
Impact of the Histidine-Containing Phosphocarrier Protein HPr on Carbon Metabolism and Virulence in Staphylococcus aureus

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Plasmids
2.2. Bacterial Growth Conditions
2.3. Mutant Construction
2.4. RNA Isolation and Purification, cDNA Synthesis and qRT-PCR
2.5. Measurement of pH, Glucose, Acetate, and Ammonium in Culture Supernatants
2.6. Biofilm Assays
2.7. Primary Attachment Assay on Polystyrene
2.8. Animal Models
2.9. Statistical Analyses
3. Results and Discussion
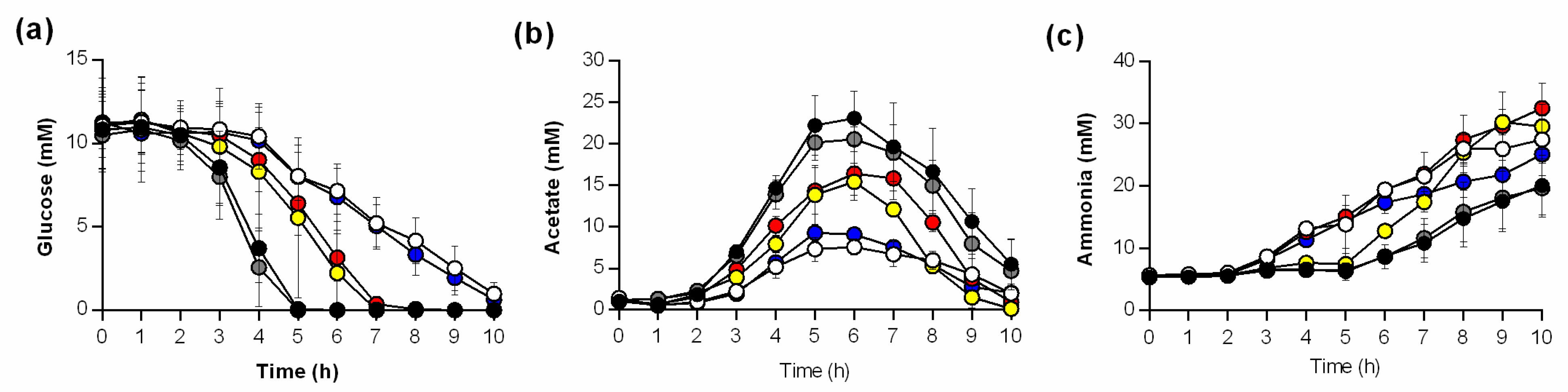
3.1. Growth, pH Characteristics, and Metabolite Profiles Differ between ptsH and ccpA Mutants
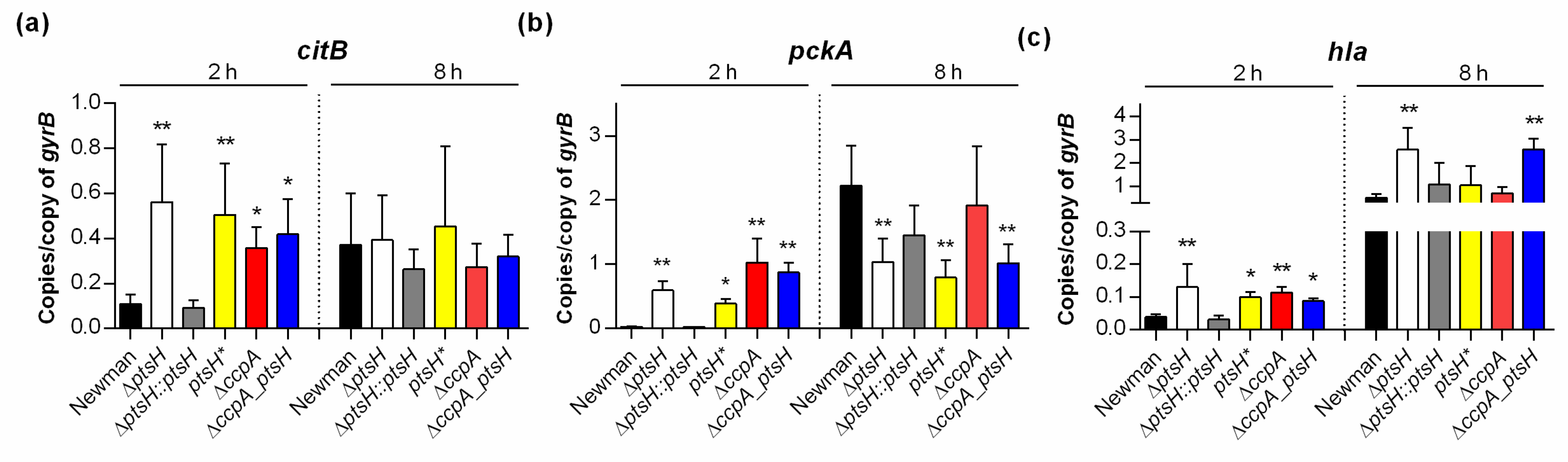
3.2. Inactivation of ptsH and/or ccpA Alters Transcription of TCA Cycle and Virulence Factor Genes
3.3. Impact of ptsH Deletion on Biofilm Formation of S. aureus SA113
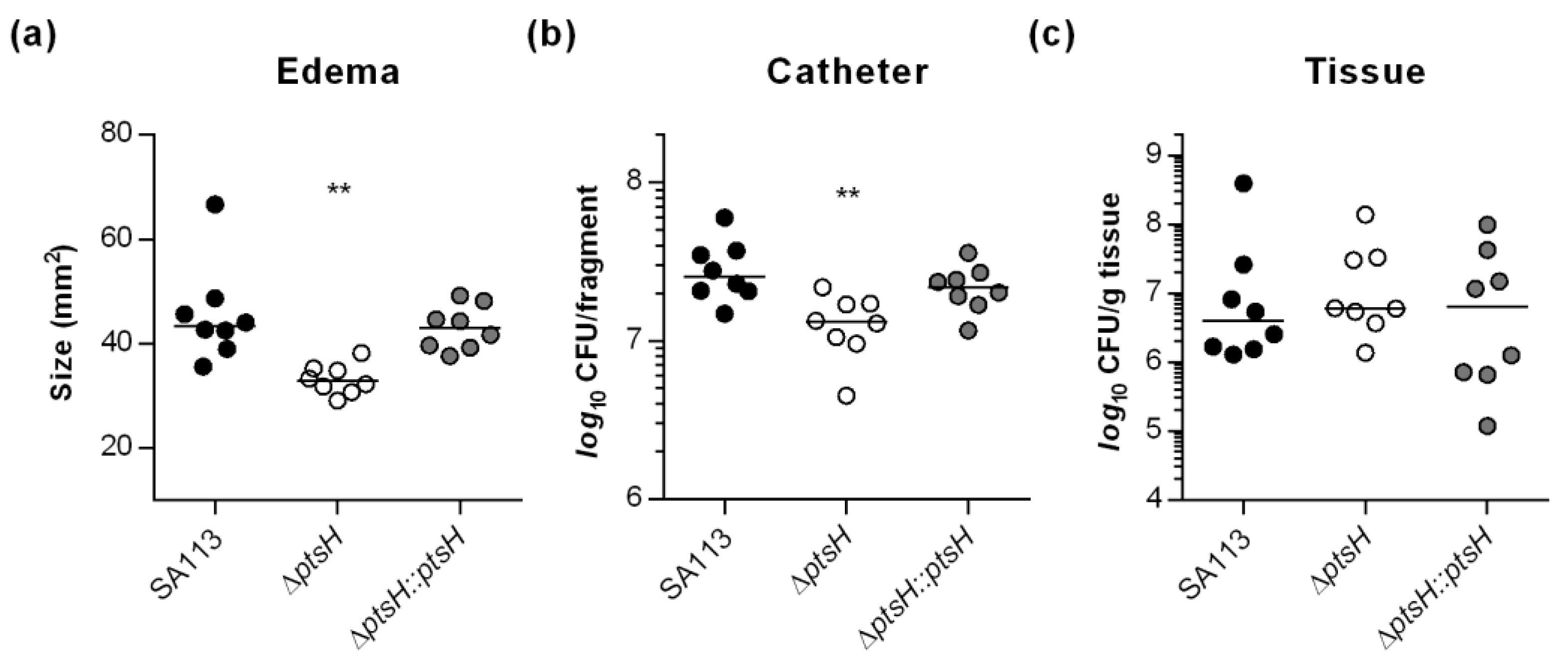
3.4. HPr Contributes to Infectivity and Biofilm Formation of S. aureus SA113 in a Murine Foreign Body Infection Model
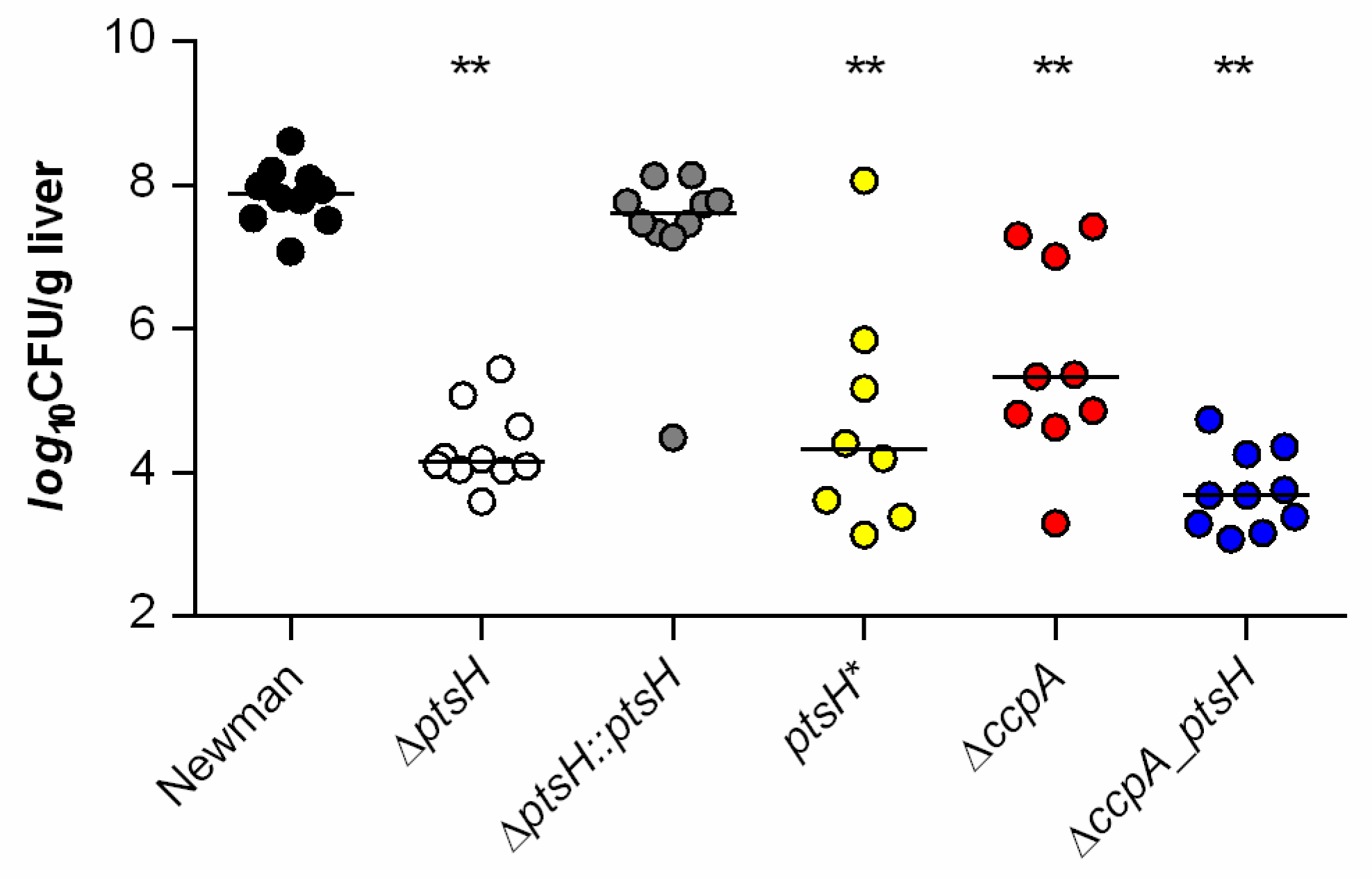
3.5. HPr and CcpA Are Both Required for Full Infectivity of S. aureus in a Murine Liver Abscess Model
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deutscher, J. The mechanisms of carbon catabolite repression in bacteria. Curr. Opin. Microbiol. 2008, 11, 87–93. [Google Scholar] [CrossRef]
- Görke, B.; Stülke, J. Carbon catabolite repression in bacteria: Many ways to make the most out of nutrients. Nat. Rev. Microbiol. 2008, 6, 613–624. [Google Scholar] [CrossRef]
- Richardson, A.R. Virulence and Metabolism. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Henkin, T.M.; Grundy, F.J.; Nicholson, W.L.; Chambliss, G.H. Catabolite repression of α amylase gene expression in Bacillus subtilis involves a trans-acting gene product homologous to the Escherichia coli lacl and galR repressors. Mol. Microbiol. 1991, 5, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Weickert, M.J.; Chambliss, G.H. Site-directed mutagenesis of a catabolite repression operator sequence in Bacillus subtilis. Proc. Natl. Acad. Sci. USA 1990, 87, 6238–6242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Sun, F.; Cho, H.; Yelavarthi, V.; Sohn, C.; He, C.; Schneewind, O.; Bae, T. CcpA mediates proline auxotrophy and is required for Staphylococcus aureus pathogenesis. J. Bacteriol. 2010, 192, 3883–3892. [Google Scholar] [CrossRef] [Green Version]
- Deutscher, J.; Saier, M.H. ATP-dependent protein kinase-catalyzed phosphorylation of a seryl residue in HPr, a phosphate carrier protein of the phosphotransferase system in Streptococcus pyogenes. Proc. Natl. Acad. Sci. USA 1983, 80, 6790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumacher, M.A.; Allen, G.S.; Diel, M.; Seidel, G.; Hillen, W.; Brennan, R.G. Structural Basis for Allosteric Control of the Transcription Regulator CcpA by the Phosphoprotein HPr-Ser46-P. Cell 2004, 118, 731–741. [Google Scholar] [CrossRef] [Green Version]
- Leiba, J.; Hartmann, T.; Cluzel, M.E.; Cohen-Gonsaud, M.; Delolme, F.; Bischoff, M.; Molle, V. A novel mode of regulation of the Staphylococcus aureus catabolite control protein A (CcpA) mediated by Stk1 protein phosphorylation. J. Biol. Chem. 2012, 287, 43607–43619. [Google Scholar] [CrossRef] [Green Version]
- Ramstrom, H.; Sanglier, S.; Leize-Wagner, E.; Philippe, C.; Van Dorsselaer, A.; Haiech, J. Properties and regulation of the bifunctional enzyme HPr kinase/phosphatase in Bacillus subtilis. J. Biol. Chem. 2003, 278, 1174–1185. [Google Scholar] [CrossRef] [Green Version]
- Hengstenberg, W.; Penberthy, W.K.; Hill, K.L.; Morse, M.L. Phosphotransferase system of Staphylococcus aureus: Its requirement for the accumulation and metabolism of galactosides. J. Bacteriol. 1969, 99, 383–388. [Google Scholar] [CrossRef] [Green Version]
- Postma, P.W.; Lengeler, J.W.; Jacobson, G.R. Phosphoenolpyruvate:carbohydrate phosphotransferase systems of bacteria. Microbiol. Rev. 1993, 57, 543. [Google Scholar] [CrossRef] [PubMed]
- Deutscher, J.; Küster, E.; Bergstedt, U.; Charrier, V.; Hillen, W. Protein kinase-dependent HPr/CcpA interaction links glycolytic activity to carbon catabolite repression in Gram-positive bacteria. Mol. Microbiol. 1995, 15, 1049–1053. [Google Scholar] [CrossRef] [PubMed]
- Deutscher, J.; Francke, C.; Postma, P.W. How Phosphotransferase System-Related Protein Phosphorylation Regulates Carbohydrate Metabolism in Bacteria. Microbiol. Mol. Biol. Rev. 2006, 70, 939. [Google Scholar] [CrossRef] [Green Version]
- Seidl, K.; Stucki, M.; Ruegg, M.; Goerke, C.; Wolz, C.; Harris, L.; Berger-Bächi, B.; Bischoff, M. Staphylococcus aureus CcpA affects virulence determinant production and antibiotic resistance. Antimicrob. Agents Chemother. 2006, 50, 1183–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidl, K.; Müller, S.; François, P.; Kriebitzsch, C.; Schrenzel, J.; Engelmann, S.; Bischoff, M.; Berger-Bächi, B. Effect of a glucose impulse on the CcpA regulon in Staphylococcus aureus. BMC Microbiol. 2009, 9, 95. [Google Scholar] [CrossRef] [Green Version]
- Reed, J.M.; Olson, S.; Brees, D.F.; Griffin, C.E.; Grove, R.A.; Davis, P.J.; Kachman, S.D.; Adamec, J.; Somerville, G.A. Coordinated regulation of transcription by CcpA and the Staphylococcus aureus two-component system HptRS. PLoS ONE 2018, 13, e0207161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidl, K.; Bischoff, M.; Berger-Bächi, B. CcpA mediates the catabolite repression of tst in Staphylococcus aureus. Infect. Immun. 2008, 76, 5093–5099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidl, K.; Goerke, C.; Wolz, C.; Mack, D.; Berger-Bächi, B.; Bischoff, M. Staphylococcus aureus CcpA affects biofilm formation. Infect. Immun. 2008, 76, 2044–2050. [Google Scholar] [CrossRef] [Green Version]
- Halsey, C.R.; Lei, S.; Wax, J.K.; Lehman, M.K.; Nuxoll, A.S.; Steinke, L.; Sadykov, M.; Powers, R.; Fey, P.D. Amino Acid Catabolism in Staphylococcus aureus and the Function of Carbon Catabolite Repression. mBio 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Nuxoll, A.S.; Halouska, S.M.; Sadykov, M.R.; Hanke, M.L.; Bayles, K.W.; Kielian, T.; Powers, R.; Fey, P.D. CcpA regulates arginine biosynthesis in Staphylococcus aureus through repression of proline catabolism. PLoS Pathog. 2012, 8, e1003033. [Google Scholar] [CrossRef] [PubMed]
- Cramton, S.E.; Gerke, C.; Schnell, N.F.; Nichols, W.W.; Götz, F. The intercellular adhesion (ica) locus is present in Staphylococcus aureus and is required for biofilm formation. Infect. Immun. 1999, 67, 5427–5433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, E.E.; Rice, K.C.; Boles, B.R.; Endres, J.L.; Ranjit, D.; Chandramohan, L.; Tsang, L.H.; Smeltzer, M.S.; Horswill, A.R.; Bayles, K.W. Modulation of eDNA release and degradation affects Staphylococcus aureus biofilm maturation. PLoS ONE 2009, 4, e5822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischoff, M.; Wonnenberg, B.; Nippe, N.; Nyffenegger-Jann, N.J.; Voss, M.; Beisswenger, C.; Sunderkötter, C.; Molle, V.; Dinh, Q.T.; Lammert, F.; et al. CcpA Affects Infectivity of Staphylococcus aureus in a Hyperglycemic Environment. Front. Cell. Infect. Microbiol. 2017, 7, 172. [Google Scholar] [CrossRef]
- Liao, X.; Yang, F.; Wang, R.; He, X.; Li, H.; Kao, R.Y.; Xia, W.; Sun, H. Identification of Catabolite Control Protein A from Staphylococcus aureus as a Target of Silver Ions. Chem. Sci. 2017, 8, 8061–8066. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, T.; Zhang, B.; Baronian, G.; Schulthess, B.; Homerova, D.; Grubmüller, S.; Kutzner, E.; Gaupp, R.; Bertram, R.; Powers, R.; et al. Catabolite control protein E (CcpE) is a LysR-type transcriptional regulator of tricarboxylic acid cycle activity in Staphylococcus aureus. J. Biol. Chem. 2013, 288, 36116–36128. [Google Scholar] [CrossRef] [Green Version]
- Giachino, P.; Engelmann, S.; Bischoff, M. Sigma(B) activity depends on RsbU in Staphylococcus aureus. J. Bacteriol. 2001, 183, 1843–1852. [Google Scholar] [CrossRef] [Green Version]
- Monk, I.R.; Shah, I.M.; Xu, M.; Tan, M.W.; Foster, T.J. Transforming the untransformable: Application of direct transformation to manipulate genetically Staphylococcus aureus and Staphylococcus epidermidis. mBio 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Leibig, M.; Krismer, B.; Kolb, M.; Friede, A.; Gotz, F.; Bertram, R. Marker removal in staphylococci via Cre recombinase and different lox sites. Appl. Environ. Microbiol. 2008, 74, 1316–1323. [Google Scholar] [CrossRef] [Green Version]
- Duthie, E.S. Variation in the antigenic composition of staphylococcal coagulase. J. Gen. Microbiol. 1952, 7, 320–326. [Google Scholar] [CrossRef] [Green Version]
- Kreiswirth, B.N.; Lofdahl, S.; Betley, M.J.; O’Reilly, M.; Schlievert, P.M.; Bergdoll, M.S.; Novick, R.P. The toxic shock syndrome exotoxin structural gene is not detectably transmitted by a prophage. Nature 1983, 305, 709–712. [Google Scholar] [CrossRef]
- Iordanescu, S.; Surdeanu, M. Two Restriction and Modification Systems in Staphylococcus aureus NCTC8325. Microbiology 1976, 96, 277–281. [Google Scholar] [CrossRef] [Green Version]
- Gaupp, R.; Wirf, J.; Wonnenberg, B.; Biegel, T.; Eisenbeis, J.; Graham, J.; Herrmann, M.; Lee, C.Y.; Beisswenger, C.; Wolz, C.; et al. RpiRc Is a Pleiotropic Effector of Virulence Determinant Synthesis and Attenuates Pathogenicity in Staphylococcus aureus. Infect. Immun. 2016, 84, 2031–2041. [Google Scholar] [CrossRef] [Green Version]
- Beenken, K.E.; Blevins, J.S.; Smeltzer, M.S. Mutation of sarA in Staphylococcus aureus limits biofilm formation. Infect. Immun. 2003, 71, 4206–4211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Nandakumar, R.; Sadykov, M.R.; Madayiputhiya, N.; Luong, T.T.; Gaupp, R.; Lee, C.Y.; Somerville, G.A. RpiR homologues may link Staphylococcus aureus RNAIII synthesis and pentose phosphate pathway regulation. J. Bacteriol. 2011, 193, 6187–6196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rupp, M.E.; Ulphani, J.S.; Fey, P.D.; Bartscht, K.; Mack, D. Characterization of the importance of polysaccharide intercellular adhesin/hemagglutinin of Staphylococcus epidermidis in the pathogenesis of biomaterial-based infection in a mouse foreign body infection model. Infect. Immun. 1999, 67, 2627–2632. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, M.T.; Manso, A.S.; Gaspar, P.; Pinho, M.G.; Neves, A.R. Effect of oxygen on glucose metabolism: Utilization of lactate in Staphylococcus aureus as revealed by in vivo NMR studies. PLoS ONE 2013, 8, e58277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitko, N.P.; Grosser, M.R.; Khatri, D.; Lance, T.R.; Richardson, A.R. Expanded glucose import capability affords Staphylococcus aureus optimized glycolytic flux during infection. mBio 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, S.R.; Fouts, D.E.; Archer, G.L.; Mongodin, E.F.; Deboy, R.T.; Ravel, J.; Paulsen, I.T.; Kolonay, J.F.; Brinkac, L.; Beanan, M.; et al. Insights on evolution of virulence and resistance from the complete genome analysis of an early methicillin-resistant Staphylococcus aureus strain and a biofilm-producing methicillin-resistant Staphylococcus epidermidis strain. J. Bacteriol. 2005, 187, 2426–2438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deutscher, J.; Herro, R.; Bourand, A.; Mijakovic, I.; Poncet, S. P-Ser-HPr--a link between carbon metabolism and the virulence of some pathogenic bacteria. Biochim. Biophys. Acta 2005, 1754, 118–125. [Google Scholar] [CrossRef]
- Pullen, K.; Rajagopal, P.; Branchini, B.R.; Huffine, M.E.; Reizer, J.; Saier, M.H., Jr.; Scholtz, J.M.; Klevit, R.E. Phosphorylation of serine-46 in HPr, a key regulatory protein in bacteria, results in stabilization of its solution structure. Protein Sci. A Publ. Protein Soc. 1995, 4, 2478–2486. [Google Scholar] [CrossRef] [Green Version]
- Thapar, R.; Nicholson, E.M.; Rajagopal, P.; Waygood, E.B.; Scholtz, J.M.; Klevit, R.E. Influence of N-cap mutations on the structure and stability of Escherichia coli HPr. Biochemistry 1996, 35, 11268–11277. [Google Scholar] [CrossRef] [PubMed]
- Sadykov, M.R.; Hartmann, T.; Mattes, T.A.; Hiatt, M.; Jann, N.J.; Zhu, Y.; Ledala, N.; Landmann, R.; Herrmann, M.; Rohde, H.; et al. CcpA coordinates central metabolism and biofilm formation in Staphylococcus epidermidis. Microbiology 2011, 157, 3458–3468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, T.; Baronian, G.; Nippe, N.; Voss, M.; Schulthess, B.; Wolz, C.; Eisenbeis, J.; Schmidt-Hohagen, K.; Gaupp, R.; Sunderkötter, C.; et al. The catabolite control protein E (CcpE) affects virulence determinant production and pathogenesis of Staphylococcus aureus. J. Biol. Chem. 2014, 289, 29701–29711. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Liu, X.; Chen, F.; Di, H.; Xu, B.; Zhou, L.; Deng, X.; Wu, M.; Yang, C.G.; Lan, L. Metabolic sensor governing bacterial virulence in Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2014, 111, E4981–E4990. [Google Scholar] [CrossRef] [Green Version]
- Balasubramanian, D.; Ohneck, E.A.; Chapman, J.; Weiss, A.; Kim, M.K.; Reyes-Robles, T.; Zhong, J.; Shaw, L.N.; Lun, D.S.; Ueberheide, B.; et al. Staphylococcus aureus coordinates leukocidin expression and pathogenesis by sensing metabolic fluxes via RpiRc. mBio 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montgomery, C.P.; Boyle-Vavra, S.; Roux, A.; Ebine, K.; Sonenshein, A.L.; Daum, R.S. CodY deletion enhances in vivo virulence of community-associated methicillin-resistant Staphylococcus aureus clone USA300. Infect. Immun. 2012, 80, 2382–2389. [Google Scholar] [CrossRef] [Green Version]
- Ha, J.H.; Hauk, P.; Cho, K.; Eo, Y.; Ma, X.; Stephens, K.; Cha, S.; Jeong, M.; Suh, J.Y.; Sintim, H.O.; et al. Evidence of link between quorum sensing and sugar metabolism in Escherichia coli revealed via cocrystal structures of LsrK and HPr. Sci. Adv. 2018, 4, eaar7063. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Description 1 | Reference or Source |
|---|---|---|
| S. aureus | ||
| Newman | Mouse pathogenic laboratory strain (ATCC 25904) | [30] |
| RN4220 | NCTC8325-4 derivative, acceptor of foreign DNA | [31] |
| SA113 | PIA-dependent biofilm producer (ATCC 35556), agr rsbU | [32] |
| Nm ccpA | MST14; Newman ΔccpA::tet(L); TcR | [15] |
| Nm ptsH | Newman ΔptsH::lox72 | This study |
| Nm ptsH-aph | Newman ΔptsH::lox66-aphaIII-lox71; KanR | This study |
| Nm ptsH::ptsH | Newman ΔptsH::pBT ptsH; TcR | This study |
| Nm ptsH* | Newman ΔptsH::lox72 pBTptsH*; TcR | This study |
| Nm ccpA_ptsH | Newman ΔccpA::tet(L) ΔptsH::lox72; TcR | This study |
| RN4220 ptsH | RN4220 ΔptsH::lox72 | This study |
| SA113 ccpA | KS66; SA113 ΔccpA::tet(L); TcR | [19] |
| SA113 ptsH | SA113 ΔptsH::lox72 | This study |
| SA113 ptsH::ptsH | SA113 ΔptsH::pBT ptsH; TcR | This study |
| SA113 ptsH* | SA113 ΔptsH::lox72 pBTptsH*; TcR | This study |
| SA113 ccpA_ptsH | SA113 ΔccpA::tet(L) ΔptsH::lox72; TcR | This study |
| E. coli | ||
| DH5α | Cloning strain | Invitrogen |
| DC10B | Δdcm in the DH10B background; Dam methylation only | [28] |
| Plasmids | ||
| pBT | S. aureus suicide plasmid; tet(L) | [27] |
| pBT lox-aph | pBT derivative harboring lox66-aphAIII-lox71; tet(L), aphIII | [26] |
| pRAB1 | Temperature sensitive E. coli-S. aureus shuttle plasmid, expression of cre in staphylococci; cat, bla | [29] |
| pBT ‘ptsI | pBT derivative harboring a C-terminal ptsI fragment; tet(L) | This study |
| pBT ptsH1 | pBT derivative harboring a T136G ptsH variant; tet(L) | This study |
| pBT ptsH KO | pBT derivative harboring the genomic regions flanking ptsH and lox66-aphAIII-lox71 of pBT lox-aph; aphIII, tet(L) | This study |
| Strain | Generation Time (min) 1 | p Value 2 |
|---|---|---|
| Newman | 28.6 ± 1.9 | |
| Nm ptsH | 34.8 ± 1.6 | <0.01 |
| Nm ptsH::ptsH | 27.8 ± 1.3 | 0.34 |
| Nm ptsH* | 33.3 ± 1.4 | <0.01 |
| Nm ccpA | 30.0 ± 1.3 | 0.19 |
| Nm ccpA_ptsH | 34.3 ± 1.0 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pätzold, L.; Brausch, A.-C.; Bielefeld, E.-L.; Zimmer, L.; Somerville, G.A.; Bischoff, M.; Gaupp, R. Impact of the Histidine-Containing Phosphocarrier Protein HPr on Carbon Metabolism and Virulence in Staphylococcus aureus. Microorganisms 2021, 9, 466. https://doi.org/10.3390/microorganisms9030466
Pätzold L, Brausch A-C, Bielefeld E-L, Zimmer L, Somerville GA, Bischoff M, Gaupp R. Impact of the Histidine-Containing Phosphocarrier Protein HPr on Carbon Metabolism and Virulence in Staphylococcus aureus. Microorganisms. 2021; 9(3):466. https://doi.org/10.3390/microorganisms9030466
Chicago/Turabian StylePätzold, Linda, Anne-Christine Brausch, Evelyn-Laura Bielefeld, Lisa Zimmer, Greg A. Somerville, Markus Bischoff, and Rosmarie Gaupp. 2021. "Impact of the Histidine-Containing Phosphocarrier Protein HPr on Carbon Metabolism and Virulence in Staphylococcus aureus" Microorganisms 9, no. 3: 466. https://doi.org/10.3390/microorganisms9030466
APA StylePätzold, L., Brausch, A. -C., Bielefeld, E. -L., Zimmer, L., Somerville, G. A., Bischoff, M., & Gaupp, R. (2021). Impact of the Histidine-Containing Phosphocarrier Protein HPr on Carbon Metabolism and Virulence in Staphylococcus aureus. Microorganisms, 9(3), 466. https://doi.org/10.3390/microorganisms9030466