
Bacillus velezensis CE 100 Inhibits Root Rot Diseases (Phytophthora spp.) and Promotes Growth of Japanese Cypress (Chamaecyparis obtusa Endlicher) Seedlings

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of B. velezensis CE 100 and Phytophthora spp.
2.2. Quantitative Analysis of Lytic Enzymes
2.3. Anti-Oomycete Activity of B. velezensis CE 100 Against Phytophthora spp.
2.4. Indole-3-Acetic Acid (IAA) Production by B. velezensis CE 100
2.5. Experimental Conditions
2.6. Analysis of Survival Rate of C. obtusa Seedlings
2.7. Determination of Seedling Growth Parameters
2.8. Statistical Analysis
3. Results
3.1. Inhibition of Root Rot Diseases Caused by Phytophthora spp. Using B. velezensis CE 100
3.1.1. Growth Pattern of B. velezensis CE 100
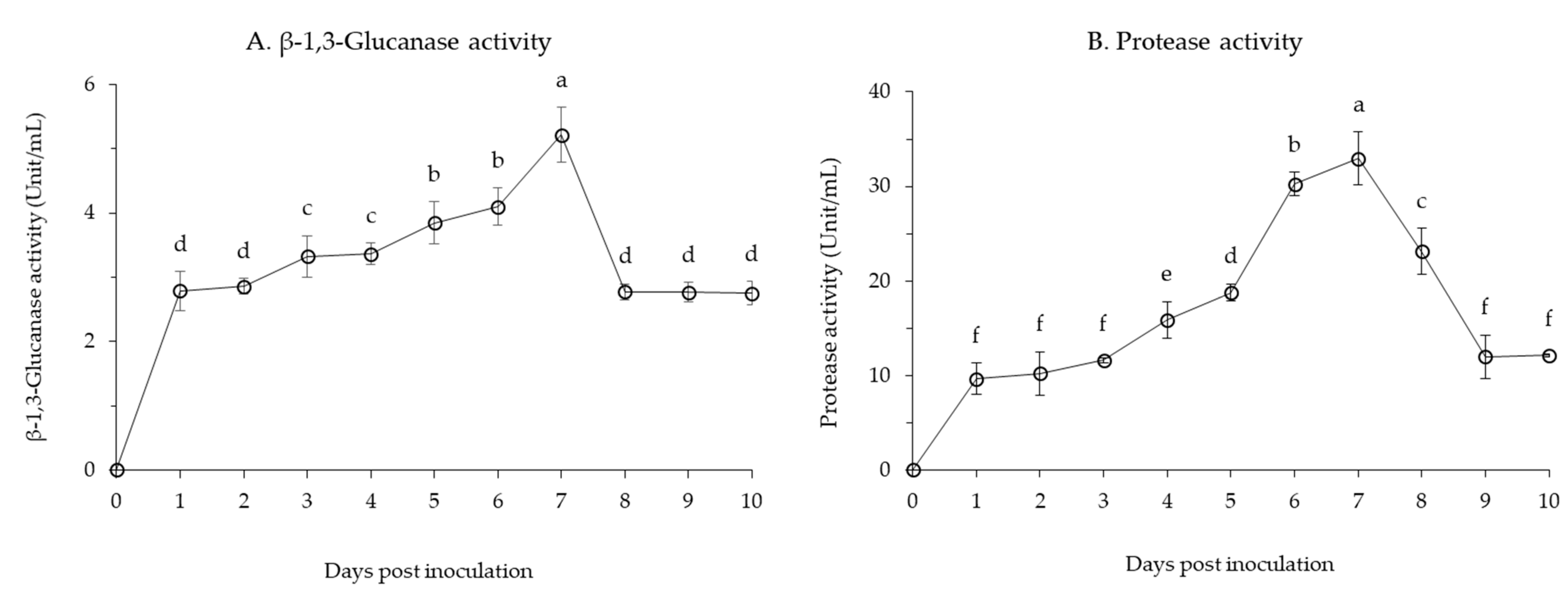
3.1.2. Production of Lytic Enzymes by B. velezensis CE 100
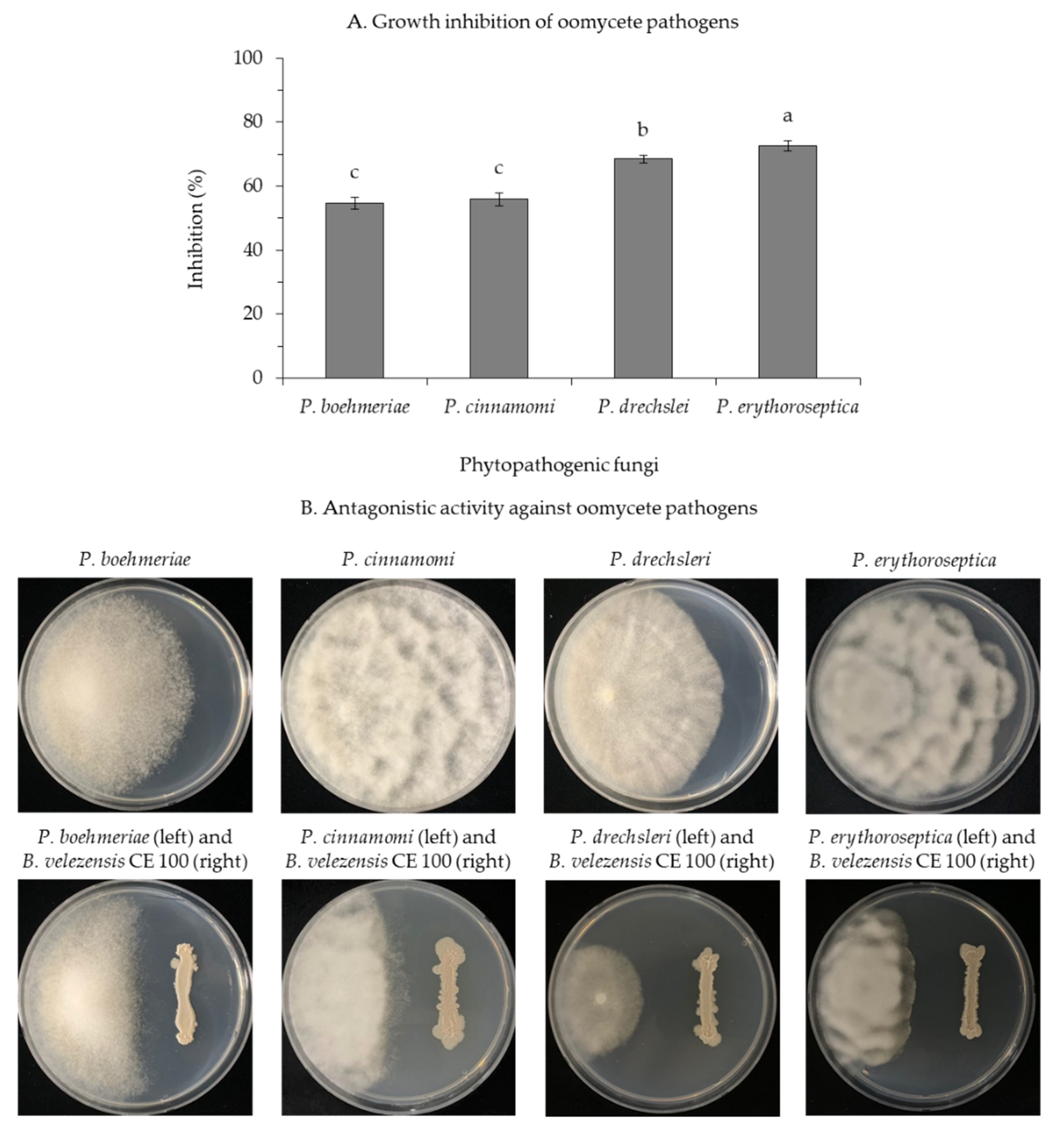
3.1.3. Inhibition of Oomycete Pathogens by B. velezensis CE 100
3.1.4. Survival Rate of C. obtusa Seedlings
3.2. Effect of B. velezensis CE 100 on Growth Promotion of C. obtusa Seedlings
3.2.1. Indole-3-Acetic Acid (IAA) Production of B. velezensis CE 100
3.2.2. Growth and Biomass Yield of C. obtusa Seedlings
4. Discussion
4.1. Anti-Oomycete Activity of B. velezensis CE 100 against Phytophthora spp. and Survival Rate of Seedlings
4.2. Effect of B. velezensis CE 100 on Growth Promotion of C. obtusa Seedlings
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matsumoto, A.; Tani, N.; LI, X.G.; Nakao, Y.; Tomaru, N.; Tsumura, Y. Development and polymorphisms of microsatellite markers for hinoki (Chamaecyparis obtusa). Mol. Ecol. Resour. 2006, 6, 310–312. [Google Scholar] [CrossRef]
- Tian, M.; Han, D.; Row, K.H. Preparation of molecularly imprinted polymer for extracting flavones from Chamaecyparis obtusa. Anal. Lett. 2011, 44, 737–746. [Google Scholar] [CrossRef]
- Tang, B.; Bi, W.; Zhang, H.; Row, K.H. Deep eutectic solvent-based HS-SME coupled with GC for the analysis of bioactive terpenoids in Chamaecyparis obtusa leaves. Chromatographia 2014, 77, 373–377. [Google Scholar] [CrossRef]
- Ikei, H.; Song, C.; Miyazaki, Y. Physiological effect of olfactory stimulation by Hinoki cypress (Chamaecyparis obtusa) leaf oil. J. Physiol. Anthropol. 2015, 34, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shik, M.H.; Woon, K.T.; Solomon, T. Growth Performance Assessment of Chamaecyparis obtusa Stand in Gyeongnam Province, S. Korea. Agric. For. Fish. 2020, 9, 135–141. [Google Scholar] [CrossRef]
- Forestry Statistical Yearbook. Available online: https://www.forest.go.kr/kfsweb/cop/bbs/selectBoardArticle.do;jsessionid=Nz0iyNG4B4kBJtjOhIQQ0yvThcQPtZTF9QxCU93XBxDKK9gYJhZblMP19SFGqLuy.frswas01_servlet_engine5?nttId=3150759&bbsId=BBSMSTR_1064&pageIndex=1&pageUnit=10&searchtitle=title&searchcont=&searchkey=&searchwriter=&searchdept=&searchWrd=&ctgryLrcls=&ctgryMdcls=&ctgrySmcls=&ntcStartDt=&ntcEndDt=&orgId=&mn=NKFS_04_05_09&component= (accessed on 2 November 2020).
- Heiskanen, J. Effects of compost additive in sphagnum peat growing medium on Norway spruce container seedlings. New For. 2013, 44, 101–118. [Google Scholar] [CrossRef]
- Ghorbani, R.; Wilcockson, S.; Koocheki, A.; Leifert, C. Soil management for sustainable crop disease control: A review. Environ. Chem. Lett. 2008, 6, 177–201. [Google Scholar] [CrossRef]
- Paungfoo-Lonhienne, C.; Yeoh, Y.K.; Kasinadhuni, N.R.P.; Lonhienne, T.G.; Robinson, N.; Hugenholtz, P.; Ragan, M.A.; Schmidt, S. Nitrogen fertilizer dose alters fungal communities in sugarcane soil and rhizosphere. Sci. Rep. 2015, 5, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.Y.; Chen, H.Y. Negative effects of fertilization on plant nutrient resorption. Ecology 2015, 96, 373–380. [Google Scholar] [CrossRef]
- Robin, C.; Piou, D.; Feau, N.; Douzon, G.; Schenck, N.; Hansen, E.M. Root and aerial infections of Chamaecyparis lawsoniana by Phytophthora lateralis: A new threat for European countries. For. Pathol. 2011, 41, 417–424. [Google Scholar] [CrossRef]
- Jung, T.; Chang, T.T.; Bakonyi, J.; Seress, D.; Pérez-Sierra, A.; Yang, X.; Hong, C.; Scanu, B.; Fu, C.H.; Hsueh, K.L.; et al. Diversity of Phytophthora species in natural ecosystems of Taiwan and association with disease symptoms. Plant Pathol. 2017, 66, 194–211. [Google Scholar] [CrossRef]
- Thines, M. Taxonomy and Phylogeny of Phytophthora and Related Oomycetes. In Phytophthora: A Global Perspective; Lamour, K., Ed.; CAB International: Oxfordshire, UK, 2013; Volume 2, pp. 11–18. [Google Scholar]
- Mora-Sala, B.; Gramaje, D.; Abad-Campos, P.; Berbegal, M. Diversity of Phytophthora species associated with Quercus ilex L. in three Spanish regions evaluated by NGS. Forests 2019, 10, 979. [Google Scholar] [CrossRef] [Green Version]
- Orlikowski, L.B. Occurrence and harmfulness of Phytophthora spp. in Polish hardy ornamental nursery stock. Acta Hortic. 2010, 885, 243–248. [Google Scholar] [CrossRef]
- Chastagner, G.A.; Benson, D.M. The Christmas tree: Traditions, production and diseases. Plant Health Prog. 2000, 1, 1–15. [Google Scholar] [CrossRef]
- Widmer, T.L. Infective potential of sporangia and zoospores of Phytophthora ramorum. Plant Dis. 2009, 93, 30–35. [Google Scholar] [CrossRef]
- Hansen, E.M. Phytophthora species emerging as pathogens of forest trees. Curr. For. Rep. 2015, 1, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Acero, F.J.; Carbú, M.; Garrido, C.; Collado, I.G.; Cantoral, J.; Vallejo, I. Screening Study of Potential Lead Compounds for Natural Product-based Fungicides Against Phytophthora Species. J. Phytopathol. 2006, 154, 616–621. [Google Scholar] [CrossRef]
- Grünwald, N.J.; Sturbaum, A.K.; Montes, G.R.; Serrano, E.G.; Lozoya-Saldaña, H.; Fry, W.E. Selection for fungicide resistance within a growing season in field populations of Phytophthora infestans at the center of origin. Phytopathology 2006, 96, 1397–1403. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, X.H.; Naing, K.W.; Lee, Y.S.; Kim, Y.H.; Moon, J.H.; Kim, K.Y. Antagonism of antifungal metabolites from Streptomyces griseus H7602 against Phytophthora capsici. J. Basic Microbiol. 2015, 55, 45–53. [Google Scholar] [CrossRef]
- Wightwick, A.; Walters, R.; Allinson, G.; Reichman, S.; Menzies, N. Environmental Risks of Fungicides Used in Horticultural Production Systems. In Fungicides; Carisse, O., Ed.; IntechOpen: Rijeka, Croatia, 2010; pp. 273–304. [Google Scholar] [CrossRef] [Green Version]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 2012, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2016, 2, 1–19. [Google Scholar] [CrossRef]
- El-Sayed, A.S.; Akbar, A.; Iqrar, I.; Ali, R.; Norman, D.; Brennan, M.; Ali, G.S. A glucanolytic Pseudomonas sp. associated with Smilax bona-nox L. displays strong activity against Phytophthora parasitica. Microbiol. Res. 2018, 207, 140–152. [Google Scholar] [CrossRef] [PubMed]
- Sowanpreecha, R.; Rerngsamran, P. Biocontrol of orchid-pathogenic mold, Phytophthora palmivora, by antifungal proteins from Pseudomonas aeruginosa RS1. Mycobiology 2018, 46, 129–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, D.; Yuan, C.; Luo, Y.; Chen, Y.; Lu, M.; Chen, G.; Ren, G.; Cui, C.; Zhang, J.; An, D. Biocontrol of tobacco black shank disease (Phytophthora nicotianae) by Bacillus velezensis Ba168. Pestic. Biochem. Physiol. 2020, 165, 104523. [Google Scholar] [CrossRef]
- Singh, D.; Ghosh, P.; Kumar, J.; Kumar, A. Rhizosphere, Microbiome and Agro-ecology. In Microbial Interventions in Agriculture and Environment, 1st ed.; Singh, D.P., Gupta, V.K., Prabha, R., Eds.; Springer Nature: Singapore, 2019; pp. 205–227. [Google Scholar] [CrossRef]
- Safitri, D.; Wiyono, S.; Soekarno, B.P.W.; Achmad, A. Epiphytic yeasts from Piperaceae as biocontrol agents for foot rot of black pepper caused by Phytophthora capsici. Bio diversitas 2021, 22, 1895–1901. [Google Scholar] [CrossRef]
- Kwon, J.H.; Won, S.J.; Moon, J.H.; Kim, C.W.; Ahn, Y.S. Control of Fungal Diseases and Increase in Yields of a Cultivated Jujube Fruit (Zizyphus jujuba Miller var. inermis Rehder) Orchard by Employing Lysobacter antibioticus HS124. Forests 2019, 10, 1146. [Google Scholar] [CrossRef] [Green Version]
- Won, S.-J.; Choub, V.; Kwon, J.-H.; Kim, D.-H.; Ahn, Y.S. The Control of Fusarium Root Rot and Development of Coastal Pine (Pinus thunbergii Parl.) Seedlings in a Container Nursery by Use of Bacillus licheniformis MH48. Forests 2018, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Won, S.-J.; Kwon, J.-H.; Kim, D.H.; Ahn, Y.S. The effect of Bacillus licheniformis MH48 on control of foliar fungal diseases and growth promotion of Camellia oleifera seedlings in the coastal reclaimed land of Korea. Pathogens 2019, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Egorshina, A.A.; Khairullin, R.M.; Sakhabutdinova, A.R.; Luk’Yantsev, M.A. Involvement of phytohormones in the development of interaction between wheat seedlings and endophytic Bacillus subtilis strain 11BM. Russ. J. Plant Physiol. 2012, 59, 134–140. [Google Scholar] [CrossRef]
- Ahmed, A.; Hasnain, S. Auxins as one of the factors of plant growth improvement by plant growth promoting rhizobacteria. Pol. J. Microbiol. 2014, 63, 261–266. [Google Scholar] [CrossRef]
- Park, H.-G.; Jeong, M.-H.; Ahn, Y.S. Inoculation with Bacillus licheniformis MH48 to improve Camellia japonica seedling development in coastal lands. Turk. J. Agric. For. 2017, 41, 381–388. [Google Scholar] [CrossRef]
- Kang, X.; Zhang, W.; Cai, X.; Zhu, T.; Xue, Y.; Liu, C. Bacillus velezensis CC09: A potential ‘vaccine’ for controlling wheat diseases. Mol. Plant Microbe Interact. 2018, 31, 623–632. [Google Scholar] [CrossRef]
- Tanimoto, E. Regulation of root growth by plant hormones—roles for auxin and gibberellin. Crit. Rev. Plant Sci. 2005, 24, 249–265. [Google Scholar] [CrossRef]
- Perrot-Rechenmann, C. Cellular responses to auxin: Division versus expansion. Cold Spring Harb. Perspect. Biol. 2010, 2, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Majda, M.; Robert, S. The role of auxin in cell wall expansion. Int. J. Mol. Sci. 2018, 19, 951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akgül, D.S.; Mirik, M. Biocontrol of Phytophthora capsici on pepper plants by Bacillus megaterium strains. J. Plant Pathol. 2008, 90, 29–34. [Google Scholar] [CrossRef]
- Lim, J.H.; Kim, S.D. Biocontrol of Phytophthora blight of red pepper caused by Phytophthora capsici using Bacillus subtilis AH18 and B. licheniformis K11 formulations. J. Korean Soc. Appl. Biol. Chem. 2010, 53, 766–773. [Google Scholar] [CrossRef]
- Anjum, M.Z.; Ghazanfar, M.U.; Hussain, I. Bio-efficacy of Trichoderma isolates and Bacillus subtilis against root rot of muskmelon Cucumis Melo L. caused by Phytophthora drechsleri under controlled and field conditions. Pak. J. Bot. 2019, 51, 1877–1882. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; MacDonald, J.E. Why seedlings grow: Influence of plant attributes. New For. 2018, 49, 1–34. [Google Scholar] [CrossRef]
- Sukhbaatar, G.; Ganbaatar, B.; Jamsran, T.; Purevragchaa, B.; Nachin, B.; Gradel, A. Assessment of early survival and growth of planted Scots pine (Pinus sylvestris) seedlings under extreme continental climate conditions of northern Mongolia. J. For. Res. 2020, 31, 13–26. [Google Scholar] [CrossRef] [Green Version]
- Close, D.C.; Paterson, S.; Corkrey, R.; McArthur, C. Influences of seedling size, container type and mammal browsing on the establishment of Eucalyptus globulus in plantation forestry. New For. 2010, 39, 105–115. [Google Scholar] [CrossRef]
- Choi, T.G.; Maung, C.E.H.; Lee, D.R.; Henry, A.B.; Lee, Y.S.; Kim, K.Y. Role of bacterial antagonists of fungal pathogens, Bacillus thuringiensis KYC and Bacillus velezensis CE 100 in control of root-knot nematode, Meloidogyne incognita and subsequent growth promotion of tomato. Biocontrol. Sci. Technol. 2020, 30, 685–700. [Google Scholar] [CrossRef]
- Lee, D.-R.; Maung, C.E.H.; Henry, A.; Kim, K.-Y. Effect of large-scale cultivation of Bacillus amlyoliquefaciens Y1 using fertilizer based medium for control of Citrus Melanose causing Diaporthe citri. Korean J. Soil. Sci. Fert. 2019, 52, 84–92. [Google Scholar] [CrossRef]
- Ghorbel-Frikha, B.; Sellami-Kamoun, A.; Fakhfakh, N.; Haddar, A.; Manni, L.; Nasri, M. Production and purification of a calcium-dependent protease from Bacillus cereus BG1. J. Ind. Microbiol. Biotechnol. 2005, 32, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.C.; Hseu, R.S.; Wang, H.H. Partial purification and characterization of a 1,3-β-d-glucanase from Ganoderma tsugae. J. Ind. Microbiol. 1995, 14, 5–9. [Google Scholar] [CrossRef]
- Rahman, A.; Sitepu, I.R.; Tang, S.Y.; Hashidoko, Y. Salkowski’s reagent test as a primary screening index for functionalities of rhizobacteria isolated from wild dipterocarp saplings growing naturally on medium-strongly acidic tropical peat soil. Biosci. Biotechnol. Biochem. 2010, 74, 2202–2208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina-cÓrdoba, L.K.; Valencia-Mosquera, L.L.; Tarazona-diaz, G.P.; Arias-palacios, J.D.C. Evaluation of the efficacy of a hydrogen peroxide disinfectant. Int. J. Pharm. Pharm. Sci. 2018, 10, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Aira, M.; Gómez-Brandón, M.; Lazcano, C.; Bååth, E.; Domínguez, J. Plant genotype strongly modifies the structure and growth of maize rhizosphere microbial communities. Soil Biol. Biochem. 2010, 42, 2276–2281. [Google Scholar] [CrossRef]
- Hardham, A.R. Cell biology of plant-oomycete interactions. Cell. Microbiol. 2007, 9, 31–39. [Google Scholar] [CrossRef]
- Mélida, H.; Sandoval-Sierra, J.V.; Diéguez-Uribeondo, J.; Bulone, V. Analyses of extracellular carbohydrates in oomycetes unveil the existence of three different cell wall types. Eukaryot. Cell 2013, 12, 194–203. [Google Scholar] [CrossRef] [Green Version]
- Noronha, E.F.; Ulhoa, C.J. Purification and characterization of an endo-β-1,3-glucanase from Trichoderma harzianum. Can. J. Microbiol. 1996, 42, 1039–1044. [Google Scholar] [CrossRef]
- Kaur, R.; Kalia, A.; Lore, J.S.; Sandhu, J.S. Antifungal effect of Trichoderma spp. β-1,3-glucanase on Phytophthora parasitica: Hyphal morphological distortions. J. Phytopathol. 2020, 168, 700–706. [Google Scholar] [CrossRef]
- Thomas, E.L.; Van der Hoorn, R.A. Ten prominent host proteases in plant-pathogen interactions. Int. J. Mol. Sci. 2018, 19, 639. [Google Scholar] [CrossRef] [Green Version]
- Mašková, T.; Herben, T. Root: Shoot ratio in developing seedlings: How seedlings change their allocation in response to seed mass and ambient nutrient supply. Ecol. Evol. 2018, 8, 7143–7150. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Pérez-Sierra, A.; Durán, A.; Jung, M.H.; Balci, Y.; Scanu, B. Canker and decline diseases caused by soil-and airborne Phytophthora species in forests and woodlands. Persoonia 2018, 40, 182–220. [Google Scholar] [CrossRef] [Green Version]
- Overvoorde, P.; Fukaki, H.; Beeckman, T. Auxin control of root development. Cold Spring Harb. Perspect. Biol. 2010, 2, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Jing, H.; Strader, L.C. Interplay of auxin and cytokinin in lateral root development. Int. J. Mol. Sci. 2019, 20, 486. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Survival Rate (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| 2019 | 2020 | |||||||
| November | December | January | February | March | April | May | June | |
| Control | 100 ± 0.0 a | 85.0 ± 5.0 b | 78.3 ± 5.8 b | 66.7 ± 7.6 b | 51.7 ± 7.6 b | 46.7 ± 3.5 b | 45.0 ± 8.7 b | 41.7 ± 5.8 b |
| Chemical fertilizer | 100 ± 0.0 a | 86.7 ± 2.9 b | 81.7 ± 5.8 b | 73.3 ± 2.9 b | 60.0 ± 5.0 b | 58.3 ± 7.6 b | 53.3 ± 5.8 b | 48.3 ± 5.8 b |
| Bacterial inoculation | 100 ± 0.0 a | 100 ± 0.0 a | 100 ± 0.0 a | 91.7 ± 2.9 a | 90.0 ± 5.0 a | 85.0 ± 5.0 a | 81.7 ± 3.5 a | 81.7 ± 3.5 a |
| Treatment | Seedling Growth | Seedling Biomass | |||
|---|---|---|---|---|---|
| Root Collar Diameter (mm) | Shoot Length (cm) | Root Length (cm) | Shoot Dry Weight (kg) | Root Dry Weight (kg) | |
| Control | 2.4 ± 0.3 b | 23.8 ± 2.3 b | 10.3 ± 0.9 c | 13.8 ± 1.7 b | 3.1 ± 0.2 c |
| Chemical fertilizer | 2.6 ± 0.4 b | 25.0 ± 1.7 b | 12.4 ± 1.4 b | 14.3 ± 1.5 b | 3.7 ± 0.4 b |
| Bacterial inoculation | 3.2 ± 0.2 a | 32.6 ± 1.5 a | 15.5 ± 0.9 a | 19.6 ± 1.2 a | 5.8 ± 0.3 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moon, J.-H.; Won, S.-J.; Maung, C.E.H.; Choi, J.-H.; Choi, S.-I.; Ajuna, H.B.; Ahn, Y.S. Bacillus velezensis CE 100 Inhibits Root Rot Diseases (Phytophthora spp.) and Promotes Growth of Japanese Cypress (Chamaecyparis obtusa Endlicher) Seedlings. Microorganisms 2021, 9, 821. https://doi.org/10.3390/microorganisms9040821
Moon J-H, Won S-J, Maung CEH, Choi J-H, Choi S-I, Ajuna HB, Ahn YS. Bacillus velezensis CE 100 Inhibits Root Rot Diseases (Phytophthora spp.) and Promotes Growth of Japanese Cypress (Chamaecyparis obtusa Endlicher) Seedlings. Microorganisms. 2021; 9(4):821. https://doi.org/10.3390/microorganisms9040821
Chicago/Turabian StyleMoon, Jae-Hyun, Sang-Jae Won, Chaw Ei Htwe Maung, Jae-Hyeok Choi, Su-In Choi, Henry B. Ajuna, and Young Sang Ahn. 2021. "Bacillus velezensis CE 100 Inhibits Root Rot Diseases (Phytophthora spp.) and Promotes Growth of Japanese Cypress (Chamaecyparis obtusa Endlicher) Seedlings" Microorganisms 9, no. 4: 821. https://doi.org/10.3390/microorganisms9040821
APA StyleMoon, J.-H., Won, S.-J., Maung, C. E. H., Choi, J.-H., Choi, S.-I., Ajuna, H. B., & Ahn, Y. S. (2021). Bacillus velezensis CE 100 Inhibits Root Rot Diseases (Phytophthora spp.) and Promotes Growth of Japanese Cypress (Chamaecyparis obtusa Endlicher) Seedlings. Microorganisms, 9(4), 821. https://doi.org/10.3390/microorganisms9040821