
Repurposing Drugs for Mayaro Virus: Identification of EIDD-1931, Favipiravir and Suramin as Mayaro Virus Inhibitors



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Compounds
2.3. Replication Kinetics
2.4. End-Point Titration Assay
2.5. Plaque Assay
2.6. Antiviral CPE Reduction and Cytotoxicity Assay
2.7. Virus Yield Assay
2.8. Quantitative Reverse Transcription PCR (qRT-PCR)
2.9. Delay of Treatment Assay
2.10. Statistical Analysis
3. Results
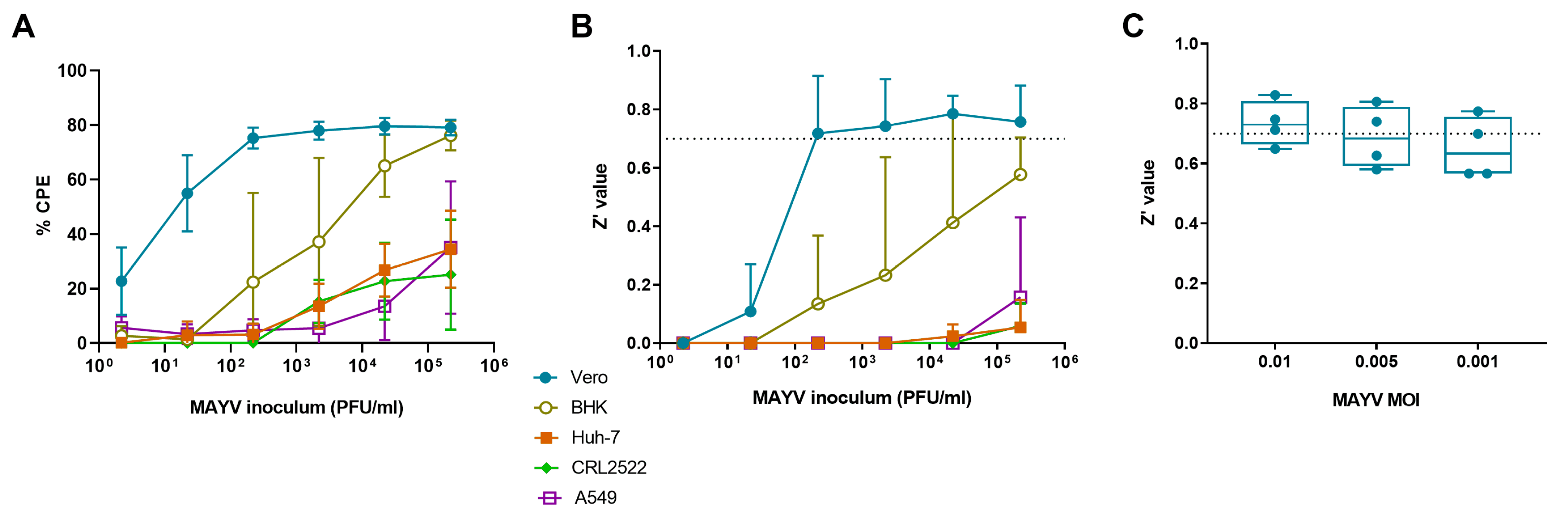
3.1. Selection of Cell Line for MAYV-Induced Cytopathogenic Effect
3.2. Selection of Optimal Virus Inoculum
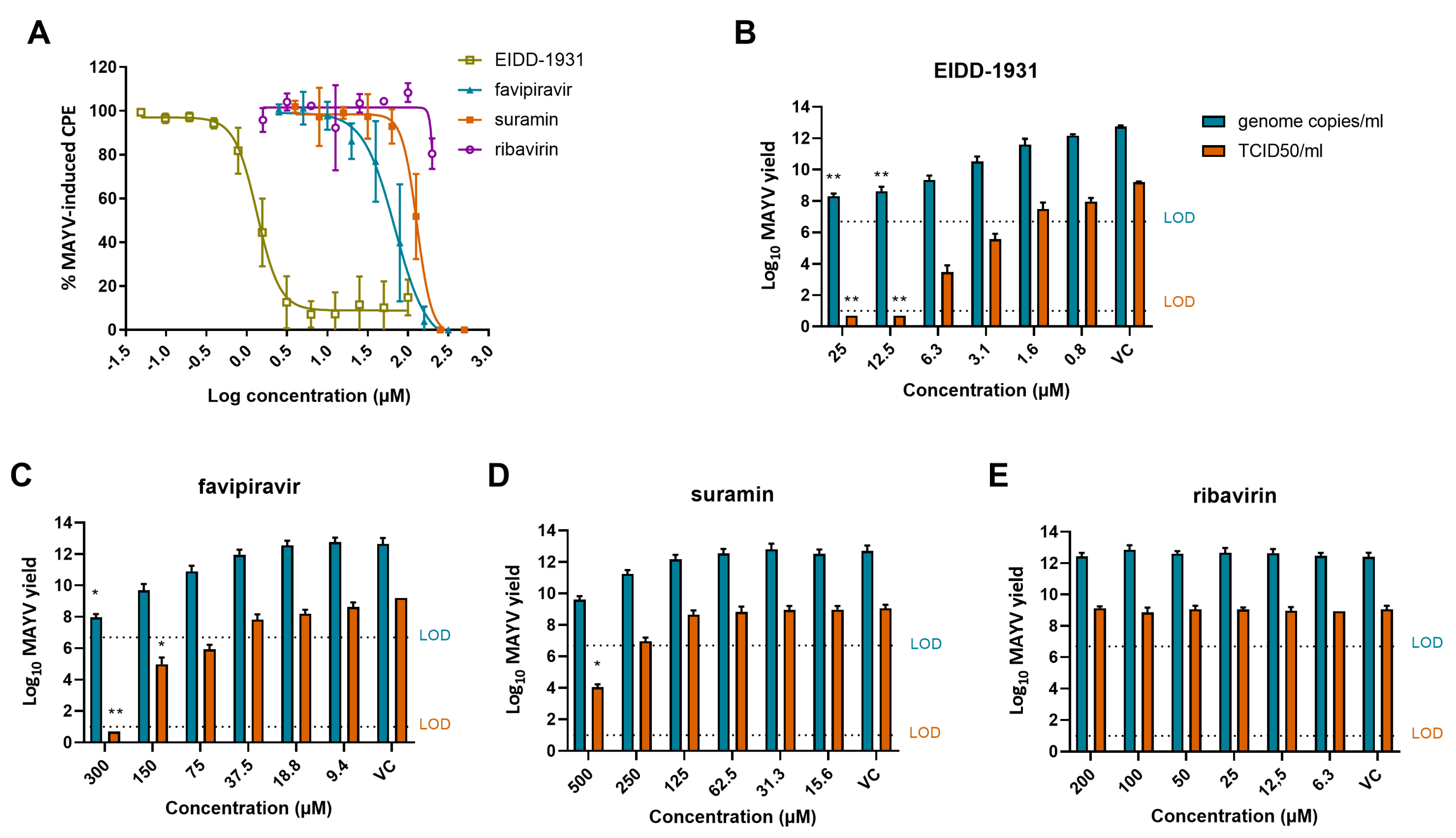
3.3. Anti-MAYV Activity of a Panel of Reference Compounds
3.4. EIDD-1931, Favipiravir and Suramin Reduce MAYV Yield
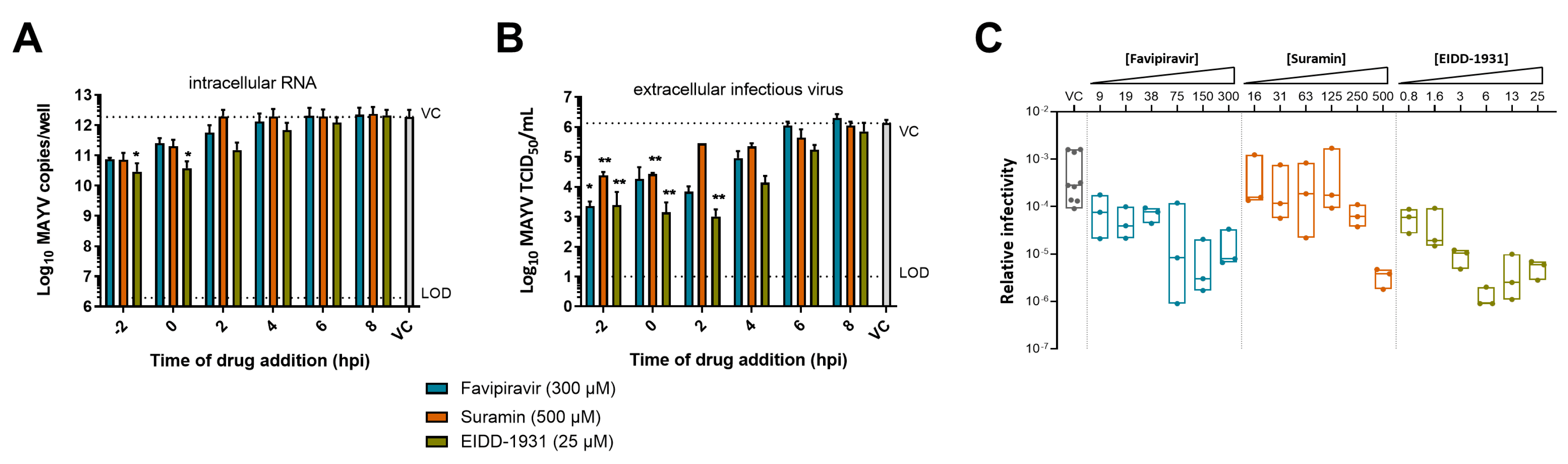
3.5. Mechanism of Action of EIDD-1931, Favipiravir and Suramin against MAYV
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mejía, C.R.; López-Vélez, R. Tropical arthritogenic alphaviruses. Reum. Clin. 2018, 14, 97–105. [Google Scholar] [CrossRef]
- Acosta-Ampudia, Y.; Monsalve, D.M.; Rodríguez, Y.; Pacheco, Y.; Anaya, J.-M.; Ramírez-Santana, C. Mayaro: An emerging viral threat? Emerg. Microbes Infect. 2018, 7, 1–11. [Google Scholar] [CrossRef]
- Levi, L.I.; Vignuzzi, M. Arthritogenic alphaviruses: A worldwide emerging threat? Microorganisms 2019, 7, 133. [Google Scholar] [CrossRef] [Green Version]
- Long, K.C.; Ziegler, S.A.; Thangamani, S.; Hausser, N.L.; Kochel, T.J.; Higgs, S.; Tesh, R.B. Experimental transmission of Mayaro virus by Aedes aegypti. Am. J. Trop. Med. Hyg. 2011, 85, 750–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezzi, L.; Rodriguez-Morales, A.J.; Reusken, C.B.; Ribeiro, G.S.; LaBeaud, A.D.; Lourenço-de-Oliveira, R.; Brasil, P.; Lecuit, M.; Failloux, A.B.; Gallian, P.; et al. GloPID-R report on Chikungunya, o’nyong-nyong and Mayaro virus, part 3: Epidemiological distribution of Mayaro virus. Antivir. Res. 2019, 172. [Google Scholar] [CrossRef] [PubMed]
- Ganjian, N.; Riviere-Cinnamond, A. Mayaro virus in Latin America and the Caribbean. Rev. Panam. Salud Pública 2020, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson, J.; Sammon, M.; Garg, M. Dengue, Zika and Chikungunya: Emerging Arboviruses in the New World. West. J. Emerg. Med. 2016, 17, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Mackay, I.M.; Arden, K.E. Mayaro virus: A forest virus primed for a trip to the city? Microbes Infect. 2016, 18, 724–734. [Google Scholar] [CrossRef]
- Robinson, D.; Cole, F.J.; McManus, A.; Pedersen, C.J. Inactivated Mayaro vaccine produced in human diploid cell cultures. Mil. Med. 1976, 141, 163–166. [Google Scholar] [CrossRef]
- Weise, W.J.; Hermance, M.E.; Forrester, N.; Adams, A.P.; Langsjoen, R.; Gorchakov, R.; Wang, E.; Alcorn, M.D.H.; Tsetsarkin, K.; Weaver, S.C. A novel live-attenuated vaccine candidate for Mayaro fever. PLoS Negl. Trop. Dis. 2014, 8, e2969. [Google Scholar] [CrossRef]
- de Oliveira Mota, M.T.; Costa, V.V.; Sugimoto, M.A.; de Freitas Guimarães, G.; Queiroz-Junior, C.M.; Moreira, T.P.; de Sousa, C.D.; Santos, F.M.; Queiroz, V.F.; Passos, I.; et al. In-depth characterization of a novel live-attenuated Mayaro virus vaccine candidate using an immunocompetent mouse model of Mayaro disease. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.; Kudchodkar, S.B.; Reuschel, E.L.; Asijaid, K.; Borole, P.; Ho, M.; Wojtak, K.; Reed, C.; Ramos, S.; Bopp, N.E.; et al. Protective immunity by an engineered DNA vaccine for mayaro virus. PLoS Negl. Trop. Dis. 2019, 13, e0005122. [Google Scholar] [CrossRef] [PubMed]
- Amorim, R.; De Meneses, M.D.F.; Borges, J.C.; Pinheiro, L.C.D.S.; Caldas, L.A.; Cirne-Santos, C.C.; De Mello, M.V.P.; De Souza, A.M.T.; Castro, H.C.; Paixão, I.C.N.D.P.; et al. Thieno [2,3-b] pyridine derivatives: A new class of antiviral drugs against Mayaro virus. Arch. Virol. 2017, 162, 1577–1587. [Google Scholar] [CrossRef] [PubMed]
- Pereira, H.; Rebello, M. Inhibition of Mayaro virus replication by cerulenin in Aedes albopictus cells. Acta Virol. 1998, 42, 383–388. [Google Scholar] [PubMed]
- Da Costa, L.; Rebello, M. Effect of brefeldin A on Mayaro virus replication in Aedes albopictus and Vero cells. Acta Virol. 1999, 43, 357–360. [Google Scholar] [PubMed]
- Caldas, L.A.; Ferreira, D.F.; Pereira Freitas, T.R. Major article prostaglandin A 1 triggers Mayaro virus inhibition and heat shock protein 70 expression in an epithelial cell model. Rev. Soc. Bras. Med. Trop. 2018, 51, 584–590. [Google Scholar] [CrossRef]
- Carvalho, C.A.M.; Sousa, I.P.; Silva, J.L.; Oliveira, A.C.; Gonçalves, R.B.; Gomes, A.M.O. Inhibition of Mayaro virus infection by bovine lactoferrin. Virology 2014, 452–453, 297–302. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, P.G.; Ferraz, A.C.; Figueiredo, J.E.; Lima, C.F.; Rodrigues, V.G.; Taranto, A.G.; Ferreira, J.M.S.; Brandão, G.C.; Vieira-Filho, S.A.; Duarte, L.P.; et al. Detection of the antiviral activity of epicatechin isolated from Salacia crassifolia (Celastraceae) against Mayaro virus based on protein C homology modelling and virtual screening. Arch. Virol. 2018, 163, 1567–1576. [Google Scholar] [CrossRef]
- dos Santos, A.E.; Kuster, R.M.; Yamamoto, K.A.; Salles, T.S.; Campos, R.; de Meneses, M.D.F.; Soares, M.R.; Ferreira, D. Quercetin and quercetin 3-O- glycosides from Bauhinia longifolia (Bong) Steud. show anti-Mayaro virus activity. Parasites Vectors 2014, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Spindola, K.C.W.; Simas, N.K.; Salles, T.S.; de Meneses, M.D.F.; Sato, A.; Ferreira, D.; Romão, W.; Kuster, R.M. Anti-Mayaro virus activity of Cassia australis extracts (Fabaceae, Leguminosae). Parasites Vectors 2014, 7, 1–7. [Google Scholar] [CrossRef]
- Camini, F.C.; Da Silva, T.F.; Caetano, C.C.D.S.; Almeida, L.T.; Ferraz, A.C.; Vitoreti, V.M.A.; Silva, B.; Silva, S.D.Q.; de Magalhães, J.C.; Magalhães, C.L.D.B. Antiviral activity of silymarin against Mayaro virus and protective effect in virus-induced oxidative stress. Antivir. Res. 2018, 158, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.-H.; Chung, T.D.Y.; Oldenburg, K.R. A simple statistical parameter for use in evaluation and validation of high throughput screening assays. J. Biomol. Screen. 1999, 4, 67–73. [Google Scholar] [CrossRef]
- Wiggins, K.; Eastmond, B.; Alto, B.W. Transmission potential of Mayaro virus in Florida Aedes aegypti and Aedes albopictus mosquitoes. Med. Vet. Entomol. 2018, 32, 436–442. [Google Scholar] [CrossRef] [Green Version]
- Delogu, I.; Pastorino, B.; Baronti, C.; Nougairède, A.; Bonnet, E.; de Lamballerie, X. In vitro antiviral activity of arbidol against Chikungunya virus and characteristics of a selected resistant mutant. Antivir. Res. 2011, 90, 99–107. [Google Scholar] [CrossRef]
- Khan, M.; Santhosh, S.R.; Tiwari, M.; Rao, L.; Parida, M. Assessment of In vitro prophylactic and therapeutic efficacy of chloroquine against Chikungunya virus In vero cells. J. Med. Virol. 2010, 82, 817–824. [Google Scholar] [CrossRef]
- Delang, L.; Li, C.; Tas, A.; Quérat, G.; Albulescu, I.C.; De Burghgraeve, T.; Guerrero, N.A.S.; Gigante, A.; Piorkowski, G.; Decroly, E.; et al. The viral capping enzyme nsP1: A novel target for the inhibition of chikungunya virus infection. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef]
- Abdelnabi, R.; Kovacikova, K.; Moesslacher, J.; Donckers, K.; Battisti, V.; Leyssen, P.; Langer, T.; Puerstinger, G.; Quérat, G.; Li, C.; et al. Novel class of Chikungunya virus small molecule inhibitors that targets the viral capping machinery. Antimicrob. Agents Chemother. 2020, 64, e00649-20. [Google Scholar] [CrossRef]
- Abulescu, I.C.; Van Hoolwerff, M.; Wolters, L.A.; Bottaro, E.; Nastruzzi, C. Suramin inhibits chikungunya virus replication through multiple mechanisms. Antivir. Res. 2015, 21, 39–46. [Google Scholar] [CrossRef]
- Albulescu, I.C.; White-Scholten, L.; Tas, A.; Hoornweg, T.E.; Ferla, S.; Kovacikova, K.; Smit, J.M.; Brancale, A.; Snijder, E.J.; van Hemert, M.J. Suramin inhibits chikungunya virus replication by interacting with virions and blocking the early steps of infection. Viruses 2020, 12, 314. [Google Scholar] [CrossRef] [Green Version]
- Graci, J.D.; Cameron, C.E. Mechanisms of action of ribavirin against distinct viruses. Rev. Med. Virol. 2006, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Delang, L.; Abdelnabi, R.; Neyts, J. Favipiravir as a potential countermeasure against neglected and emerging RNA viruses. Antivir. Res. 2018, 153, 85–94. [Google Scholar] [CrossRef]
- Borrego, B.; de Ávila, A.I.; Domingo, E.; Brun, A. Lethal mutagenesis of Rift Valley fever virus induced by favipiravir. Antimicrob. Agents Chemother. 2019, 63, e00669-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escribano-Romero, E.; De Oya, N.J.; Domingo, E.; Saiza, J.C. Extinction of west nile virus by favipiravir through lethal mutagenesis. Antimicrob. Agents Chemother. 2017, 61, e01400-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zmurko, J.; Margues, R.E.; Schols, D.; Verbeken, E.; Kaptein, S.; Neyts, J. The viral polymerase inhibitor 7-Deaza-2’-C-methyladenosine is a potent inhibitor of in vitro Zika virus replication and delays disease progression in a robust mouse infection model. PLoS Negl. Trop. Dis. 2016, 10, e0004695. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Tseng, C.K.; Wu, Y.H.; Kaushik-Basu, N.; Lin, C.K.; Chen, W.C.; Wu, H.N. Characterization of the activity of 2′-C-methylcytidine against dengue virus replication. Antivir. Res. 2015, 116, 1–9. [Google Scholar] [CrossRef]
- Painter, G.R.; Bowen, R.A.; Bluemling, G.R.; DeBergh, J.; Edpuganti, V.; Gruddanti, P.R.; Guthrie, D.B.; Hager, M.; Kuiper, D.L.; Lockwood, M.A.; et al. The prophylactic and therapeutic activity of a broadly active ribonucleoside analog in a murine model of intranasal venezuelan equine encephalitis virus infection. Antivir. Res. 2019, 171, 104597. [Google Scholar] [CrossRef] [PubMed]
- Ehteshami, M.; Tao, S.; Zandi, K.; Hsiao, H.-M.; Jiang, Y.; Hammond, E.; Amblard, F.; Russell, O.O.; Merits, A.; Schinazi, R.F. Characterization of B-D-N4-Hydroxycytidine as a novel inhibitor of Chikungunya virus. Antimicrob. Agents Chemother. 2017, 61, e02395-16. [Google Scholar] [CrossRef] [Green Version]
- De Clercq, E. New nucleoside analogues for the treatment of hemorrhagic fever virus infections. Chem. Asian J. 2019, 14, 3962–3968. [Google Scholar] [CrossRef]
- Urakova, N.; Kuznetsova, V.; Crossman, D.K.; Sokratian, A.; Guthrie, D.B.; Kolykhalov, A.A.; Lockwood, M.A.; Natchus, M.G.; Crowley, M.R.; Painter, G.R.; et al. β-D-N4-Hydroxycytidine Is a potent anti-alphavirus compound that induces a high level of mutations in the viral genome. J. Virol. 2018, 92, e01965-17. [Google Scholar] [CrossRef] [Green Version]
- Gould, E.; Pettersson, J.; Higgs, S.; Charrel, R.; de Lamballerie, X. Emerging arboviruses: Why today? One Health 2017, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Vairo, F.; Haider, N.; Kock, R.; Ntoumi, F.; Ippolito, G.; Zumla, A. Chikungunya: Epidemiology, pathogenesis, clinical features, management, and prevention. Infect. Dis. Clin. N. Am. 2019, 33, 1003–1025. [Google Scholar] [CrossRef] [PubMed]
- WHO Chikungunya. Available online: https://www.who.int/news-room/fact-sheets/detail/chikungunya (accessed on 12 January 2021).
- Schilte, C.; Stoikovsky, F.; Couderc, T.; Madec, Y.; Carpentier, F.; Kassab, S.; Albert, M.L.; Lecuit, M.; Michault, A. Chikungunya virus-associated long-term arthralgia: A 36-month prospective longitudinal study. PLoS Negl. Trop. 2013, 7, e2137. [Google Scholar] [CrossRef]
- Li, X.; Zhang, H.; Zhang, Y.; Li, J.; Wang, Z.; Deng, C.; Jardim, A.C.G.; Terzian, A.C.B.; Nogueira, M.L.; Zhang, B. Development of a rapid antiviral screening assay based on eGFP reporter virus of Mayaro virus. Antivir. Res. 2019, 168, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Lani, R.; Hassandarvish, P.; Chiam, C.W.; Moghaddam, E.; Chu, J.J.H.; Rausalu, K.; Merits, A.; Higgs, S.; Vanlandingham, D.; Abu Bakar, S.; et al. Antiviral activity of silymarin against chikungunya virus. Sci. Rep. 2015, 5, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardiot, D.; Koukni, M.; Smets, W.; Carlens, G.; McNaughton, M.; Kaptein, S.; Dallmeier, K.; Chaltin, P.; Neyts, J.; Marchand, A. discovery of indole derivatives as novel and potent dengue virus inhibitors. J. Med. Chem. 2018, 61, 8390–8401. [Google Scholar] [CrossRef]
- Toots, M.; Yoon, J.; Hart, M.; Natchus, M.G.; Painter, G.R.; Plemper, R.K. Quantitative efficacy paradigms of the influenza clinical drug candidate EIDD-2801 in the Ferret Model. Transl. Res. 2020, 218, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.M.; Wolf, J.D.; Plemper, R.K. Therapeutically administered ribonucleoside analogue MK-4482/EIDD-2801 blocks SARS-CoV-2 transmission in ferrets. Nat. Microbiol. 2021, 6, 11–18. [Google Scholar] [CrossRef]
- Abdelnabi, R.; Delang, L. Antiviral strategies against arthritogenic alphaviruses. Microorganisms 2020, 8, 1365. [Google Scholar] [CrossRef]
- McGeary, R.; Bennett, A.; Tran, Q.; Cosgrove, K.; Ross, B. Suramin: Clinical uses and structure-activity relationships. Mini Rev. Med. Chem. 2008, 8, 1384–1394. [Google Scholar] [CrossRef]
- Malin, J.J.; Suárez, I.; Priesner, V.; Fätkenheuer, G.; Rybniker, J. Remdesivir against COVID-19 and other viral diseases. Clin. Microbiol. Rev. 2021, 34, e00162-20. [Google Scholar] [CrossRef]
- Bugert, J.J.; Hucke, F.; Zanetta, P.; Bassetto, M.; Brancale, A. Antivirals in medical biodefense. Virus Genes 2020, 56, 150–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baranovich, T.; Wong, S.-S.; Armstrong, J.; Marjuki, H.; Webby, R.J.; Webster, R.G.; Govorkova, E.A. T-705 (Favipiravir) induces lethal mutagenesis in influenza A H1N1 viruses In vitro. J. Virol. 2013, 87, 3741–3751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delang, L.; Guerrero, N.S.; Tas, A.; Quérat, G.; Pastorino, B.; Froeyen, M.; Dallmeier, K.; Jochmans, D.; Herdewijn, P.; Bello, F.; et al. Mutations in the chikungunya virus non-structural proteins cause resistance to favipiravir (T-705), a broad-spectrum antiviral. J. Antimicrob. Chemother. 2014, 69, 2770–2784. [Google Scholar] [CrossRef] [PubMed]
- Smee, D.F.; Hurst, B.L.; Egawa, H.; Takahashi, K.; Kadota, T.; Furuta, Y. Intracellular metabolism of favipiravir (T-705) in uninfected and influenza A (H5N1) virus-infected cells. J. Antimicrob. Chemother. 2009, 64, 741–746. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Compound | MOA * | EC50 (µM) | CC50 (µM) |
|---|---|---|---|
| Arbidol | Entry inhibitors [25,26,30] | >60 | >100 |
| Chloroquine | >200 | 145 ± 20 | |
| Suramin | 124 ± 26 | >2000 | |
| EIDD-1931 | Virus replication inhibitors [32,35,36,38,39] | 1.6 ± 0.5 | >100 |
| Favipiravir | 79 ± 4 | 2837 ± 86 | |
| 7DMA | >60 | >600 | |
| 2’CMC | >20 | 167 ± 7 | |
| Galidesivir | >100 | 238 ± 36 | |
| Remdesivir | >100 | 297 ± 101 | |
| Ribavirin | Replication inhibitor + host effects [31] | >200 | 250 ± 81 |
| MADTP | Capping inhibitors [27,28] | >100 | >400 |
| CHVB-032 | >100 | >75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Langendries, L.; Abdelnabi, R.; Neyts, J.; Delang, L. Repurposing Drugs for Mayaro Virus: Identification of EIDD-1931, Favipiravir and Suramin as Mayaro Virus Inhibitors. Microorganisms 2021, 9, 734. https://doi.org/10.3390/microorganisms9040734
Langendries L, Abdelnabi R, Neyts J, Delang L. Repurposing Drugs for Mayaro Virus: Identification of EIDD-1931, Favipiravir and Suramin as Mayaro Virus Inhibitors. Microorganisms. 2021; 9(4):734. https://doi.org/10.3390/microorganisms9040734
Chicago/Turabian StyleLangendries, Lana, Rana Abdelnabi, Johan Neyts, and Leen Delang. 2021. "Repurposing Drugs for Mayaro Virus: Identification of EIDD-1931, Favipiravir and Suramin as Mayaro Virus Inhibitors" Microorganisms 9, no. 4: 734. https://doi.org/10.3390/microorganisms9040734
APA StyleLangendries, L., Abdelnabi, R., Neyts, J., & Delang, L. (2021). Repurposing Drugs for Mayaro Virus: Identification of EIDD-1931, Favipiravir and Suramin as Mayaro Virus Inhibitors. Microorganisms, 9(4), 734. https://doi.org/10.3390/microorganisms9040734