
Enhanced Viral Activity in the Surface Microlayer of the Arctic and Antarctic Oceans

 ,
,  ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Sampling Sites and Strategy
2.2. Physicochemical and Atmospheric Parameters
2.3. Viral and Microbial Abundance
2.4. Prokaryotic Heterotrophic Production
2.5. Viral Production and Rate of Lysed Prokaryotes
2.6. Data Analyses
3. Results
3.1. Environmental Parameters
3.2. Viral and Microbial Parameters
3.3. Viral and Microbial Abundances and Prokaryotic Heterotrophic Production
3.4. Viral Life Strategies (Lysis vs. Lysogeny)
3.5. Mortality Rates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liss, P.S.; Duce, R.A. The Sea Surface and Global Change, 1st ed.; Cambridge University Press: Cambridge, UK, 1997; 537p. [Google Scholar] [CrossRef]
- Cunliffe, M.; Engel, A.; Frka, S.; Gasparovic, B.; Guitart, C.; Murreell, J.C.; Salter, M.; Stolle, C. Sea surface microlayers: A unified physicochemical and biological perspective of the air-ocean interface. Prog. Oceanogr. 2013, 1109, 104–116. [Google Scholar] [CrossRef]
- Hardy, J.T. The sea-surface microlayer: Biology, chemistry and anthropogenic enrichment. Prog. Oceanogr. 1982, 11, 307–328. [Google Scholar] [CrossRef]
- Williams, P.M.; Carlucci, A.F.; Henrichs, S.M.; Van Vleet, E.S.; Horrigan, S.G.; Reid, F.M.H.; Roberston, K.J. Chemical and microbiological studies of sea-surface films in the southern gulf of California and off the west coast of Baja California. Mar. Chem. 1986, 19, 17–98. [Google Scholar] [CrossRef]
- Tovar-Sánchez, A.; González-Ortegón, E.; Duarte, C.M. Trace metal partitioning in the top meter of the ocean. Sci. Tot. Environ. 2019, 652, 907–914. [Google Scholar] [CrossRef]
- Kuznetsova, M.; Lee, C. Enhanced extracellular enzymatic peptide hydrolysis in the sea-surface microlayer. Mar. Chem. 2001, 73, 319–332. [Google Scholar] [CrossRef]
- Martinez-Varela, A.; Casas, G.; Piña, B.; Dachs, J.; Vila-Costa, M. Large Enrichment of Anthropogenic Organic Matter Degrading Bacteria in the Sea-Surface Microlayer at Coastal Livingston Island (Antarctica). Front. Microbiol. 2020, 11, 571983. [Google Scholar] [CrossRef]
- Rahlff, J. The virioneuston: A review on viral-bacterial associations at air-water interfaces. Viruses 2019, 11, 191. [Google Scholar] [CrossRef] [Green Version]
- Baylor, E.; Baylor, M.; Blanchard, D.C.; Syzdek, L.D.; Appel, C. Virus transfer from surf to wind. Science 1997, 198, 575–580. [Google Scholar] [CrossRef]
- Wilson, T.W.; Ladino, L.A.; Alpert, P.A.; Breckels, M.N.; Brooks, I.M.; Browse, J.; Burrows, S.M.; Carslaw, K.S.; Huffman, J.A.; Judd, C.; et al. A marine biogenic source of atmospheric ice-nucleating particles. Nature 2015, 525, 234–238. [Google Scholar] [CrossRef] [Green Version]
- Obernosterer, I.; Catala, P.; Reinthaler, T.; Herndl, G.J.; Lebaron, P. Enhanced heterotrophic activity in the surface microlayer of the Mediterranean Sea. Aquat. Microb. Ecol. 2005, 39, 293–302. [Google Scholar] [CrossRef]
- Rahlff, J.; Stolle, C.; Wurl, O. SISI: A new device for in situ incubations at the ocean surface. J. Mar. Sci. Eng. 2017, 5, 46. [Google Scholar] [CrossRef] [Green Version]
- Pradeep Ram, A.S.; Mari, X.; Brune, J.; Torreton, J.P.; Chu, V.T.; Raimbault, P.; Niggemann, J.; Sime-Ngando, T. Bacterial-viral interactions in the sea surface microlayer of a black carbon-dominated tropical coastal ecosystem (Halong Bay, Vietnam). Element. Sci. Anth. 2018, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Joux, F.; Agogué, H.; Obernosterer, I.; Dupuy, C.; Reinthaler, T.; Herndl, G.J.; Lebaron, P. Microbial community structure in the sea surface microlayer at two contrasting coastal sites in the northwestern Mediterranean Sea. Aquat. Microb. Ecol. 2006, 42, 91–104. [Google Scholar] [CrossRef] [Green Version]
- Tapper, M.A.; Hicks, R.E. Temperate viruses and lysogeny in Lake Superior bacterioplankton. Limnol. Oceanogr. 1998, 43, 95–103. [Google Scholar] [CrossRef]
- Bigg, E.K.; Leck, C.; Tranvik, L. Particulates of the surface microlayer of open water in the central Arctic Ocean in summer. Mar. Chem. 2004, 91, 131–141. [Google Scholar] [CrossRef]
- Fuhrman, J.A. Marine viruses and their biogeochemical and ecological effects. Nature 1999, 399, 541–548. [Google Scholar] [CrossRef]
- Sotomayor-Garcia, A.; Sala, M.M.; Ferrera, I.; Estrada, E.; Vázquez-Domínguez, E.; Emelianov, M.; Cortés, P.; Marrasé, C.; Ortega-Retuerta, E.; Nunes, S.; et al. Assessing viral abundance and community composition in four contrasting regions of the southernocean. Life 2020, 10, 107. [Google Scholar] [CrossRef]
- Boras, J.A.; Sala, M.M.; Arrieta, J.M.; Sà, E.L.; Felipe, J.; Agustí, S.; Duarte, C.M.; Vaqué, D. Effect of ice melting on bacterial carbon fluxes channeled by viruses and protists in the Arctic Ocean. Pol. Biol. 2010, 33, 1695–1707. [Google Scholar] [CrossRef]
- Vaqué, D.; Boras, J.A.; Torrent-Llagostera, F.; Agustí, S.; Arrieta, J.M.; Lara, E.; Castillo, Y.M.; Duarte, C.M.; Sala, M.M. Viruses and protists induced-mortality of prokaryotes around the Antarctic Peninsula during the austral summer. Front. Microbiol. 2017, 8, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sala, M.M.; Arrieta, J.M.; Boras, J.A.; Duarte, C.M.; Vaqué, D. The impact of ice melting on bacterioplankton in the Arctic Ocean. Pol. Biol. 2010, 33, 1683–1694. [Google Scholar] [CrossRef] [Green Version]
- Stortini, A.M.; Cincinelli, A.; Degli Innocenti, N.; Tovar-Sánchez, A.; Knulst, J. 1.12—Surface Microlayer. In Comprehensive Sampling and Sample Preparation; Pawliszyn, J., Ed.; Oxford Academic Press: Oxford, UK, 2012; pp. 223–246. [Google Scholar]
- Hansen, H.P.; Koroleff, F. Determination of nutrients. In Methods of Seawater Analysis, 3rd ed.; Grasshoff, K., Kremling, K., Ehrhardt, M., Eds.; Wiley: Hoboken, NJ, USA, 1999; Chapter 10; Available online: https://doi.org/10.1002/9783527613984.ch10 (accessed on 27 December 2007).
- Kerouel, R.; Aminot, A. Fluorometric determination of ammonia in sea and estuarine waters by direct segmented flow analysis. Mar. Chem. 1997, 57, 265–275. [Google Scholar] [CrossRef]
- Spyres, G.S.G.; Worsfold, P.J.; Miller, E.P.A.; Mimmo, M.; Miller, A.J. Determination of dissolved organic carbon in seawater using high-temperature catalytic oxidation techniques. Trends Anal. Chem. 2000, 19, 498–506. [Google Scholar] [CrossRef]
- Hsu, S.A.; Meindl, E.A.; Gilhousen, D.B. Determining the power-law wind-profile exponent under near-neutral stability conditions at sea. J. Appl. Met. 1994, 33, 757–765. [Google Scholar] [CrossRef] [Green Version]
- Brussaard, C.P.D. Optimisation of procedures for counting viruses by flow cytometry. Appl. Environ. Microbiol. 2004, 70, 1506–1513. [Google Scholar] [CrossRef] [Green Version]
- Gasol, J.M.; del Giorgio, P.A. Using flow cytometry for counting natural planktonic bacteria and understanding the structure of planktonic bacterial communities. Sci. Mar. 2000, 64, 197–224. [Google Scholar] [CrossRef] [Green Version]
- Evans, C.; Pearce, I.; Brussaard, C.P.D. Viral-mediated lysis of microbes and carbon release in the sub-Antarctic and Polar Frontal zones of the Australian Southern Ocean. Environ. Microbiol. 2009, 11, 2924–2934. [Google Scholar] [CrossRef]
- Del Giorgio, P.A.; Bird, D.F.; Prairie, Y.T.; Planas, D. Flow cytometric determination of bacterial abundance in lake plankton with the green nucleic acid stain SYTO 13. Limnol. Oceanogr. 1996, 41, 783–789. [Google Scholar] [CrossRef]
- Sieracki, M.E.; Johnson, P.W.; Sieburth, J.M. Detection, enumeration, and sizing of planktonic bacteria by image analyzed epifluorescence microscopy. Appl. Environ. Microbiol. 1985, 49, 799–810. [Google Scholar] [CrossRef] [Green Version]
- Kirchman, D.; K’nees, E.; Hodson, R. Leucine incorporation and its potential as a measure of protein synthesis by bacteria in natural aquatic systems. Appl. Environ. Microbiol. 1985, 43, 599–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.C.; Azam, F. A simple, economical method for measuring bacterial protein synthesis rates in seawater using 3H-leucine. Mar. Microb. Food Webs 1992, 6, 107–114. [Google Scholar]
- Kirchman, D.L. Incorporation of thymidine and leucine in the subarctic Pacific: Application to estimating bacterial production. Mar. Ecol. Prog. Ser. 1992, 82, 301–309. [Google Scholar] [CrossRef]
- Wilhelm, S.W.; Brigden, S.M.; Suttle, C.A. A dilution technique for the direct measurement of viral production: A comparison in stratified and tidally mixed coastal waters. Microb. Ecol. 2002, 43, 168–173. [Google Scholar] [CrossRef]
- Winget, D.M.; Williamson, K.E.; Helton, R.R.; Wommack, K.E. Tangential flow diafiltration: An improved technique for estimation of virioplankton production. Aquat. Microb. Ecol. 2005, 41, 221–232. [Google Scholar] [CrossRef] [Green Version]
- Weinbauer, M.G.; Suttle, C.A. Lysogeny and prophage induction in coastal and offshore bacterial communities. Aquat. Microb. Ecol. 1999, 18, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Weinbauer, M.G.; Winter, C.; Höfle, M.G. Reconsidering transmission electron microscopy-based estimates of viral infection of bacterioplankton using conversion factors derived from natural communities. Aquat. Microb. Ecol. 2002, 27, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.C.; Paul, J.H. Occurrence of lysogenic bacteria in marine microbial communities as determined by prophage induction. Mar. Ecol. Prog. Ser. 1996, 142, 27–38. [Google Scholar] [CrossRef]
- Steward, G.F.; Fandino, L.B.; Hollibaugh, J.T.; Whitledge, T.E.; Azam, F. Microbial biomass and viral infections of heterotrophic prokaryotes in the sub-surface layer of the central Arctic Ocean. Deep Sea Res. Part I Oceanogr. Res. Pap. 2007, 54, 1744–1757. [Google Scholar] [CrossRef]
- Bonilla-Findji, O.; Malits, A.; Lefèvre, D.; Rochelle-Newall, E.; Lemée, R.; Weinbauer, M.G.; Gattuso, J.-P. Viral effects on bacterial respiration, production and growth efficiency: Consistent trends in the Southern Ocean and the Mediterranean Sea. Deep Sea Res. II Top. Stud. Oceanogr. 2008, 55, 790–800. [Google Scholar] [CrossRef]
- Guixa-Boixereu, N.; Vaqué, D.; Gasol, J.M.; Sánchez-Cámara, J.; Pedrós-Alió, C. Viral distribution and activity in Antarctic waters. Deep Sea Res. II Top. Stud. Oceanogr. 2002, 49, 827–845. [Google Scholar] [CrossRef]
- Norland, S. The relationship between biomass and volume of bacteria. In Handbook of Methods in Aquatic Microbial Ecology; Kemp, P., Sherr, B.F., Sherr, E.B., Cole, J.J., Eds.; Lewis Publisher: Boca Raton, CA, USA, 1993; pp. 303–307. [Google Scholar]
- Simon, M.; Azam, F. Protein content and protein synthesis rates of planktonic marine bacteria. Mar. Ecol. Prog. Ser. 1989, 51, 201–213. [Google Scholar] [CrossRef]
- Vaqué, D.; Agustí, S.; Duarte, C.M. Response of bacterial grazing rates to experimental manipulation of an Antarctic coastal nanoflagellate community. Aquat. Microb. Ecol. 2004, 36, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Rahlff, J.; Ribas-Ribas, M.; Brown, S.M.; Hamizah Mustaffa, N.I.; Renz, J.; Peck, M.A.; Bird, K.; Cunliffe, M.; Melkonian, K.; Zappa, C.J. Blue pigmentation of neustonic copepods benefits exploitation of a prey-rich niche at the air-sea boundary. Sci. Rep. 2018, 8, 11510. [Google Scholar] [CrossRef]
- Danovaro, R.; Corinaldesi, C.; Dell’Anno, A.; Fuhrman, J.A.; Middelburg, J.J.; Noble, R.T.; Suttle, C.A. Marine viruses and global climate change. FEMS Microbiol. Rev. 2011, 35, 993–1034. [Google Scholar] [CrossRef]
- Nakajima, R.; Tsuchiya, K.; Nakatomi, N.; Yoshida, T.; Tada, Y.; Konno, F.; Toda, T.; Kuwahara, V.S.; Hamasaki, K.; Othman, B.H.R.; et al. Enrichment of microbial abundance in the sea-surface microlayer over a coral reef: Implications for biogeochemical cycles in reef ecosystems. Mar. Prog. Ser. 2013, 490, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Aller, J.Y.; Kuznetsova, M.R.; Jahns, C.J.; Kemp, P.F. The sea surface microlayer as a source of viral and bacterial enrichment in marine aerosols. J. Aerosol Sci. 2005, 36, 801–812. [Google Scholar] [CrossRef]
- Reche, I.; D’Orta, G.; Mladenov, N.; Winget, D.M.; Suttle, C.A. Deposition rates of viruses and bacteria above the atmospheric boundary layer. ISME J. 2018, 12, 1154–1162. [Google Scholar] [CrossRef] [Green Version]
- Reinthaler, T.; Sintes, E.; Herndl, G.J. Dissolved organic matter and bacterial production and respiration in the sea-surface microlayer of the open Atlantic and the western Mediterranean Sea. Limnol. Oceanogr. 2008, 53, 122–136. [Google Scholar] [CrossRef] [Green Version]
- Cunliffe, M.; Salter, M.; Mann, P.J.; Whiteley, A.; Upstill-Goddard, R.C.; Murrell, J.C. Dissolved organic carbon and bacterial populations in the gelatinous surface microlayer of a Norwegian fjord mesocosm. FEMS Microbiol. Lett. 2009, 299, 248–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agogué, H.; Casamayor, E.O.; Joux, F.; Obernosterer, I.; Dupuy, C.; Lantoine, F.; Catala, P.; Weinbauer, M.G.; Rheinthaler, T.; Herndl, G.J.; et al. Comparison of samplers for the biological characterization of the sea surface microlayer. Limnol. Oceanogr. Meth. 2004, 2, 213–225. [Google Scholar] [CrossRef]
- Cunliffe, M.; Harrison, E.; Salter, M.; Schäfe, H.; Upstill-Goddard, R.C.; Murrell, J.C. Comparison and validation of sampling strategies for the molecular microbial analysis of surface microlayers. Aquat. Microb. Ecol. 2009, 57, 69–77. [Google Scholar] [CrossRef] [Green Version]
- Wurl, O.; Wurl, E.; Miller, L.; Johnson, K.; Vagle, S. Formation and global distribution of sea-surface microlayers. Biogeosciences 2011, 8, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Engel, A.; Bange, H.W.; Cunliffe, M.; Burrows, S.M.; Friedrichs, G.; Galgani, L.; Herrmann, H.; Hertkorn, N.; Johnson, M.; Liss, P.S.; et al. The Ocean’s Vital Skin: Toward an Integrated Understanding of the Sea Surface Microlayer. Front. Mar. Sci. 2017, 4, 165. [Google Scholar] [CrossRef] [Green Version]
- Rahlff, J.; Stolle, C.; Giebel, H.A.; Brinkhoff, T.; Ribas-Ribas, M.; Hodapp, D.; Wurl, O. High wind speeds prevent formation of a distinct bacterioneuston community in the sea-surface microlayer. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [PubMed]
- Buma, A.G.J.; Helbing, E.W.; de Boer, M.K.; Villafañe, V.E. Patterns of DNA damage and photoinhibition in temperate South-Atlantic picophytoplankton exposed to solar ultraviolet radiation. J. Photochem. Photobiol. B 2001, 62, 9–18. [Google Scholar] [CrossRef]
- Hardy, J.T.; Apts, C.W. Photosynthetic carbon reduction: High rates in the sea-surface microlayer. Mar. Biol. 1989, 101, 411–417. [Google Scholar] [CrossRef]
- Suttle, C.A.; Chen, F. Mechanisms and Rates of Decay of Marine Viruses in Seawater. Appl. Environ. Microb. 1992, 58, 3721–3729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noble, R.T.; Fuhrman, J.A. Virus decay and its causes in coastal waters. Appl. Environ. Microbiol. 1997, 63, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, S.W.; Weinbauer, M.G.; Suttle, C.A.; Jeffrey, W.H. The role of sunlight in the removal and repair of viruses in the sea. Limnol. Oceanogr. 1998, 43, 586–592. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, S.W.; Weinbauer, M.G.; Suttle, C.A.; Pledger, R.J.; Mitchell, D.L. Measurements of DNA damage and photoreactivation imply that most viruses in marine surface waters are infective. Aquat. Microb. Ecol. 1998, 14, 215–222. [Google Scholar] [CrossRef]
- Weinbauer, M.G. Ecology of prokaryotic viruses. FEMS Microbiol. Rev. 2004, 28, 127–181. [Google Scholar] [CrossRef] [Green Version]
- Wilcox, R.M.; Fuhrman, J.A. Bacterial-viruses in coastal seawater—Lytic rather than lysogenic production. Mar. Ecol. Prog. Ser. 1994, 114, 35–45. [Google Scholar] [CrossRef]
- Knowles, B.; Silveira, C.B.; Bailey, B.A.; Barott, K.; Cantu, V.A.; Cobian-Guemes, A.G.; Coutinho, F.H.; Dinsdale, E.A.; Felts, B.; Furby, K.A.; et al. Lytic to temperate switching of viral communities. Nature 2016, 531, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Riemann, L.; Middelboe, M. Viral lysis of marine bacterioplankton: Implications for organic matter cycling and bacterial clonal composition. Ophelia 2002, 56, 57–68. [Google Scholar] [CrossRef]
- Suttle, C.A. Marine viruses—Major players in the global ecosystem. Nat. Rev. Microbiol. 2007, 5, 801–812. [Google Scholar] [CrossRef]
- Boras, J.; Vaqué, D.; Maynou, F.; Sà, E.L.; Weinbauer, M.G.; Sala, M.M. Factors shaping bacterial phylogenetic and functional diversity in coastal waters of the NW Mediterranean Sea. Est. Coast. Shelf Sci. 2015, 154, 102–110. [Google Scholar] [CrossRef]
- Zimmerman, A.E.; Howard- Varona, C.; Needham, D.M.; John, S.G.; Worden, A.Z.; Sullivan, M.B.; Waldbauer, J.R.; Coleman, M.L. Metabolic and biogeochemical consequences of viral infection in aquatic ecosystems. Nature Microbiol. Rev. 2020, 18, 21–34. [Google Scholar] [CrossRef]
- Sherr, E.B.; Sherr, B.F. Significance of predation by protists in aquatic microbial food webs. Ant. Van Leeuw. 2002, 81, 293–308. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
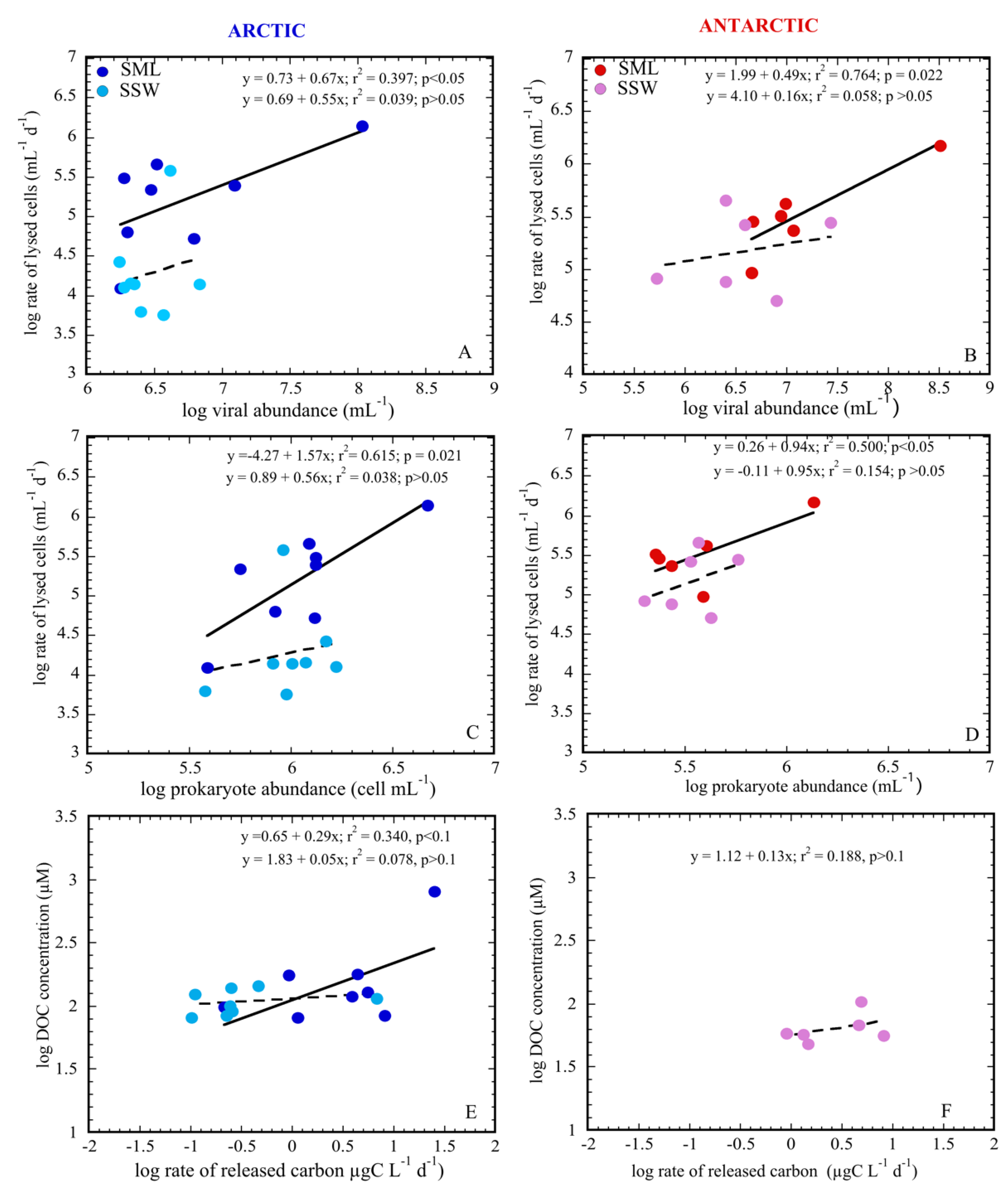{kind=link}
| VAR | AR | SML | SSW | EF (SML/SSW) | n | AN | SML | SSW | EF (SML/SSW) | n |
|---|---|---|---|---|---|---|---|---|---|---|
| Temp | All | 2.7 (−0.9–4.8) | 2.1 (−0.9–5.5) | 1.2 (0.2–2.9) | 19 | All | - | 1.1 (−0.4–3.2) | - | 18 |
| °C | T | 3.7 (2.8–4.4) | 2.6 (2.3–2.9) | 1.4 (1.2–1.6) | 5 | Br | - | 1.6 (−0.2–2.5) | - | 6 |
| S | 2.3 (−0.9–4.8) | 2.0 (−0.9–5.5) | 1.3 (0.2–2.9) | 14 | We | - | 0.1 (−0.4–0.9) | - | 6 | |
| Be | - | 1.7 (−1.1–3.2) | - | 6 | ||||||
| Salinity | All | 32.6 (31.2–34.4) | 32.7 (31.5–34.5) | 1.0 (1.0–1.0) | 19 | All | - | 33.9 (32.5–34.3) | - | 18 |
| T | 32.2 (31.3–33.4) | 32.2 (31.5–33.4) | 1.0 (1.0–1.1) | 5 | Br | - | 34.2 (34.1–34.2) | - | 6 | |
| S | 32.8 (31.2–34.4) | 32.8 (31.5–34.5) | 1.0 (1.0–1.0) | 14 | We | - | 33.9 (33.4–34.3) | - | 6 | |
| Be | - | 33.5(32.5–33.8) | - | 6 | ||||||
| DOC | All | 144.2 (71.5–808.8) | 98.3 (71.4–145.0) | 1.4 (0.8–5.9) | 18 | All | - | 72.3 (46.0–241.9) | - | 18 |
| µM | T | 91.2 (71.5–111.1) | 82.9 (71.4–92.5) | 1.1 (0.9–1.4) | 5 | Br | - | 58.6 (50.4–71.1) | - | 6 |
| S | 164.6 (79.2–808.8) | 105.3 (80.4–145.0) | 1.5 (0.8–5.9) | 13 | We | - | 60.5 (46.0–103.5) | - | 6 | |
| Be | - | 95.5 (47.9–241.9) | - | 6 | ||||||
| PO4 | All | 0.1 (0.0–0.9) | 0.1 (0.0–0.3) | 1.5 (0.5–6.0) | 18 | All | 1.2 (0.5–1.6) | 1.1 (0.3–1.6) | 1.2 (0.9–2.5) | 14 |
| µM | T | 0.1 (0.1–0.2) | 0.1 (0.0–0.2) | 1.3 (0.5–1.8) | 5 | Br | 1.5 (1.3–1.6) | 1.2 (0.6–1.4) | 1.6 (1.0–2.5) 1.1(0.9–1.4) 1.1(1.0–1.2) | 3 |
| S | 0.1 (0.0–0.9) | 0.1 (0.0–0.3) | 1.6 (0.5–6.0) | 13 | We | 0.9 (0.5–1,4) | 1.0 (0.3–1.4) | 1.1 (0.9–1.4) | 5 | |
| Be | 1.3 (1.1–1.6) | 1.2 (1.1–1.6) | 1.1 (1.0–1.2) | 6 | ||||||
| SiO4 | All | 3.0 (0.5–27.3) | 1.5 (0.6–3.6) | 1.7 (0.6–12.7) | 18 | All | 58.2 (40.2–68.5) | 58.3 (39.6–71.1) | 1.0 (1.0–1.3) | 14 |
| µM | T | 1.1 (0.5–2.3) | 1.1 (0.6–2.3) | 1.0 (0.9–1.1) | 5 | Br | 65.4 (63.5–68.5) | 65.2 (51.9–71.1) | 1.1 (1.0–1.3) | 3 |
| S | 3.7 (0.7–27.3) | 1.7 (0.8–3.6) | 1.9 (0.6–12.7) | 13 | We | 49.0 (40.2–63.3) | 50.5 (39.6–63.1) | 1.0 (1.0–1.1) | 5 | |
| Be | 62.4 (47.8–68.3) | 60.3 (48.6–66.3) | 1.0 (1.0–1.2) | 6 | ||||||
| NO3+NO2 | All | 0.7 (0.1–3.2) | 0.9 (0.0–5.3) | 1.3 (0.6–5.0) | 18 | All | 15.0 (6.6–21.0) | 15.6 (6.6–21.1) | 1.1 (0.7–1.7) | 13 |
| µM | T | 0.4 (0.2–0.5) | 0.3 (0.0–0.6) | 1.9 (0.9–5.0) | 5 | Br | 18.3 (18.1–20.1) | 17.0 (7.4–20.9) | 1.6 (0.9–1.7) | 3 |
| S | 0.8 (0.1–3.16) | 1.1 (0.1–5.3) | 1.0 (0.6–1.2) | 13 | We | 13.5 (6.6–21.0) | 15.0 (6.6–21.0) | 1.0 (0.9–1.1) | 4 | |
| Be | 16.7 (10.0–20.7) | 17.2 (15.3–21.1) | 1.0 (0.7–1.2) | 6 | ||||||
| NH4 | All | 0.5 (0.2–1.2) | 0.3 (0.1–0.7) | 1.7 (0.8–3.8) | 19 | All | 1.3 (0.1–4.6) | 1.1 (0.1–3.1) | 1.1 (0.3–2.2) | 6 |
| µM | T | 0.5 (0.2–1.2) | 0.4 (0.2–0.7) | 1.2 (0.9–1.9) | 5 | Br | 0.1 | 0.3 (0.2–0.3) | 0.3 | 1 |
| S | 0.4 (0.2–0.9) | 0.3 (0.1–0.7) | 1.8 (0.8–3.8) | 14 | We | 0.3 (0.1–0.5) | 1.2 (0.1–3.1) | 1.1 (1.1–1.3) | 2 | |
| Be | 2.8 (1.6–4.6) | 1.8 (1.4–2.1) | 1.1 (1.1–2.2) | 3 | ||||||
| VAR | AR | Atmosphere | n | AN | Atmosphere | n | ||||
| Wind | All | 5.0 (1.2–7.9) | 19 | All | 5.5 (2.6–9.9) | 19 | ||||
| m s−1 | T | 4.3 (1.2–6.8) | 5 | Br | 5.7 (3.3–8.5) | 6 | ||||
| S | 5.2 (1.8–7.9) | 14 | We | 5.9 (3.5–9.9) | 7 | |||||
| Be | 4.7 (2.6–7.9) | 6 | ||||||||
| UV | All | 5.4 (1.6–8.9) | 13 | All | 2.5 (0.7–5.2) | 11 | ||||
| index | T | - | - | Br | 2.1 (1.4–3.9) | 4 | ||||
| S | 5.4 (1.6–8.9) | 13 | We | 3.5 (2.1–5.2) | 4 | |||||
| Be | 1.6 (0.7–2.4) | 3 |
| VAR | AR | SML | SSW | EF | n | AN | SML | SSW | EF | n |
|---|---|---|---|---|---|---|---|---|---|---|
| VA | All | 9.6 (1.3–108.3) | 3.3 (1.2–6.8) | 2.2 (0.5–15.9) | 19 | All | 25.7 (0.4–321.0) | 8.1 (1.3–42.9) | 2.7 (0.3–11.7) | 19 |
| 106 mL−1 | T | 4.8 (1.5–11.7) | 3.5 (1.2–6.7) | 1.7 (0.5–3.4) | 5 | Br | 6.4 (3.6–11.5) | 4.3 (2.5–7.9) | 1.8 (0.7–4.5) | 6 |
| S | 11.6 (1.3–108.3) | 3.2 (1.6–6.8) | 2.4 (0.7–15.9) | 14 | We | 49.9 (0.4–321.0) | 5.6 (1.5–27.4) | 4.1 (0.3–11.7) | 7 | |
| Be | 16.9 (3.0–22.5) | 14.5 (1.3–42.9) | 1.8 (0.4–4.8) | 6 | ||||||
| PA | All | 1.0 (0.2–4.7) | 0.8 (0.4–1.9) | 1.2 (0.4–4.7) | 19 | All | 0.5 (0.2–2.1) | 0.4 (0.2–0.8) | 1.3 (0.7–2.8) | 19 |
| 106 mL−1 | T | 1.0 (0.2–1.5) | 0.8 (0.4–1.9) | 1.1 (0.4–1.5) | 5 | Br | 0.4 (0.3–0.8) | 0.4 (0.3–0.8) | 1.0 (1.0–1.1) | 6 |
| S | 1.1 (0.4–4.7) | 0.8 (0.4–1.7) | 1.3 (0.5–4.7) | 14 | We | 0.5 (0.2–1.4) | 0.4 (0.2–0.6) | 1.4 (0.8–2.4) | 7 | |
| Be | 0.7 (0.2–2.1) | 0.4 (0.2–0.7) | 1.4 (0.7–2.8) | 6 | ||||||
| VPR | All | 7.1 (1.2–23.0) | 4.8 (1.1–9.1) | 1.7 (0.0–6.8) | 19 | All | 34.1 (1.6–136.5) | 22.8 (2.7–181.1) | 2.1 (0.2–7.3) | 19 |
| T | 8.5 (1.2–19.5) | 4.9 (2.9–9.1) | 2.2 (0.0–6.8) | 5 | Br | 18.1 (6.4–42.6) | 11.1 (5.5–18.7) | 1.8 (0.6–4.5) | 6 | |
| S | 6.7 (1.4–23.0) | 4.8 (1.1–8.5) | 1.5 (0.7–3.4) | 14 | We | 43.9 (1.6–236.2) | 13.0 (2.7–47.7) | 2.9 (0.2–7.3) | 7 | |
| Be | 38.5 (5.3–136.5) | 46.0 (4.2–181.1) | 1.6 (0.4–4.1) | 6 | ||||||
| PHP | All | 3.8 (0.4–18.6) | 1.3 (0.3–5.1) | 1.5 (0.6–4.3) | 19 | All | 3.9 (0.6–18.7) | 3.9 (0.8–21.4) | 1.2 (0.2–4.7) | 19 |
| µg C L−1 d−1 | T | 2.0 (0.5–3.9) | 1.9 (0.3–5.1) | 1.2 (0.8–1.6) | 5 | Br | 2.1 (0.6–2.7) | 5.1(0.8–21.4) | 1.0 (0.2–2.2) | 6 |
| S | 3.4 (0.4–18.6) | 2.1 (0.4–4.3) | 1.6 (0.6–4.3) | 14 | We | 4.2 (1.0–10.9) | 3.1 (1.6–9.5) | 1.1 (0.4–2.2) | 7 | |
| Be | 5.3 (1.6–18.7) | 3.1 (2.2–4.0) | 1.5 (0.7–4.7) | 6 | ||||||
| VPL | All | 0.07 (nd−0.1) | 0.06 (nd−0.1) | 1.9 (nd−4.4) | 9 | All | 8.4 (2.2–18.9) | 1.9 (0.3–4.4) | 5.4 (1.0–11.2) | 6 |
| 107 mL−1 d−1 | T | - | - | - | - | Br | 9.3 (4.2–18.9) | 2.3 (0.7–4.4) | 6.4 (1.0–11.2) | 3 |
| S | 0.07 (nd−0.1) | 0.06 (nd−0.1) | 1.9 (nd−4.4) | 9 | We | 10.4 (2.2–18.7) | 2.3 (1.0–3.5) | 3.7 (2.1–5.4) | 2 | |
| Be | 1.7 | 0.3 | 5.4 | 1 | ||||||
| VMM | All | 20.2 (nd−38.6) | 7.3 (nd−29.6) | 6.9 (nd−17.4) | 9 | All | 97.4 (24.1–141.4) | 55.0 (11.9–123.5) | 3.7 (0.2–8.7) | 6 |
| % | T | - | - | - | Br | 71.2 (24.1–103.5) | 54.4 (11.9–123.5) | 4.0 (0.2–8.7) | 3 | |
| S | 20.2 (nd−38.6) | 7.3 (nd−29.6) | 6.9 (nd−17.4) | 9 | We | 114.7 (108.5–120.9) | 44.5 (41.2–47.8) | 4.2 (2.9–5.4) | 2 | |
| Be | 141.4 | 78.2 | 1.8 | 1 | ||||||
| Lysogeny | All | 6.8 (nd−13.5) | 0.9 (nd−2.2) | nd | 9 | All | 40.3 (nd−40,3) | 16.2 (nd−36.7) | nd | 6 |
| % | T | - | - | - | - | Br | nd | 6.0 (nd−10.2) | nd | 3 |
| S | 6.8 (nd−13.5) | 0.9 (nd−2.2) | nd | 9 | We | nd | 36.7 (nd−36.7) | nd | 2 | |
| Be | 40.3 | nd | nd | 1 | ||||||
| RLC | All | 0.3 (nd−1.4) | 0.1(nd−0.4) | 27.2 (nd−99.4) | 9 | All | 0.7 (0.1–1.5) | 0.2 (0.1–0.5) | 7.2 (0.2–8.3) | 6 |
| 106 mL−1d−1 | T | - | - | - | - | Br | 0.3 (0.1–0.4) | 0.2 (0.1–0.5) | 3.4 (0.2–8.3) | 3 |
| S | 0.3 (nd−1.4) | 0.1 (nd−0.4) | 27.2 (nd−99.4) | 9 | We | 0.9 (0.3–1.5) | 0.2 (0.1–0.3) | 4.4 (3.5–5.4) | 2 | |
| Be | 0.3 | 0.1 | 1.2 | 1 | ||||||
| RLC_C | All | 6.1 (nd−24.9) | 1.1 (nd−6.8) | 27.2 (nd−99.4) | 9 | All | 11.6 (1.7–26.7) | 3.6 (0.9–8.1) | 7.2 (0.2–8.3) | 6 |
| µg C L−1d−1 | T | - | - | - | - | Br | 10.8 (1.7–26.7) | 3.5 (0.9–8.1) | 3.4 (0.2–8.3) | 3 |
| S | 6.1 (nd−24.9) | 1.1 (nd−6.8) | 27.2 (nd−99.4) | 9 | We | 15.8 (5.1–26.4) | 3.2 (1.5–4.9) | 4.4 (3.5–5.4) | 2 | |
| Be | 5.7 | 4.7 | 1.2 | 1 | ||||||
| HNF | All | 2.6 (0.2–7.0) | 2.5 (0.2–8.9) | 1.6 (0.2–4.1) | 19 | All | 1.4 (0.5–2.4) | 1.5 (0.3–2.7) | 1.3 (0.4–3.1) | 6 |
| 103 mL−1 | T | 4.3 (1.3–6.7) | 3.9 (1.53–8.9) | 1.4 (0.8–3.1) | 5 | Br | 1.5 (1.0–2.4) | 1.5 (0.3–2.5) | 1.6 (0.4–3.1) | 3 |
| S | 1.8 (0.2–7.0) | 1.7 (0.2–6.1) | 1.6 (0.2–4.1) | 14 | We | 1.9 | 1.9 (1.0–2.7) | 0.7 | 2 | |
| Be | 0.5 | 1.0 | 0.5 | 1 | ||||||
| %HNF < 5 | All | 80.5 (47.2–100) | 83.5 (39.5–100) | 1.0 (0.7–1.6) | 19 | All | 82.4 (70.8–91.6) | 67.1 (28.8–82.6) | 1.5 (1.0–3.0) | 6 |
| T | 61.9 (42.4–88.3) | 75.7 (39.5–98.9) | 0.9 (0.7–1.6) | 5 | Br | 83.2 (76.0–86.6) | 59.3 (28.8–82.6) | 1.7 (1.1–3.0) | 3 | |
| S | 89.8 (47.2–100) | 87.4 (63.7–100) | 1.0 (0.7–1.3) | 14 | We | 70.8 | 72.2 (70.7–73.7) | 1.0 | 2 | |
| Be | 91.6 | 80.1 | 1.1 | 1 | ||||||
| PNF | All | 9.2 (0.2–44.3) | 10.1 (1.7–19.0) | 0.8 (0.1–2.8) | 19 | All | 3.5 (0.9–5.1) | 2.0 (0.5–3.1) | 3.1 (0.8–9.8) | 6 |
| 103 mL−1 | T | 2.8 (0.2–7.5) | 4.8 (1.7–7.9) | 0.6 (0.1–1.0) | 5 | Br | 3.8 (2.4–5.0) | 2.1 (0.5–3.1) | 4.0 (0.8–9.8) | 3 |
| S | 12.4 (0.5–44.3) | 12.2 (5.7–19.0) | 0.9 (0.1–2.8) | 14 | We | 5.1 | 2.2 (1.8–2.5) | 2.8 | 2 | |
| Be | 0.9 | 1.0 | 0.9 | 1 |
| Variables | r | Arctic (p) | n | r | Antarctic (p) | n |
|---|---|---|---|---|---|---|
| VA | 0.607 | <0.006 | 19 | 0.689 | <0.001 | 19 |
| PA | 0.782 | <0.001 | 19 | 0.823 | <0.0001 | 19 |
| VPR | 0.423 | <0.05 | 19 | 0.586 | <0.008 | 19 |
| PHP | 0.805 | <0.0001 | 19 | 0.625 | <0.0042 | 18 |
| VPL | 0.486 | >0.05 | 8 | 0.621 | >0.05 | 6 |
| RLC | 0.221 | >0.05 | 8 | −0.011 | >0.05 | 6 |
| %VMM | −0.139 | >0.05 | 8 | −0.795 | <0.05 | 6 |
| HNF | 0.812 | <0.0001 | 15 | 0.406 | >0.05 | 5 |
| PNF | 0.804 | <0.0001 | 14 | 0.018 | >0.05 | 5 |
| T | 0.929 | <0.0001 | 19 | - | - | - |
| Sal | 0.994 | <0.0001 | 19 | - | - | - |
| DOC | 0.582 | <0.01 | 18 | - | - | - |
| PO4 | 0.718 | <0.001 | 17 | 0.771 | <0.001 | 13 |
| SiO4 | 0.702 | <0.001 | 18 | 0.892 | <0.0001 | 13 |
| NO3+NO2 | 0.949 | <0.0001 | 18 | 0.605 | <0.03 | 12 |
| NH4 | 0.629 | <0.001 | 19 | 0.921 | <0.01 | 5 |
| Variable | Water/Air | df | F | p | Value Comparison |
|---|---|---|---|---|---|
| Temperature | SSW | 36 | 3.72 | 0.05 | * |
| Salinity | SSW | 36 | 21.94 | <0.0001 | * |
| DOC | SSW | 34 | 13.2 | <0.0009 | * |
| EF | 28 | 0.780 | 0.345 | ns | |
| PO4 | SML | 29 | 91.11 | <0.00001 | * |
| SSW | 34 | 121.36 | <0.00001 | * | |
| EF | 29 | 0.66 | 0.42 | ns | |
| SiO4 | SML | 29 | 195.75 | <0.00001 | * |
| SSW | 34 | 777.68 | <0.00001 | * | |
| EF | 28 | 0.15 | 0.70 | ns | |
| NO3+NO2 | SML | 28 | 112.60 | <0.00001 | * |
| SSW | 33 | 114.91 | <0.00001 | * | |
| EF | 24 | 1.69 | 0.206 | ns | |
| NH4 | SML | 24 | 1.84 | 0.190 | ns |
| SSW | 28 | 70.58 | 0.003 | * | |
| Wind | air | 36 | 0.70 | 0.391 | ns |
| UVB | air | 21 | 12.51 | 0.0021 | * |
| EF | 37 | 0.16 | 0.693 | ns | |
| VA | SML | 37 | 1.81 | 0.186 | ns |
| SSW | 37 | 2.05 | 0.161 | ns | |
| EF | 37 | 0.67 | 0.418 | ns | |
| VPR | SML | 37 | 10.46 | 0.003 | * |
| SSW | 37 | 14.52 | 0.0005 | * | |
| EF | 37 | 0.06 | 0.805 | ns | |
| PA | SML | 37 | 5.49 | 0.025 | * |
| SSW | 37 | 18.21 | 0.0001 | * | |
| EF | 37 | 1.09 | 0.301 | ns | |
| PHP | SML | 37 | 0.97 | 0.331 | ns |
| SSW | 37 | 5.80 | 0.021 | * | |
| EF | 13 | 5.98 | 0.031 | * | |
| VPL | SML | 13 | 95.44 | <0.00001 | * |
| SSW | 13 | 41.47 | <0.0001 | * | |
| EF | 13 | 1.46 | 0.251 | ns | |
| RLC | SML | 13 | 1.69 | 0.219 | ns |
| SSW | 13 | 11.08 | 0.006 | * | |
| EF | 13 | 0.83 | 0.380 | ns | |
| %VMM | SML | 13 | 14.49 | 0.003 | * |
| SSW | 13 | 21.47 | 0.0006 | * | |
| EF | 19 | 0.37 | 0.552 | ns | |
| HNF | SML | 19 | 0.27 | 0.609 | ns |
| SSW | 19 | 0.003 | 0.955 | ns | |
| EF | 18 | 5.22 | 0.036 | * | |
| PNF | SML | 19 | 0.23 | 0.636 | ns |
| SSW | 19 | 20.27 | 0.0003 | * |
| Site | Wind Speed | n | EF–VA | n | EF–PA | n | EF–RLC | n | EF–DOC | n | EF–PO4 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| AR | Range: 1.2–7.9 | ||||||||||
| ≤5 m s−1 | 9 | 3.0 ± 1.5 | 9 | 1.4 ± 0.4 | 4 | 47.9 ± 21.2 | 9 | 1.6 ± 0.5 | 8 | 1.4 ± 0.2 | |
| 5–6 m s−1 | 4 | 2.4 ± 0.4 | 4 | 1.2 ± 0.1 | 1 | 3.6 ± 0.0 | 3 | 1.3 ± 0.3 | 4 | 1.3 ± 0.1 | |
| >6 m s−1 | 6 | 0.9 ± 0.1 | 6 | 0.9 ± 0.1 | 3 | 7.5 ± 3.4 | 6 | 0.9 ± 0.1 | 5 | 0.9 ± 0.3 | |
| AN | Range: 2.6–9.9 | ||||||||||
| ≤5 m s−1 | 7 | 2.6 ± 1.4 | 7 | 1.6 ± 0.2 | 1 | 6.2 ± 0.00 | - | 7 | 1.3 ± 0.2 | ||
| 5–6 m s−1 | 5 | 3.7 ± 1.3 | 5 | 1.1 ± 0.04 | 3 | 5.7 ± 0.6 | - | 1 | 1.0 ± 0.0 | ||
| >6 m s−1 | 7 | 1.9 ± 0.3 | 9 | 1.1 ± 0.1 | 2 | 5.2 ± 0.2 | - | 6 | 1.0 ± 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaqué, D.; Boras, J.A.; Arrieta, J.M.; Agustí, S.; Duarte, C.M.; Sala, M.M. Enhanced Viral Activity in the Surface Microlayer of the Arctic and Antarctic Oceans. Microorganisms 2021, 9, 317. https://doi.org/10.3390/microorganisms9020317
Vaqué D, Boras JA, Arrieta JM, Agustí S, Duarte CM, Sala MM. Enhanced Viral Activity in the Surface Microlayer of the Arctic and Antarctic Oceans. Microorganisms. 2021; 9(2):317. https://doi.org/10.3390/microorganisms9020317
Chicago/Turabian StyleVaqué, Dolors, Julia A. Boras, Jesús Maria Arrieta, Susana Agustí, Carlos M. Duarte, and Maria Montserrat Sala. 2021. "Enhanced Viral Activity in the Surface Microlayer of the Arctic and Antarctic Oceans" Microorganisms 9, no. 2: 317. https://doi.org/10.3390/microorganisms9020317
APA StyleVaqué, D., Boras, J. A., Arrieta, J. M., Agustí, S., Duarte, C. M., & Sala, M. M. (2021). Enhanced Viral Activity in the Surface Microlayer of the Arctic and Antarctic Oceans. Microorganisms, 9(2), 317. https://doi.org/10.3390/microorganisms9020317