Diversity of Microorganisms in Biocrusts Surrounding Highly Saline Potash Tailing Piles in Germany


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
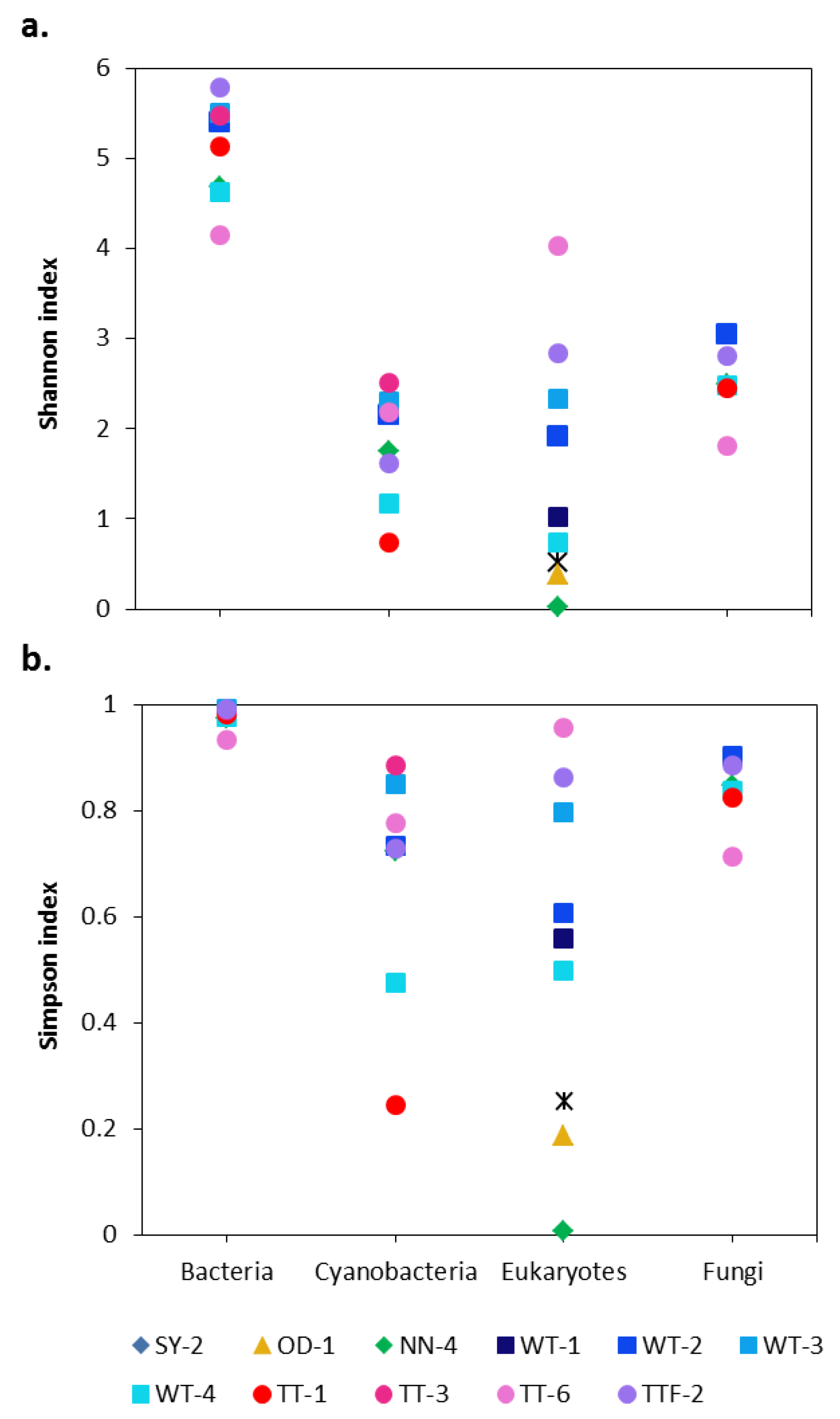
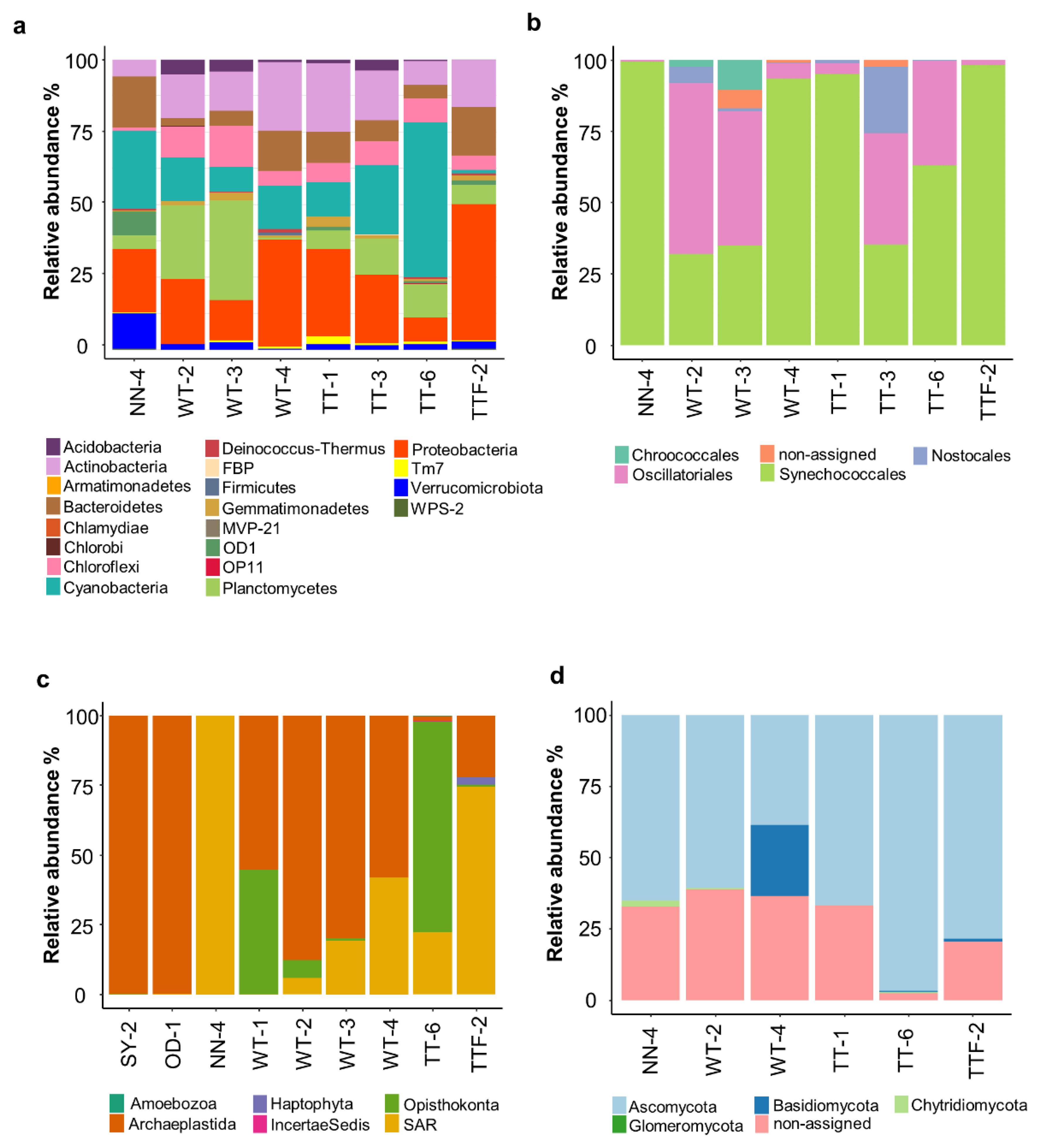
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cañedo-Argüelles, M.; Kefford, B.J.; Piscart, C.; Prat, N.; Schäfer, R.B.; Schulz, C.J. Salinisation of rivers: An urgent ecological issue. Environ. Pollut. 2013, 173, 157–167. [Google Scholar] [CrossRef]
- Belnap, J.; Weber, B.; Büdel, B. Biological Soil Crusts as an Organizing Principle in Drylands; Springer International Publishing: Cham, Switzerland, 2016; pp. 3–13. [Google Scholar] [CrossRef]
- Garve, E.; Garve, V. Halophyten an Kalihalden in Deutschland und Frankreich (Eisass). Tuexenia 2000, 20, 375–417. [Google Scholar]
- John, H. Zur Ausbreitung von Halophyten und salztoleranten Pflanzen in der Umgebung von Kali-Rückstandshalden am Beispiel des FND “Salzstelle bei Teutschenthal-Bahnhof” (Saalkreis). Mitt. Florist. Kart. Sachsen-Anhalt 2000, 5, 175–197. [Google Scholar]
- Flügel, H.-J.; Brenner, U. Die Käferfauna der Salzstellen bei Heringen (Insecta: Coleoptera), ein 50-jähriges Gemeinschaftsprojekt der Arbeitsgemeinschaft Hessischer Koleopterologen. Mitt. Internat. Entomol. Ver. Frankfurt A. M. 2013, 38, 119–165. [Google Scholar]
- Trost, M. Die Historische und Aktuelle Bestandssituation der Halobionten und Halophilen Laufkäferfauna (Coleoptera, Carabidae) im Gebiet der Mansfelder Seen Westlich von Halle/Saale (Sachsen-Anhalt). Hercynia N. F. 2006, 39, 121–149. [Google Scholar]
- Canfora, L.; Vendramin, E.; Antisari, L.V.; Lo Papa, G.; Dazzi, C.; Benedetti, A.; Iavazzo, P.; Adamo, P.; Jungblut, A.D.; Pinzari, F. Compartmentalization of gypsum and halite associated with cyanobacteria in saline soil crusts. FEMS Microbiol. Ecol. 2016, 92, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Cantrell, S.A.; Casillas-Martínez, L.; Molina, M. Characterization of fungi from hypersaline environments of solar salterns using morphological and molecular techniques. Mycol. Res. 2006, 110, 962–970. [Google Scholar] [CrossRef] [PubMed]
- Evans, S.; Hansen, R.W.; Stone, H.M.; Schneegurt, M.A. Isolation and characterization of halotolerant soil fungi from the great salt plains of Oklahoma (USA). Cryptogam. Mycol. 2013, 34, 329–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiman, R.; Ford, L.; Ducros, V.; Lafond, J.L.; Guiraud, P. First survey of fungi in hypersaline soil and water of Mono Lake area (California). Antonie Van Leeuwenhoek 2004, 85, 69–83. [Google Scholar] [CrossRef]
- Bachran, M.; Kluge, S.; Lopez-Fernandez, M.; Cherkouk, A. Microbial Diversity in an Arid, Naturally Saline Environment. Microb. Ecol. 2019, 78, 494–505. [Google Scholar] [CrossRef]
- Eilmus, S.; Rösch, C.; Bothe, H. Prokaryotic life in a potash-polluted marsh with emphasis on N-metabolizing microorganisms. Environ. Pollut. 2007, 146, 478–491. [Google Scholar] [CrossRef]
- Fourçans, A.; De Oteyza, T.G.; Wieland, A.; Solé, A.; Diestra, E.; Van Bleijswijk, J.; Grimalt, J.O.; Kühl, M.; Esteve, I.; Muyzer, G.; et al. Characterization of functional bacterial groups in a hypersaline microbial mat community (Salins-de-Giraud, Camargue, France). FEMS Microbiol. Ecol. 2004, 51, 55–70. [Google Scholar] [CrossRef] [Green Version]
- Oren, A. Microbial life at high salt concentrations: Phylogenetic and metabolic diversity. Saline Syst. 2008, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Mikhailyuk, T.; Vinogradova, O.M.; Glaser, K.; Demchenko, E.M.; Karsten, U. Diversity of terrestrial algae of cape Kazantip (the sea of azov, Ukraine) and some remarks on their phylogeny and ecology. Int. J. Algae 2018, 20, 313–338. [Google Scholar] [CrossRef]
- Pajares, S.; Escalante, A.E.; Noguez, A.M.; García-Oliva, F.; Martínez-Piedragil, C.; Cram, S.S.; Eguiarte, L.E.; Souza, V. Spatial heterogeneity of physicochemical properties explains differences in microbial composition in arid soils from Cuatro Cienegas, Mexico. PeerJ 2016, 2016, e2459. [Google Scholar] [CrossRef] [Green Version]
- Sommer, V.; Karsten, U.; Glaser, K. Halophilic algal communities in biological soil crusts isolated from potash tailings pile areas. Front. Ecol. Evol. 2020, 8, 46. [Google Scholar] [CrossRef] [Green Version]
- Sommer, V.; Mikhailyuk, T.; Glaser, K.; Karsten, U. Uncovering Unique Green Algae and Cyanobacteria Isolated from Biocrusts in Highly Saline Potash Tailing Pile Habitats, Using an Integrative Approach. Microorganisms 2020, 8, 1667. [Google Scholar] [CrossRef] [PubMed]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. correspondence QIIME allows analysis of high-throughput community sequencing data Intensity normalization improves color calling in SOLiD sequencing. Nat. Publ. Gr. 2010, 7, 335–336. [Google Scholar] [CrossRef]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Abarenkov, K.; Nilsson, R.H.; Larsson, K.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; Pennanen, T.; et al. The UNITE database for molecular identification of fungi—Recent updates and future perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Oksanen, J. Multivariate analysis of ecological communities in R:vegan tutorial. R Doc. 2013, 1–43. [Google Scholar] [CrossRef]
- Clarke, K.; Somerfield, P.; Gorley, R. Testing of null hypotheses in exploratory community analyses: Similarity profiles and biota-environment linkage. J. Exp. Mar. Bio. Ecol. 2008, 366, 56–69. [Google Scholar] [CrossRef]
- Rippin, M.; Lange, S.; Sausen, N.; Becker, B. Biodiversity of biological soil crusts from the Polar Regions revealed by metabarcoding. FEMS Microbiol. Ecol. 2018, 94, 36. [Google Scholar] [CrossRef] [PubMed]
- Pushkareva, E.; Pessi, I.S.; Wilmotte, A.; Elster, J. Cyanobacterial community composition in Arctic soil crusts at different stages of development. FEMS Microbiol. Ecol. 2015, 91, fiv143. [Google Scholar] [CrossRef]
- Pombubpa, N.; Pietrasiak, N.; De Ley, P.; Stajich, J.E. Insights into dryland biocrust microbiome: Geography, soil depth and crust type affect biocrust microbial communities and networks in Mojave Desert, USA. FEMS Microbiol. Ecol. 2020, 96, fiaa125. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Kong, W.; Nan, W.; Zhang, Y. Bacterial diversity and community along the succession of biological soil crusts in the Gurbantunggut Desert, Northern China. J. Basic Microbiol. 2016, 56, 670–679. [Google Scholar] [CrossRef]
- Weber, B.; Tamm, A.; Maier, S.; Rodríguez-Caballero, E. Biological soil crusts of the Succulent Karoo: A review. Afr. J. Range Forage Sci. 2018, 35, 335–350. [Google Scholar] [CrossRef] [Green Version]
- Chilton, A.M.; Neilan, B.A.; Eldridge, D.J. Biocrust morphology is linked to marked differences in microbial community composition. Plant Soil 2018, 429, 65–75. [Google Scholar] [CrossRef]
- Rath, K.M.; Fierer, N.; Murphy, D.V.; Rousk, J. Linking bacterial community composition to soil salinity along environmental gradients. ISME J. 2019, 13, 836–846. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.; Gong, J. A meta-analysis of the publicly available bacterial and archaeal sequence diversity in saline soils. World J. Microbiol. Biotechnol. 2013, 29, 2325–2334. [Google Scholar] [CrossRef]
- Imhoff, J.F. True marine and halophilic anoxygenic phototrophic bacteria. Arch. Microbiol. 2001, 176, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Grimaud, R. Marinobacter. In Handbook of Hydrocarbon and Lipid Microbiology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 1289–1296. [Google Scholar]
- Williams, L.; Loewen-Schneider, K.; Maier, S.; Büdel, B. Cyanobacterial diversity of western European biological soil crusts along a latitudinal gradient. FEMS Microbiol. Ecol. 2016, 92, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pessi, I.S.; Pushkareva, E.; Lara, Y.; Borderie, F.; Wilmotte, A.; Elster, J. Marked Succession of Cyanobacterial Communities Following Glacier Retreat in the High Arctic. Microb. Ecol. 2019, 77, 136–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pushkareva, E.; Pessi, I.S.; Namsaraev, Z.; Mano, M.J.; Elster, J.; Wilmotte, A. Cyanobacteria inhabiting biological soil crusts of a polar desert: Sør Rondane Mountains, Antarctica. Syst. Appl. Microbiol. 2018, 41, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Abed, R.M.M.; Garcia-Pichel, F.; Hernández-Mariné, M. Polyphasic characterization of benthic, moderately halophilic, moderately thermophilic cyanobacteria with very thin trichomes and the proposal of Halomicronema excentricum gen. nov., sp. nov. Arch. Microbiol. 2002, 177, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Housman, D.C.; Powers, H.H.; Collins, A.D.; Belnap, J. Carbon and nitrogen fixation differ between successional stages of biological soil crusts in the Colorado Plateau and Chihuahuan Desert. J. Arid Environ. 2006, 66, 620–634. [Google Scholar] [CrossRef] [Green Version]
- Büdel, B.; Dulic, T.; Darienko, T.; Rybalka, N.; Friedl, T. Cyanobacteria and Algae of Biological Soil Crusts. In Biological Soil Crusts: An Organizing Principle in Drylands; Springer International Publishing: Cham, Switzerland, 2016; pp. 55–80. [Google Scholar]
- Gustavs, L.; Eggert, A.; Michalik, D.; Karsten, U. Physiological and biochemical responses of green microalgae from different habitats to osmotic and matric stress. Protoplasma 2010, 243, 3–14. [Google Scholar] [CrossRef]
- Rossi, F.; Mugnai, G.; De Philippis, R. Complex role of the polymeric matrix in biological soil crusts. Plant Soil 2018, 429, 19–34. [Google Scholar] [CrossRef]
- Osińska-Jaroszuk, M.; Jarosz-Wilkołazka, A.; Jaroszuk-Ściseł, J.; Szałapata, K.; Nowak, A.; Jaszek, M.; Ozimek, E.; Majewska, M. Extracellular polysaccharides from Ascomycota and Basidiomycota: Production conditions, biochemical characteristics, and biological properties. World J. Microbiol. Biotechnol. 2015, 31, 1823–1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pushkareva, E.; Sommer, V.; Barrantes, I.; Karsten, U. Diversity of Microorganisms in Biocrusts Surrounding Highly Saline Potash Tailing Piles in Germany. Microorganisms 2021, 9, 714. https://doi.org/10.3390/microorganisms9040714
Pushkareva E, Sommer V, Barrantes I, Karsten U. Diversity of Microorganisms in Biocrusts Surrounding Highly Saline Potash Tailing Piles in Germany. Microorganisms. 2021; 9(4):714. https://doi.org/10.3390/microorganisms9040714
Chicago/Turabian StylePushkareva, Ekaterina, Veronika Sommer, Israel Barrantes, and Ulf Karsten. 2021. "Diversity of Microorganisms in Biocrusts Surrounding Highly Saline Potash Tailing Piles in Germany" Microorganisms 9, no. 4: 714. https://doi.org/10.3390/microorganisms9040714
APA StylePushkareva, E., Sommer, V., Barrantes, I., & Karsten, U. (2021). Diversity of Microorganisms in Biocrusts Surrounding Highly Saline Potash Tailing Piles in Germany. Microorganisms, 9(4), 714. https://doi.org/10.3390/microorganisms9040714