
Stool Serology: Development of a Non-Invasive Immunological Method for the Detection of Enterovirus-Specific Antibodies in Congo Gorilla Faeces

 ,
,  , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Gorilla Faeces Samples
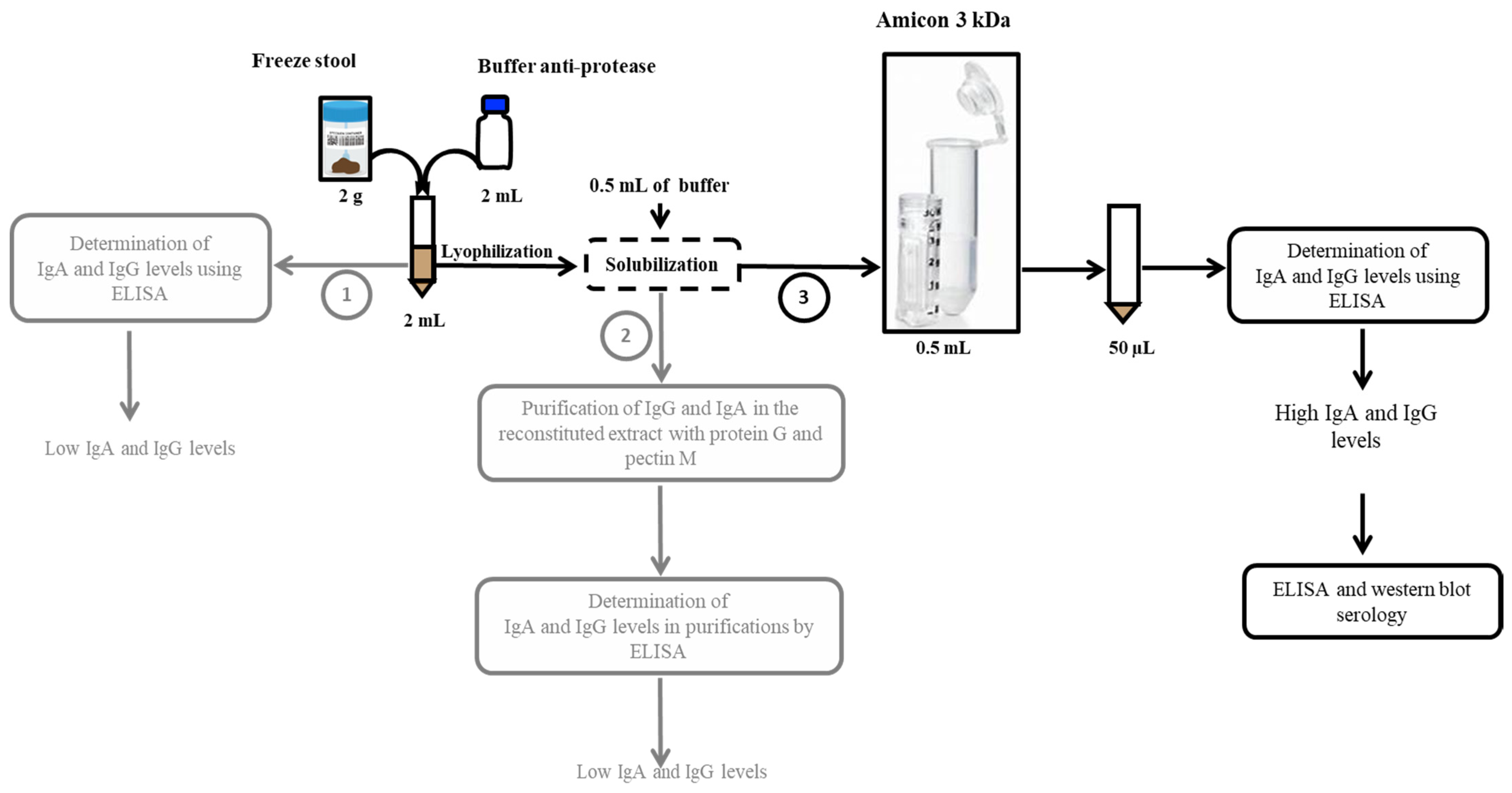
2.2. Concept and Optimisation of the Methodological Approach
2.3. Experimental Protocol Adopted for Stool Protein Extraction
2.4. Quantification of Anti-Poliovirus Immunoglobulins by ELISA
2.5. Characterisation of Enterovirus-Specific Antibodies by Western Blot from Faecal Samples
3. Results
3.1. Differentiation of the Faeces of Individual Gorillas
3.2. Selection of the Stool Preparation Protocol
3.3. Quantification and Detection of Poliovirus-Specific Immunoglobulins by ELISA and Western Blot
3.3.1. Quantification of Poliovirus-Specific Immunoglobulins (IgA-IgG)
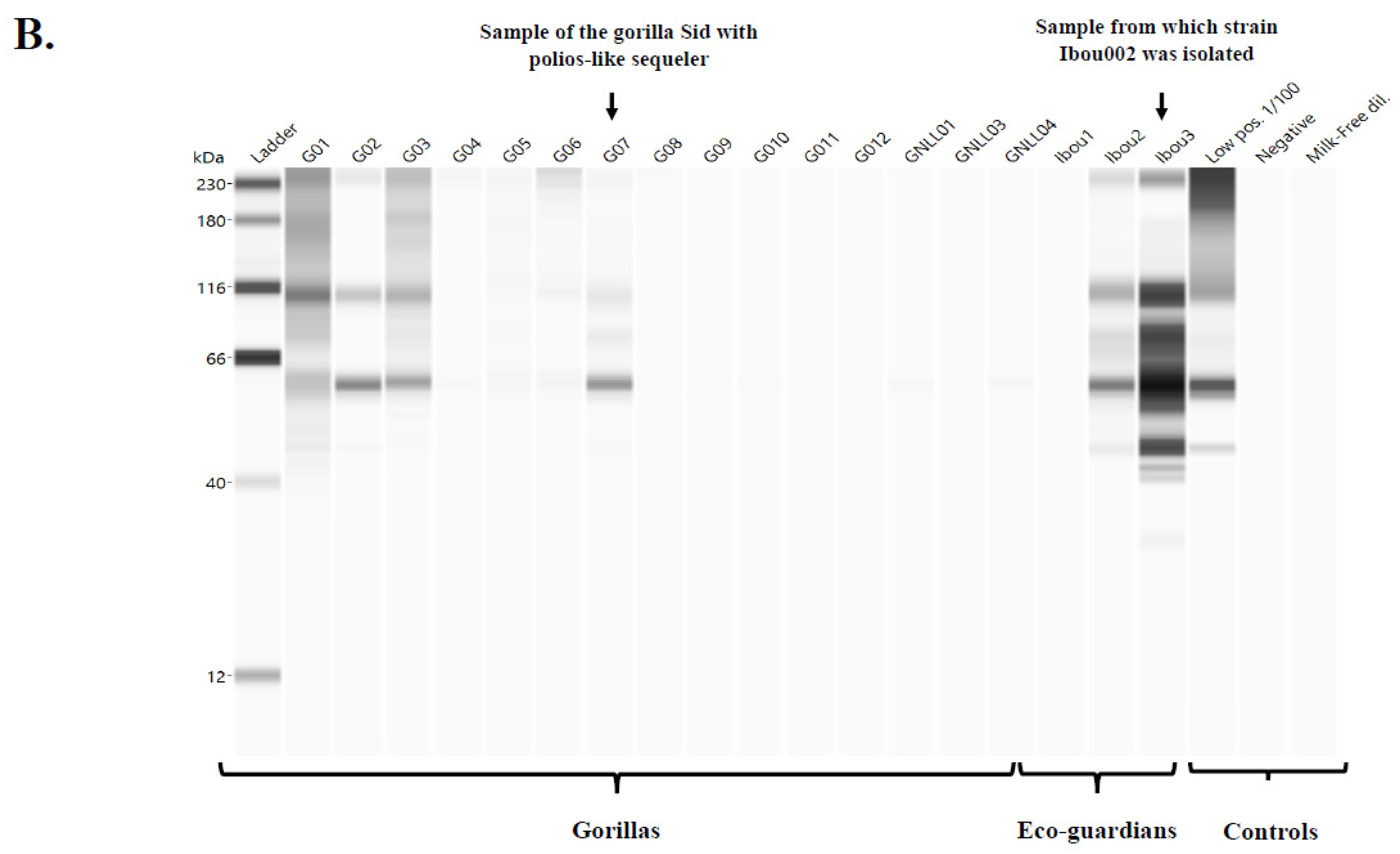
3.3.2. Detection of Poliovirus- and Enterovirus C-Specific Immunoglobulins by Western Blot
Western Blot
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Fields, B.N.; Knipe, D.M.; Howley, P.M. (Eds.) Fields Virology, 5th ed.; Wolters Kluwer Health/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; ISBN 978-0-7817-6060-7. [Google Scholar]
- Blondel, B.; Autret, A.; Brisac, C.; Pelletier, I.; Martin-Latil, S.; Jegouic, S.; Bessaud, M.; Joffret, M.L.; Balanant, J.; Colbère-Garapin, F.; et al. Genetic evolution of poliovirus: Success and difficulties in the eradication of paralytic poliomyelitis. Med. Trop. 2008, 68, 189–202. [Google Scholar]
- Pelletier, J.; Sonenberg, N. Internal Initiation of Translation of Eukaryotic MRNA Directed by a Sequence Derived from Poliovirus RNA. Nature 1988, 334, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Datta, S.D.; Quddus, A.; Vertefeuille, J.F.; Burns, C.C.; Jorba, J.; Wassilak, S.G.F. Progress Toward Polio Eradication—Worldwide, January 2016–March 2018. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 524–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Weekly Epidemiological Record: Progress towards Poliomyelitis Eradication; World Health Organization (WHO): Geneva, Switzerland, 2019. [Google Scholar]
- Duintjer Tebbens, R.J.; Thompson, K.M. Polio Endgame Risks and the Possibility of Restarting the Use of Oral Poliovirus Vaccine. Expert Rev. Vaccines 2018, 17, 739–751. [Google Scholar] [CrossRef] [Green Version]
- Thompson, K.M.; Duintjer Tebbens, R.J. Lessons from the Polio Endgame: Overcoming the Failure to Vaccinate and the Role of Subpopulations in Maintaining Transmission. J. Infect. Dis. 2017, 216, S176–S182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javelle, E.; Raoult, D. Antibiotics against Poliovirus Carriage: An Additional Tool in the Polio Endgame? Clin. Microbiol. Infect. 2020, 26, 542–544. [Google Scholar] [CrossRef]
- Akhtar, R.; Mahmood, N.; Alam, M.M.; Naeem, M.; Zaidi, S.S.Z.; Sharif, S.; Khattak, Z.; Arshad, Y.; Khurshid, A.; Mujtaba, G.; et al. Genetic Epidemiology Reveals 3 Chronic Reservoir Areas With Recurrent Population Mobility Challenging Poliovirus Eradication in Pakistan. Clin. Infect. Dis. 2020, 71, e58–e67. [Google Scholar] [CrossRef]
- Kalkowska, D.A.; Pallansch, M.A.; Thompson, K.M. Updated Modelling of the Prevalence of Immunodeficiency-Associated Long-Term Vaccine-Derived Poliovirus (IVDPV) Excreters. Epidemiol. Infect. 2019, 147, e295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, M.; Akbar, I.E.; Wadood, M.Z.; Shukla, H.; Jorba, J.; Ehrhardt, D. Progress toward Poliomyelitis Eradication—Afghanistan, January 2019–July 2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 1464–1468. [Google Scholar] [CrossRef]
- Platt, L.R.; Estívariz, C.F.; Sutter, R.W. Vaccine-Associated Paralytic Poliomyelitis: A Review of the Epidemiology and Estimation of the Global Burden. J. Infect. Dis. 2014, 210 (Suppl. 1), S380–S389. [Google Scholar] [CrossRef] [PubMed]
- Mbaeyi, C.; Alleman, M.M.; Ehrhardt, D.; Wiesen, E.; Burns, C.C.; Liu, H.; Ewetola, R.; Seakamela, L.; Mdodo, R.; Ndoutabe, M.; et al. Update on Vaccine-Derived Poliovirus Outbreaks—Democratic Republic of the Congo and Horn of Africa, 2017–2018. MMWR Morb. Mortal. Wkly. Rep. 2019, 68, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Saleem, A.F.; Mach, O.; Yousafzai, M.T.; Khan, A.; Weldon, W.C.; Steven Oberste, M.; Zaidi, S.S.; Alam, M.M.; Quadri, F.; Sutter, R.W.; et al. Immunogenicity of Different Routine Poliovirus Vaccination Schedules: A Randomized, Controlled Trial in Karachi, Pakistan. J. Infect. Dis. 2018, 217, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J. Fighting a Polio Outbreak in Papua New Guinea. Lancet 2018, 392, 2155–2156. [Google Scholar] [CrossRef]
- Roberts, L. Polio Eradication Campaign Loses Ground. Science 2019, 365, 106–107. [Google Scholar] [CrossRef] [PubMed]
- McArthur, D.B. Emerging Infectious Diseases. Nurs. Clin. N. Am. 2019, 54, 297–311. [Google Scholar] [CrossRef]
- Hsiung, G.D. Diagnostic Virology: From Animals to Automation. Yale J. Biol. Med. 1984, 57, 727–733. [Google Scholar] [PubMed]
- Ida-Hosonuma, M.; Sasaki, Y.; Toyoda, H.; Nomoto, A.; Gotoh, O.; Yonekawa, H.; Koike, S. Host Range of Poliovirus Is Restricted to Simians Because of a Rapid Sequence Change of the Poliovirus Receptor Gene during Evolution. Arch. Virol. 2003, 148, 29–44. [Google Scholar] [CrossRef]
- Rakoto-Andrianarivelo, M.; Gumede, N.; Jegouic, S.; Balanant, J.; Andriamamonjy, S.N.; Rabemanantsoa, S.; Birmingham, M.; Randriamanalina, B.; Nkolomoni, L.; Venter, M.; et al. Reemergence of Recombinant Vaccine-Derived Poliovirus Outbreak in Madagascar. J. Infect. Dis. 2008, 197, 1427–1435. [Google Scholar] [CrossRef] [Green Version]
- Harvala, H.; Van Nguyen, D.; McIntyre, C.; Ahuka-Mundeke, S.; Ngole, E.M.; Delaporte, E.; Peeters, M.; Simmonds, P. Co-Circulation of Enteroviruses between Apes and Humans. J. Gen. Virol. 2014, 95, 403–407. [Google Scholar] [CrossRef] [Green Version]
- Mombo, I.M.; Berthet, N.; Lukashev, A.N.; Bleicker, T.; Brünink, S.; Léger, L.; Atencia, R.; Cox, D.; Bouchier, C.; Durand, P.; et al. First Detection of an Enterovirus C99 in a Captive Chimpanzee with Acute Flaccid Paralysis, from the Tchimpounga Chimpanzee Rehabilitation Center, Republic of Congo. PLoS ONE 2015, 10, e0136700. [Google Scholar] [CrossRef]
- Mombo, I.M.; Lukashev, A.N.; Bleicker, T.; Brünink, S.; Berthet, N.; Maganga, G.D.; Durand, P.; Arnathau, C.; Boundenga, L.; Ngoubangoye, B.; et al. African Non-Human Primates Host Diverse Enteroviruses. PLoS ONE 2017, 12, e0169067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, S.W.; Sunwoo, S.Y.; Hyun, B.H.; Lyoo, Y.S. Detection of Antibodies against Classical Swine Fever Virus in Fecal Samples from Wild Boar. Vet. Microbiol. 2012, 161, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.A.; Tai, N.; Ekanayake-Alper, D.K.; Peng, J.; Hu, Y.; Hager, K.; Compton, S.; Wong, F.S.; Smith, P.C.; Wen, L. Norovirus Changes Susceptibility to Type 1 Diabetes by Altering Intestinal Microbiota and Immune Cell Functions. Front. Immunol. 2019, 10, 2654. [Google Scholar] [CrossRef] [PubMed]
- Mombo, I.M.; Fritz, M.; Becquart, P.; Liegeois, F.; Elguero, E.; Boundenga, L.; Mebaley, T.N.; Prugnolle, F.; Maganga, G.D.; Leroy, E.M. Detection of Ebola Virus Antibodies in Fecal Samples of Great Apes in Gabon. Viruses 2020, 12, 1347. [Google Scholar] [CrossRef]
- Nieto-Pelegrín, E.; Rivera-Arroyo, B.; Sánchez-Vizcaíno, J.M. First Detection of Antibodies against African Swine Fever Virus in Faeces Samples. Transbound. Emerg. Dis. 2015, 62, 594–602. [Google Scholar] [CrossRef]
- Amona, I.; Medkour, H.; Akiana, J.; Davoust, B.; Tall, M.L.; Grimaldier, C.; Gazin, C.; Zandotti, C.; Levasseur, A.; Scola, B.L.; et al. Isolation and Molecular Characterization of Enteroviruses from Great Apes and Humans in the Republic of Congo: Recombination within Enterovirus C Serotypes. Microorganisms 2020, 8, 1779. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Haitjema, C.H.; Heger, C.D.; Boge, A. A Proof-of-Concept Analysis of Carbohydrate-Deficient Transferrin by Imaged Capillary Isoelectric Focusing and in-Capillary Immunodetection. BioTechniques 2020, 68, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Ha, S.; Tsuji, M.; Fagarasan, S. Intestinal IgA Synthesis: A Primitive Form of Adaptive Immunity That Regulates Microbial Communities in the Gut. Semin. Immunol. 2007, 19, 127–135. [Google Scholar] [CrossRef]
- Fadlallah, J.; El Kafsi, H.; Sterlin, D.; Juste, C.; Parizot, C.; Dorgham, K.; Autaa, G.; Gouas, D.; Almeida, M.; Lepage, P.; et al. Microbial Ecology Perturbation in Human IgA Deficiency. Sci. Transl. Med. 2018, 10, eaan1217. [Google Scholar] [CrossRef] [Green Version]
- Kummitha, C.M.; Mayle, K.M.; Christman, M.A.; Deosarkar, S.P.; Schwartz, A.L.; McCall, K.D.; Kohn, L.D.; Malgor, R.; Goetz, D.J. A Sandwich ELISA for the Detection of Wnt5a. J. Immunol. Methods 2010, 352, 38–44. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sereme, Y.; Zarza, S.M.; Medkour, H.; Amona, I.; Fenollar, F.; Akiana, J.; Mezouar, S.; Orain, N.; Vitte, J.; Davoust, B.; et al. Stool Serology: Development of a Non-Invasive Immunological Method for the Detection of Enterovirus-Specific Antibodies in Congo Gorilla Faeces. Microorganisms 2021, 9, 810. https://doi.org/10.3390/microorganisms9040810
Sereme Y, Zarza SM, Medkour H, Amona I, Fenollar F, Akiana J, Mezouar S, Orain N, Vitte J, Davoust B, et al. Stool Serology: Development of a Non-Invasive Immunological Method for the Detection of Enterovirus-Specific Antibodies in Congo Gorilla Faeces. Microorganisms. 2021; 9(4):810. https://doi.org/10.3390/microorganisms9040810
Chicago/Turabian StyleSereme, Youssouf, Sandra Madariaga Zarza, Hacène Medkour, Inestin Amona, Florence Fenollar, Jean Akiana, Soraya Mezouar, Nicolas Orain, Joana Vitte, Bernard Davoust, and et al. 2021. "Stool Serology: Development of a Non-Invasive Immunological Method for the Detection of Enterovirus-Specific Antibodies in Congo Gorilla Faeces" Microorganisms 9, no. 4: 810. https://doi.org/10.3390/microorganisms9040810
APA StyleSereme, Y., Zarza, S. M., Medkour, H., Amona, I., Fenollar, F., Akiana, J., Mezouar, S., Orain, N., Vitte, J., Davoust, B., Raoult, D., & Mediannikov, O. (2021). Stool Serology: Development of a Non-Invasive Immunological Method for the Detection of Enterovirus-Specific Antibodies in Congo Gorilla Faeces. Microorganisms, 9(4), 810. https://doi.org/10.3390/microorganisms9040810