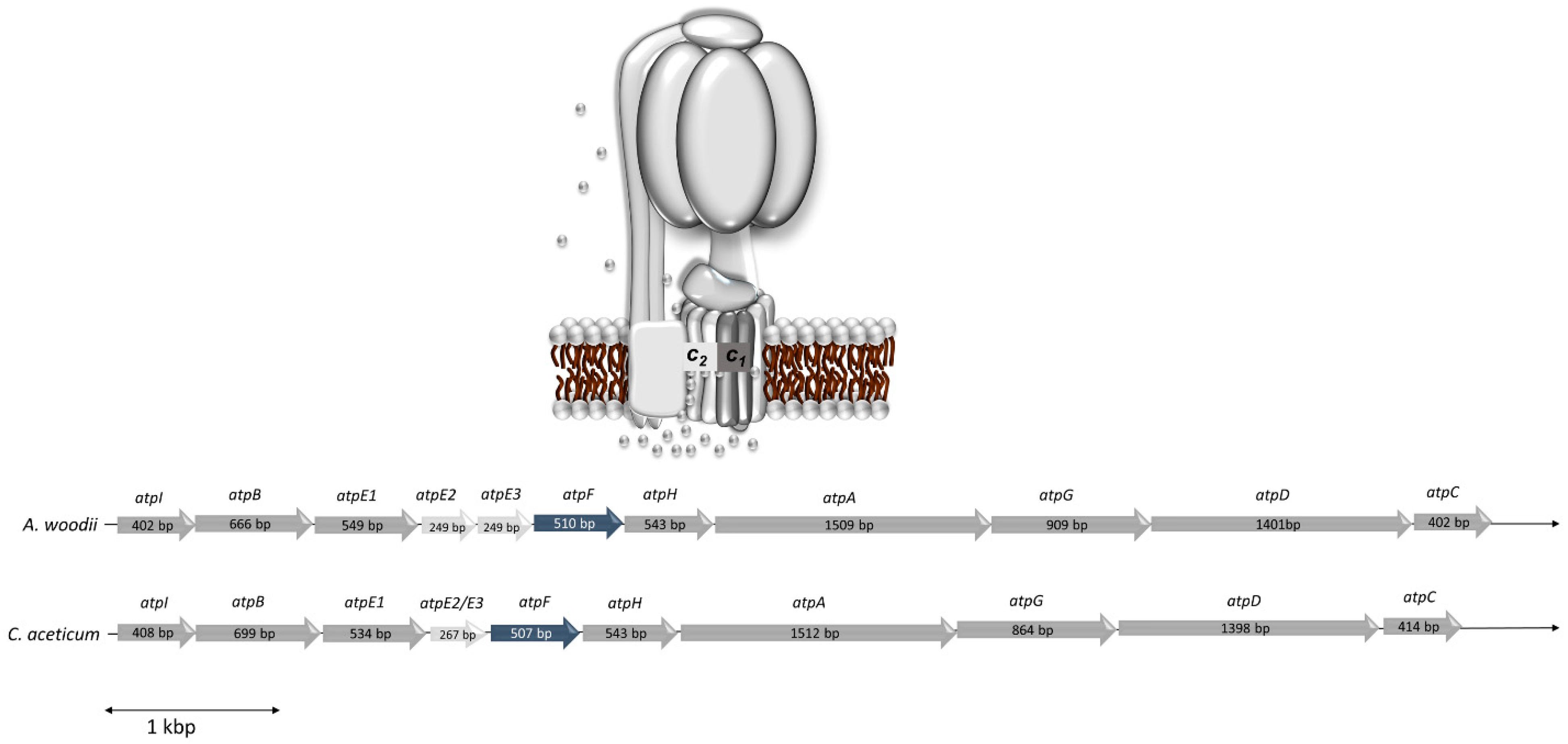
3.2. The ATP Synthase from C. aceticum Requires Na+ for Activity
3.3. The Methylene-THF Reductase Is of the MetF/MetV-Type
3.4. The Methylene-THF Dehydrogenase Is NAD Dependent
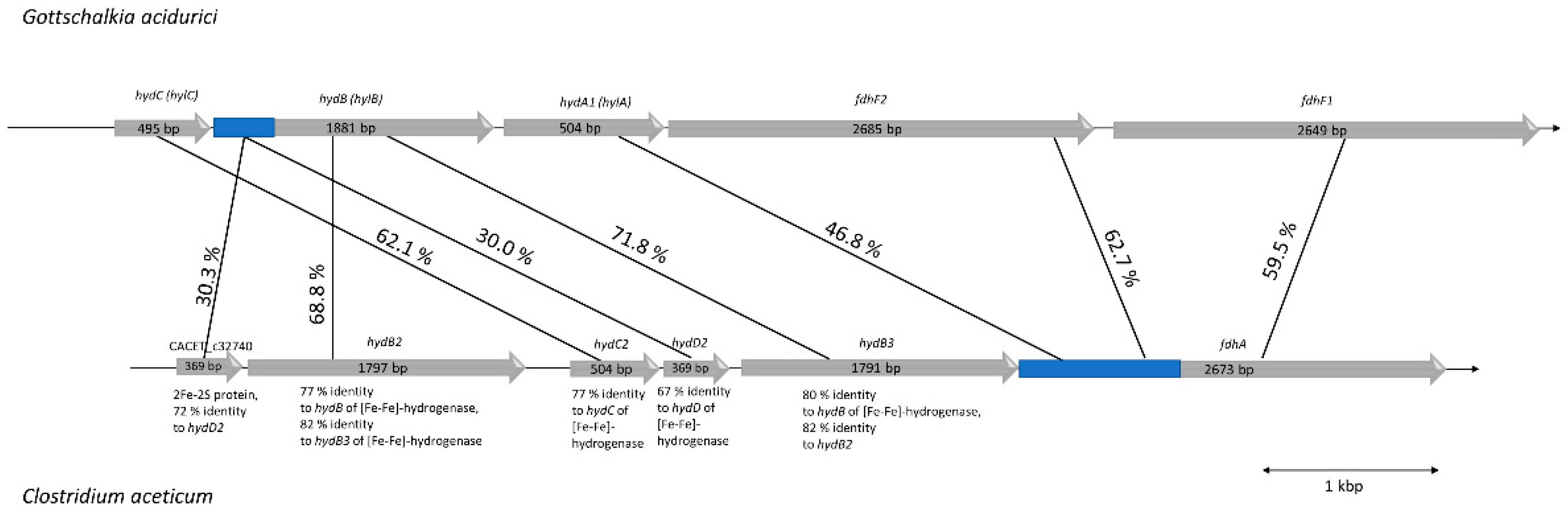
3.5. C. aceticum Has an Electron-Bifurcating Formate Dehydrogenase
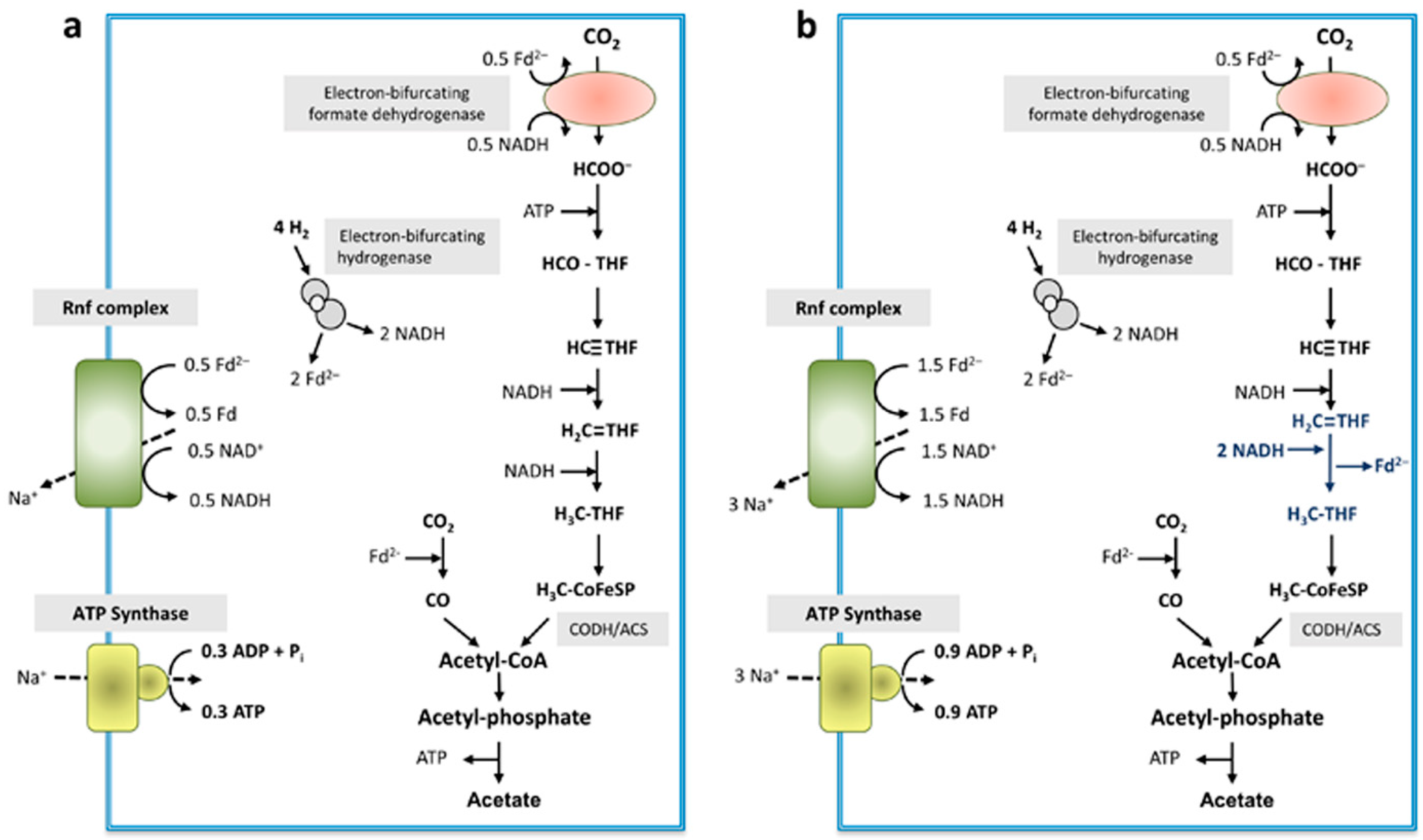
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Drake, H.L. Acetogenesis, Acetogenic Bacteria, and the Acetyl-CoA “Wood/Ljungdahl” Pathway: Past and Current Perspectives. In Acetogenesis; Drake, H.L., Ed.; Springer: New York, NY, USA, 1994; pp. 3–60. [Google Scholar]
- Ljungdahl, L.G. The Acetyl-CoA Pathway and the Chemiosmotic Generation of ATP during Acetogenesis. In Acetogenesis; Drake, H.L., Ed.; Chapman & Hall: New York, NY, USA, 1994; pp. 63–87. [Google Scholar]
- Ragsdale, S.W.; Pierce, E. Acetogenesis and the Wood-Ljungdahl Pathway of CO2 fixation. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2008, 1784, 1873–1898. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, G. Alternative Pathways of Carbon Dioxide Fixation: Insights into the Early Evolution of Life? Annu. Rev. Microbiol. 2011, 65, 631–658. [Google Scholar] [CrossRef] [PubMed]
- Ljungdahl, L.G. The Autotrophic Pathway of Acetate Synthesis in Acetogenic Bacteria. Annu. Rev. Microbiol. 1986, 40, 415–450. [Google Scholar] [CrossRef] [PubMed]
- Schuchmann, K.; Müller, V. Autotrophy at the thermodynamic limit of life: A model for energy conservation in acetogenic bacteria. Nat. Rev. Microbiol. 2014, 12, 809–821. [Google Scholar] [CrossRef] [PubMed]
- Drake, H.L.; Gößner, A.S.; Daniel, S.L. Old Acetogens, New Light. Ann. N. Y. Acad. Sci. 2008, 1125, 100–128. [Google Scholar] [CrossRef] [PubMed]
- Ragsdale, S.W. Enzymology of the Wood-Ljungdahl Pathway of Acetogenesis. Ann. N. Y. Acad. Sci. 2008, 1125, 129–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoelmerich, M.C.; Müller, V. Energy-converting hydrogenases: The link between H2 metabolism and energy conservation. Cell. Mol. Life Sci. 2020, 77, 1461–1481. [Google Scholar] [CrossRef]
- Biegel, E.; Müller, V. Bacterial Na+-translocating ferredoxin:NAD+ oxidoreductase. Proc. Natl. Acad. Sci. USA 2010, 107, 18138–18142. [Google Scholar] [CrossRef] [Green Version]
- Biegel, E.; Schmidt, S.; González, J.M.; Müller, V. Biochemistry, evolution and physiological function of the Rnf complex, a novel ion-motive electron transport complex in prokaryotes. Cell. Mol. Life Sci. 2011, 68, 613–634. [Google Scholar] [CrossRef]
- Brandt, K.; Müller, V. Hybrid rotors in F1Fo ATP synthases: Subunit composition, distribution, and physiological significance. Biol. Chem. 2015, 396, 1031–1042. [Google Scholar] [CrossRef]
- Schuchmann, K.; Müller, V. A Bacterial Electron bifurcating Hydrogenase. J. Biol. Chem. 2012, 287, 31165–31171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poehlein, A.; Cebulla, M.; Ilg, M.M.; Bengelsdorf, F.R.; Schiel-Bengelsdorf, B.; Whited, G.; Andreesen, J.R.; Gottschalk, G.; Daniel, R.; Dürre, P. The Complete Genome Sequence of Clostridium aceticum: A missing link between Rnf- and cytochrome-containing autotrophic acetogens. mBio 2015, 6, e01168-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierce, E.; Xie, G.; Barabote, R.D.; Saunders, E.; Han, C.S.; Detter, J.C.; Richardson, P.; Brettin, T.S.; Das, A.; Ljungdahl, L.G.; et al. The complete genome sequence of Moorella thermoacetica (f. Clostridium thermoaceticum). Environ. Microbiol. 2008, 10, 2550–2573. [Google Scholar] [CrossRef] [Green Version]
- Braun, M.; Mayer, F.; Gottschalk, G. Clostridium aceticum (Wieringa), a microorganism producing acetic acid from molecular hydrogen and carbon dioxide. Arch. Microbiol. 1981, 128, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Hess, V.; Schuchmann, K.; Müller, V. The Ferredoxin:NAD+ Oxidoreductase (Rnf) from the Acetogen Acetobacterium woodii Requires Na+ and is Reversibly Coupled to the Membrane Potential. J. Biol. Chem. 2013, 288, 31496–31502. [Google Scholar] [CrossRef] [Green Version]
- Heinonen, J.E.; Lahti, R.J. A new and convenient colorimetric determination of inorganic orthophosphate and its application to the assay of inorganic pyrophosphatase. Anal. Biochem. 1981, 113, 313–317. [Google Scholar] [CrossRef]
- Wohlfarth, G.; Diekert, G. Thermodynamics of methylenetetrahydrofolate reduction to methyltetrahydrofolate and its implications for the energy metabolism of homoacetogenic bacteria. Arch. Microbiol. 1991, 155, 378–381. [Google Scholar] [CrossRef]
- Sheppard, C.A.; Trimmer, E.E.; Matthews, R.G. Purification and Properties of NADH-Dependent 5, 10-methylenetetrahydrofolate Reductase (MetF) from Escherichia coli. J. Bacteriol. 1999, 181, 718–725. [Google Scholar] [CrossRef] [Green Version]
- Bertsch, J.; Öppinger, C.; Hess, V.; Langer, J.D.; Müller, V. A Heterotrimeric NADH-Oxidizing Methylenetetrahydrofolate Reductase from the Acetogenic Bacterium Acetobacterium woodii. J. Bacteriol. 2015, 197, 1681–1689. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of proteine-dye-binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin-phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Wittig, I.; Carrozzo, R.; Santorelli, F.M.; Schägger, H. Functional assays in high-resolution clear native gels to quantify mitochondrial complexes in human biopsies and cell lines. Electrophoresis 2007, 28, 3811–3820. [Google Scholar] [CrossRef] [PubMed]
- Kuhns, M.; Trifunović, D.; Huber, H.; Müller, V. The Rnf complex is a Na+ coupled respiratory enzyme in a fermenting bacterium, Thermotoga maritima. Commun. Biol. 2020, 3, 1–10. [Google Scholar] [CrossRef]
- Müller, V.; Grüber, G. ATP synthases: Structure, function and evolution of unique energy converters. Cell. Mol. Life Sci. 2003, 60, 474–494. [Google Scholar] [CrossRef] [PubMed]
- Rahlfs, S.; Müller, V. Sequence of subunit a of the Na+-translocating F1FO-ATPase of Acetobacterium woodii: Proposal for residues involved in Na+ binding. FEBS Lett. 1999, 453, 35–40. [Google Scholar] [CrossRef] [Green Version]
- Rahlfs, S.; Müller, V. Sequence of subunit c of the Na+-translocating F1FO ATPase of Acetobacterium woodii: Proposal for determinants of Na+ specificity as revealed by sequence comparisons. FEBS Lett. 1997, 404, 269–271. [Google Scholar] [CrossRef] [Green Version]
- Aufurth, S.; Schägger, H.; Müller, V. Identification of subunits a, b, and c1 from Acetobacterium woodii Na+-F1FO-ATPase. Subunits c1, c2, and c3 constitute a mixed c-oligomer. J. Biol. Chem. 2000, 275, 33297–33301. [Google Scholar] [CrossRef] [Green Version]
- Müller, V. An Exceptional Variability in the Motor of Archaeal A1AO ATPases: From Multimeric to Monomeric Rotors Comprising 6-13 Ion Binding Sites. J. Bioenerg. Biomembr. 2004, 36, 115–125. [Google Scholar] [CrossRef]
- Rahlfs, S.; Aufurth, S.; Müller, V. The Na+-F1FO-ATPase operon from Acetobacterium woodii. Operon structure and presence of multiple copies of atpE which encode proteolipids of 8- and 18-kDa. J. Biol. Chem. 1999, 274, 33999–34004. [Google Scholar] [CrossRef] [Green Version]
- Spruth, M.; Reidlinger, J.; Müller, V. Sodium ion dependence of inhibition of the Na+-translocating F1FO-ATPase from Acetobacterium woodii. Probing the site(s) involved in ion transport. Biochim. Biophys. Acta (BBA) 1995, 1229, 96–102. [Google Scholar] [CrossRef] [Green Version]
- Schlegel, K.; Leone, V.; Faraldo-Gomez, J.D.; Müller, V. Promiscuous archaeal ATP synthase concurrently coupled to Na+ and H+ translocation. Proc. Natl. Acad. Sci. USA 2012, 109, 947–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thauer, R.K.; Jungermann, K.; Decker, K. Energy conservation in chemotrophic anaerobic bacteria. Bact. Rev. 1977, 41, 100–180. [Google Scholar] [CrossRef] [PubMed]
- Müller, V. Energy conservation in acetogenic bacteria. Appl. Environ. Microbiol. 2003, 69, 6345–6353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckel, W.; Thauer, R.K. Flavin-Based Electron bifurcation, a new mechanism of biological energy coupling. Chem. Rev. 2018, 118, 3862–3886. [Google Scholar] [CrossRef] [Green Version]
- Mock, J.; Wang, S.; Huang, H.; Kahnt, J.; Thauer, R.K. Evidence for a Hexaheteromeric Methylenetetrahydrofolate Reductase in Moorella thermoacetica. J. Bacteriol. 2014, 196, 3303–3314. [Google Scholar] [CrossRef] [Green Version]
- Kremp, F.; Roth, J.; Müller, V. The Sporomusa type Nfn is a novel type of electron-bifurcating transhydrogenase that links the redox pools in acetogenic bacteria. Sci. Rep. 2020, 10, 14872. [Google Scholar] [CrossRef]
- Wang, S.; Huang, H.; Kahnt, J.; Thauer, R.K. Clostridium acidurici electron-bifurcating formate dehydrogenase. Appl. Environ. Microbiol. 2013, 79, 6176–6179. [Google Scholar] [CrossRef] [Green Version]
- Mayer, F.; Müller, V. Adaptations of anaerobic archaea to life under extreme energy limitation. FEMS Microbiol. Rev. 2014, 38, 449–472. [Google Scholar] [CrossRef] [Green Version]
- Spahn, S.; Brandt, K.; Müller, V. A low phosphorylation potential in the acetogen Acetobacterium woodii reflects its lifestyle at the thermodynamic edge of life. Arch. Microbiol. 2015, 197, 745–751. [Google Scholar] [CrossRef]
- Müller, V.; Hess, V. The minimum biological energy quantum. Front. Microbiol. 2017, 8, 2019. [Google Scholar] [CrossRef]
- Matthies, D.; Zhou, W.; Klyszejko, A.L.; Anselmi, C.; Yildiz, O.; Brandt, K.; Müller, V.; Faraldo-Gomez, J.D.; Meier, T. High-resolution structure and mechanism of an F/V-hybrid rotor ring in a Na+-coupled ATP synthase. Nat. Commun. 2014, 5, 5286. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiechmann, A.; Müller, V. Energy Conservation in the Acetogenic Bacterium Clostridium aceticum. Microorganisms 2021, 9, 258. https://doi.org/10.3390/microorganisms9020258
Wiechmann A, Müller V. Energy Conservation in the Acetogenic Bacterium Clostridium aceticum. Microorganisms. 2021; 9(2):258. https://doi.org/10.3390/microorganisms9020258
Chicago/Turabian StyleWiechmann, Anja, and Volker Müller. 2021. "Energy Conservation in the Acetogenic Bacterium Clostridium aceticum" Microorganisms 9, no. 2: 258. https://doi.org/10.3390/microorganisms9020258
APA StyleWiechmann, A., & Müller, V. (2021). Energy Conservation in the Acetogenic Bacterium Clostridium aceticum. Microorganisms, 9(2), 258. https://doi.org/10.3390/microorganisms9020258